











 text new page (beta)
text new page (beta) Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO 
 Permalink
PermalinkIntroducción
Detectar los patrones y entender los procesos que determinan la distribución y abundancia de los seres vivos son los objetivos principales de la ecología (Townsend, Begon y Harper, 2008). Por tales razones, se invierte bastante esfuerzo en el desarrollo de métodos de muestreo para estimar la diversidad y composición de las comunidades animales (Magurran y McGill, 2011). En el caso de los estudios de aves, y específicamente en los trópicos donde la riqueza de especies es muy alta, se aplica el método de puntos de conteo (Lynch, 1995, Rappole et al., 1998). Este método de muestreo consiste en que un observador permanece en un sitio por un tiempo determinado con la finalidad de contar e identificar las aves vistas y escuchadas; regularmente, este procedimiento se repite varias veces en lugares distintos con el objetivo de estimar el número de especies y su abundancia, así como sus asociaciones con el hábitat (Bibby et al., 2000, Gregory et al., 2004). De manera general, los puntos de conteo, debido a su sencillez y conveniencia en zonas con densa vegetación, son apropiados para el conteo y monitoreo de Passeriformes y afines y han contribuido en el avance del conocimiento ecológico de la avifauna en los trópicos (Bibby et al., 2000, Carrara et al., 2015, Cavarzere et al., 2012, Martínez-Morales, 2005). Por otra parte, en la última década, gracias al fácil acceso a nuevas tecnologías como las grabadoras de sonido digitales y sistemas autónomos de grabación, el método bioacústico se ha utilizado como una técnica adicional de muestreo. Este método consiste en realizar grabaciones de paisaje sonoro en los sitios de interés con la finalidad de detectar a las aves a través de sus vocalizaciones u otras señales audibles (Brandes, 2008, Celis-Murillo et al., 2009). El método bioacústico busca resolver algunos de los sesgos de los puntos de conteo, tales como las diferencias en la detección de especies entre observadores y los errores de identificación de especies durante el muestreo (Haselmayer y Quinn, 2000, La y Nudds, 2016). El método bioacústico trata también de ofrecer estimaciones confiables de la riqueza de especies, abundancia de individuos, composición y estructura de la comunidad (Brandes, 2008, Celis-Murillo et al., 2012).
Debido a que el método bioacústico es aún incipiente, recientemente se han hecho algunos trabajos de comparación entre este y los puntos de conteo. Estos trabajos comparativos se han realizado en ambientes boscosos templados y tropicales, y han llegado a resultados contrastantes. Por ejemplo, algunos estudios encontraron que el método bioacústico es superior a los puntos de conteo (Haselmayer y Quinn, 2000, Venier et al., 2012, Zwart et al., 2014); también es evidente que no existe una diferencia en el desempeño de ambos métodos (Acevedo y Villanueva-Rivera, 2006, Alquezar y Machado, 2015, Celis-Murillo et al., 2009, Celis-Murillo et al., 2012) y, además, se ha encontrado que los puntos de conteo son más convenientes para el estudio de las aves en bosques templados (Hutto y Stutzman, 2009). No obstante, hasta donde sabemos, no se ha hecho un estudio comparativo entre estas técnicas en el bosque mesófilo de montaña, en donde el estudio de las comunidades de aves es particularmente un reto debido a las condiciones fisonómicas de la vegetación, a su topografía y a las características biológicas de sus especies de aves, ya que muchas de ellas son huidizas y activas en el dosel.
Es claro que toda técnica de conteo de aves tiene ventajas y desventajas. Las ventajas de los puntos de conteo son su sencillez, su versatilidad (pueden llevarse a cabo en muchos ambientes) y su fácil tratamiento estadístico (Hutto, Pletschet y Hendricks, 1986), mientras que las ventajas del método bioacústico son su capacidad para detectar especies raras o que son más vocales, la posibilidad de que sea realizado por personas con poca experiencia en identificación de especies (Hobson, Rempel, Greenwood, Turnbull y van Wilgenburg, 2002), así como el registro permanente de las grabaciones (Haselmayer y Quinn, 2000). Para describir una comunidad biológica es necesario obtener una buena completitud de muestreo (Gotelli, 2008); para esto, algunos investigadores recomiendan utilizar varios métodos en campo (estos dependen del grupo biológico estudiado), con la finalidad de registrar la mayor cantidad posible de especies en un sitio y de obtener datos confiables de la abundancia de cada una de ellas (Nichols et al., 2008). Asimismo, es importante estimar correctamente la diversidad de la comunidad, ya que muchas veces este atributo es utilizado como un indicativo de la biodiversidad y puede servir para tomar decisiones relacionadas con la protección de ciertas áreas (Iknayan, Tingley, Furnas y Beissinger, 2014).
Por tanto, en este estudio se hizo una comparación entre el método de puntos de conteo y el método bioacústico para describir en temporada reproductiva la comunidad de aves de un bosque de niebla en términos de su riqueza, dominancia, estructura y composición, así como en la estimación de la ocupación de las especies (entendida como la proporción del sitio ocupada por una especie). Asimismo, se buscó determinar si el uso simultáneo de ambos métodos de muestreo mejora la obtención de los atributos ecológicos antes mencionados.
Materiales y métodos
El estudio se realizó en el Santuario del Bosque de Niebla, un área de conservación ecológica de aproximadamente 30 ha, administrado por el Instituto de Ecología, A.C. y ubicado en el km 2.5 de la carretera antigua Xalapa-Coatepec (Veracruz, México). El tipo de vegetación predominante es bosque mesófilo de montaña, con algunos remanentes de vegetación secundaria y cafetales (García-Franco, Castillo-Campos, Mehltreter, Martínez y Vázquez, 2008). Las especies dominantes de árboles son Liquidambar styraciflua, Quercus xalapensis, Q. leiophylla, Q. germana y Clethra mexicana y árboles más pequeños como Turpinia insignis, Cinnamomum effusum, Carpinus caroliniana y Oreopanax xalapensis. Algunos arbustos presentes en varios sitios son Palicourea padifolia, Eugenia xalapensis, Miconia glaberrima y Ocotea psychotrioides (García-Franco et al., 2008).
Se seleccionaron 10 puntos de muestreo a lo largo de un sendero que rodea al Santuario del Bosque de Niebla. Los puntos seleccionados tuvieron una distancia entre sí de aproximadamente 250 m para asegurar la independencia del muestreo (Bibby et al., 2000). Cada punto fue muestreado por OSG durante 5 min, tanto por el método de puntos de conteo de radio fijo como por el método bioacústico a través del sistema SRS (Celis-Murillo et al., 2009), el cual está compuesto por un equipo de grabación de 4 canales discretos, micrófonos omnidireccionales y trípode. Todos los muestreos se llevaron a cabo desde el amanecer y hasta 3 h después. Se eligieron 5 min como duración de cada conteo porque es el tiempo suficiente para registrar la mayor cantidad de especies presentes (Lynch, 1995), porque dicho periodo permitió realizar más puntos de conteo en las 3 primeras horas de la mañana (cuando hay mayor actividad de las aves) y porque al ser temporada de reproducción muchas aves territoriales podrían ser contadas 2 veces si el tiempo de registro fuera mayor (Leu et al., 2017). Al llegar a cada punto se esperó alrededor de 3 min para iniciar el conteo, y cada punto fue muestreado con el uso simultáneo de ambos métodos en 3 ocasiones los días 13 de marzo, 6 y 30 de abril del 2013 (excepto el punto 10, el cual no se muestreó el 6 de abril debido a las malas condiciones meteorológicas); se realizaron 29 muestreos de aves por el método de puntos de conteo y 29 por el método bioacústico, lo que dio un total de 58 muestras. Para cada especie detectada se registró el número de individuos, el modo de detección (visual o auditivo) y si se encontraba dentro del radio de 25 m, que es una distancia estándar utilizada como límite visual en ambientes con vegetación densa (Hutto et al., 1986). Las identificaciones de aves en campo se hicieron con la ayuda de la guía de Howell y Webb (1995). Todas las grabaciones se realizaron a una tasa de muestreo de 44.1 kHz y 16 bits, en formato WAV y fueron depositadas en la Biblioteca de Sonidos de las Aves de México (Instituto de Ecología, A.C.). Posteriormente, los datos obtenidos en los puntos de conteo y las grabaciones fueron interpretados y analizados en el laboratorio por el mismo observador (OSG), usando el programa Audacity 2.02 y la base de datos Xeno canto (http://www.xeno-canto.org) y de la Biblioteca de Sonidos de las Aves de México, con la finalidad de identificar las especies de aves.
Por medio del programa ESTIMATES 7.5 (Colwell, 2009), se generó una curva de acumulación de especies y se calculó el estimador de riqueza no paramétrico Chao 1. Se eligió este estimador de riqueza porque se basa en datos de abundancia (número de especies representadas por un individuo en la muestra o “singletons” y el número de especies representadas por 2 individuos en la muestra o “doubletons”) y porque tiene un buen desempeño cuando las muestras son pequeñas (Hortal, Borges y Gaspar, 2006). De esta manera, mediante la comparación de las especies observadas y estimadas, se calculó la eficiencia de muestreo o completitud del inventario. Para expresar la diversidad (número de especies y distribución de la abundancia de individuos: Morin, 2011) de la comunidad se calculó la probabilidad de encuentro interespecífico (PEI) (Gotelli, 2008), el cual es insensible al número de individuos en la muestra y es de fácil interpretación. Este valor oscila entre 0 (cuando la comunidad está compuesta por uno o varios individuos de una sola especie) y 1 (cuando todas las especies que integran la comunidad están representadas por exactamente el mismo número de individuos). Para describir la estructura y la composición de la comunidad se utilizaron curvas de rango de abundancia (Magurran y McGill, 2011) y se resaltaron las 4 especies más abundantes.
Para probar si hubo diferencia significativa entre el número de especies registradas por cada uno de los métodos en cada punto de muestreo y dado que ambas muestras eran dependientes entre sí, se hizo la prueba de rangos con signo de Wilcoxon mediante el paquete estadístico R (R Core Team, 2014). Para evaluar la similitud entre las especies registradas por cada método de muestreo, se utilizó el índice de Bray-Curtis. Este índice compara el número de especies compartidas, así como sus abundancias absolutas entre muestras del mismo tamaño, de modo que cuando su valor es 0 las muestras son completamente diferentes, mientras que cuando su valor es 1 las muestras son idénticas entre sí (Magurran y McGill, 2011).
Para calcular la ocupación (probabilidad de que una especie esté presente en un lugar determinado) y la probabilidad de detección (probabilidad de que una especie sea detectada por un observador) de todas las especies registradas, se llevaron a cabo los análisis single-season (MacKenzie et al., 2002) y multimétodo (Nichols et al., 2008) del programa PRESENCE (Hines, 2006), el cual es un paquete que permite crear modelos de ocupación basados en datos de presencia y ausencia y, sobre todo, distinguir ausencias reales de ausencias falsas. La probabilidad de detección se estima porque el hecho de que una especie no se registre en un lugar determinado no significa que dicha especie no esté presente en ese lugar (MacKenzie et al., 2002). Los modelos de ocupación, que incluyen la probabilidad de detección, tratan de solucionar este problema. Por ejemplo, cuando un observador acude al campo y utiliza un método de muestreo para estudiar a una especie puede detectarla o no detectarla; esta información se puede ordenar en un historial de visitas, que es un vector formado por valores 1 y 0, los cuales expresan detección y no detección, respectivamente, para cada una de las ocasiones en las que dicho observador acudió al sitio. Generalmente, con estos historiales de visita se puede calcular la ocupación y la probabilidad de detección de cada una de las especies registradas (MacKenzie et al., 2002). A diferencia de los métodos tradicionales de estimación de la ocupación (MacKenzie y Bailey, 2004, MacKenzie et al., 2002), en nuestro estudio se calculó la ocupación y la probabilidad de detección de cada una de las especies detectadas por cada uno de los métodos por separado y de manera complementaria (uso combinado de ambos métodos). De esta manera, y siguiendo la metodología de Nichols et al. (2008), que consiste en utilizar varios métodos simultáneos de detección para generar modelos de ocupación, se estimaron de manera más efectiva los parámetros mencionados de cada una de las especies detectadas. Este tipo de modelaje multimétodo hace un uso eficiente de las detecciones y permite hacer inferencias acerca de las probabilidades de detección específicas de cada método (Celis-Murillo et al., 2012, Nichols et al., 2008).
Resultados
Se registró un total de 38 especies mediante el uso de ambos métodos de muestreo (Tabla 1). De estas 38 especies, 35 son residentes permanentes y 3 son residentes de invierno. Del total de registros, 32 especies fueron registradas en los puntos de conteo por el observador y 37 especies por el sistema de grabación (en un total de 145 min de observación y grabación). Por medio del estimador Chao 1, se calculó una riqueza de 34 especies por el método de puntos de conteo, 44 por el método bioacústico y 40 con el uso de ambos métodos. La eficiencia de muestreo por puntos de conteo fue del 94%, por el método bioacústico fue del 84% y por el muestreo simultáneo fue del 95% (Tabla 2). El número de “singletons” y “doubletons” registrados por cada método aparece en la Tabla 2.
Tabla 1 Listado total de especies de aves registradas por ambos métodos de muestreo.
| Nombre científico | Estatus migratorio | Psi (EE) (multimétodo) |
p (EE) (multimétodo) |
Psi (EE) (puntos de conteo) |
p (EE) (puntos de conteo) |
Psi (EE) (método bioacústico) |
p (EE) (método bioacústico) |
|---|---|---|---|---|---|---|---|
| Leptotila verreauxi | R | 0.943 (0.103) | 0.539 (0.072) | 0.9164 (0.1630) | 0.5594 (0.1208) | 0.9164 (0.1630) | 0.5594 (0.1208) |
| Ortalis vetula | R | 0.9471 (0.282) | 0.201 (0.081) | 1 (0.000) | 0.2414 (0.0795) | 1 (0.000) | 0.1379 (0.0640) |
| Amazona albifrons | R | 0.410 (0.235) | 0.207 (0.119) | 1 (0.000) | 0.0690 (0.0471) | 1 (0.000) | 0.0690 (0.0471) |
| Psittacara holochlorus | R | 1 (0.000) | 0.069 (0.033) | 1 (0.000) | 0.0690 (0.0471) | 1 (0.000) | 0.0690 (0.0471) |
| Trogon caligatus | R | 0.652 (0.254) | 0.216 (0.098) | 0.9885 (0.7035) | 0.2094 (0.1718) | 1 (0.000) | 0.0690 (0.0471) |
| Trogon collaris | R | 1 (0.000) | 0.035 (0.024) | 1 (0.000) | 0.0345 (0.0339) | 1 (0.000) | 0.0345 (0.0339) |
| Piaya cayana | R | 1 (0.000) | 0.086 (0.037) | 1 (0.000) | 0.0345 (0.0339) | 1 (0.000) | 0.0690 (0.0471) |
| Momotus momota | R | 1 (0.000) | 0.155 (0.048) | 1 (0.000) | 0.1724 (0.0701) | 0.4122 (0.2645) | 0.3119 (0.2440) |
| Campylopterus curvipennisa | R | 1 (0.000) | 0.0172 (0.0171) | - | - | 1 (0.000) | 0.0345 (0.0339) |
| Colaptes rubiginosus | R | 1 (0.000) | 0.017 (0.017) | 1 (0.000) | 0.0345 (0.0339) | - | - |
| Melanerpes formicivorus | R | 0.562 (0.1853) | 0.3302 (0.0975) | 1 (0.000) | 0.2069 (0.0752) | 0.3608 (0.1915) | 0.4679 (0.2099) |
| Melanerpes aurifrons | R | 0.491 (0.358) | 0.1458 (0.1226) | 1 (0.000) | 0.0690 (0.0471) | 1 (0.000) | 0.0690 (0.0471) |
| Picoides scalaris | R | 1 (0.000) | 0.017 (0.017) | - | - | 1 (0.000) | 0.0345 (0.0339) |
| Micrastur semitorquatusa | R | 1 (0.000) | 0.017 (0.017) | - | - | 1 (0.000) | 0.033 (0.0328) |
| Cyanocorax yncasa | R | 1 (0.000) | 0.017 (0.017) | - | - | 1 (0.000) | 0.0345 (0.0339) |
| Psilorhinus morio | R | 1 (0.000) | 0.672 (0.062) | 1 (0.000) | 0.7586 (0.0795) | 0.9544 (0.1122) | 0.6153 (0.1106) |
| Piranga rubra | RI | 1 (0.000) | 0.035 (0.024) | 1 (0.000) | 0.0345 (0.0339) | 1 (0.000) | 0.0345 (0.0339) |
| Empidonax occidentalis | R | 0.522 (0.235) | 0.227 (0.105) | 1 (0.000) | 0.0690 (0.0471) | 0.8244 (0.6280) | 0.2084 (0.1739) |
| Myiarchus tuberculifer | R | 0.954 (0.109) | 0.380 (0.074) | 1 (0.000) | 0.3103 (0.0859) | 0.620 (0.162) | 0.6819 (0.126) |
| Myiozetetes similis | R | 0.943 (0.105) | 0.403 (0.074) | 1 (0.000) | 0.4138 (0.0915) | 0.9083 (0.088) | 0.400 (0.155) |
| Pitangus sulphuratus | R | 1 (0.000) | 0.069 (0.033) | 1 (0.000) | 0.1379 (0.0640) | 1 (0.000) | 0.1379 (0.0640) |
| Basileuterus culicivorus | R | 1 (0.000) | 0.276 (0.0588) | 1 (0.000) | 0.2069 (0.0752) | 1 (0.000) | 0.3448 (0.0883) |
| Cardellina pusilla | RI | 0.726 (0.303) | 0.188 (0.089) | 0.5092 (0.3794) | 0.2681 (0.2111) | 0.5092 (0.3794) | 0.2681 (0.2111) |
| Sittasomus griseicapillus | R | 1 (0.000) | 0.276 (0.059) | 1 (0.000) | 0.3793 (0.0901) | 1 (0.000) | 0.1724 (0.070) |
| Catharus mexicanus | R | 1 (0.000) | 0.2241 (0.055) | 1 (0.000) | 0.2759 (0.0830) | 1 (0.000) | 0.1724 (0.070) |
| Turdus grayi | R | 0.5918 (0.2024) | 0.2860 (0.0964) | 1 (0.000) | 0.1034 (0.0566) | 0.4633 (0.1931) | 0.51 (0.1775) |
| Polioptila caerulea | R | 0.787 (0.386) | 0.155 (0.092) | 1 (0.000) | 0.1379 (0.0640) | 1 (0.000) | 0.1034 (0.0566) |
| Dumetella carolinensisa | RI | 1 (0.000) | 0.035 (0.024) | - | - | 1 (0.000) | 0.0345 (0.0339) |
| Campylorhynchus zonatus | R | 1 (0.000) | 0.052 (0.029) | 1 (0.000) | 0.0345 (0.0339) | 1 (0.000) | 0.0690 (0.0471) |
| Henicorhina leucophrys | R | 1 (0.000) | 0.224 (0.055) | 1 (0.000) | 0.1724 (0.0701) | 0.6331 (0.239) | 0.4293 (0.1648) |
| Pheugopedius maculipectus | R | 0.855 (0.319) | 0.183 (0.086) | 0.8244 (0.6280) | 0.2084 (0.1739) | 1 (0.000) | 0.1379 (0.064) |
| Dives dives | R | 1 (0.000) | 0.172 (0.05) | 1 (0.000) | 0.2414 (0.0795) | 1 (0.000) | 0.103 (0.056) |
| Quiscalus mexicanus | R | 1 (0.000) | 0.103 (0.04) | 1 (0.000) | 0.1034 (0.0566) | 1 (0.000) | 0.103 (0.056) |
| Arremon brunneinucha | R | 1 (0.000) | 0.035 (0.024) | 1 (0.000) | 0.0345 (0.0339) | 1 (0.000) | 0.0345 (0.0339) |
| Chlorospingus flavopectus | R | 1 (0.000) | 0.552 (0.065) | 1 (0.000) | 0.5862 (0.0915) | 1 (0.000) | 0.5172 (0.0928) |
| Euphonia elegantissimaa | R | 1 (0.000) | 0.172 (0.05) | - | - | 0.8244 (0.6280) | 0.2084 (0.1739) |
| Euphonia hirundinacea | R | 1 (0.000) | 0.121 (0.043) | 1 (0.000) | 0.1379 (0.0640) | 1 (0.000) | 0.1034 (0.0566) |
| Saltator atriceps | R | 0.633 (0.341) | 0.162 (0.097) | 1 (0.000) | 0.1034 (0.0566) | 0.2709 (0.2059) | 0.3750 (0.264) |
Psi es la proporción de área ocupada por la especie y p es su probabilidad de detección. Entre paréntesis se muestra el error estándar. Estos 2 parámetros fueron calculados con los historiales de detección de cada método por separado y utilizados simultáneamente en 10 puntos de muestreo, que fueron visitados 3 veces.
EE: error estándar; R: residente todo el año; RI: residente de invierno.
a Especies detectadas solamente por el método bioacústico.
Tabla 2 Comparación de 3 métodos para el estudio de las aves en el Santuario del Bosque de Niebla (Xalapa, Veracruz, México).
| Método | Especies observadas | Especies estimadas | “Singletons” | “Doubletons” | Eficiencia (%)a | PEI |
|---|---|---|---|---|---|---|
| Puntos de conteo | 32 | 34 | 6 | 3 | 94 | 0.94 |
| Método bioacústico | 37 | 44 | 9 | 4 | 84 | 0.96 |
| Multimétodo | 38 | 40 | 15 | 7 | 95 | 0.95 |
Los datos fueron obtenidos en 10 puntos de muestreo visitados 3 veces.
PEI: probabilidad de encuentro interespecífico.
a Porcentaje de especies observadas en relación con las estimadas.
Atlapetes brunneinucha, Piranga rubra y Trogon collaris fueron singletones en ambos métodos, por lo que pueden considerarse especies raras en el sitio y momento de muestreo. Pitangus sulphuratus, Euphonia hirundinacea y Empidonax occidentalis fueron registrados como “singletons” por el método de puntos de conteo, pero fueron registradas más de 2 veces por el método bioacústico, lo que indica que a menudo no fueron detectadas por el observador. Por el contrario, Melanerpes aurifrons, Trogon caligatus y Piaya cayana fueron “singletons” en el método bioacústico, pero fueron registrados más de 2 veces por el método de puntos de conteo, debido a que sus avistamientos fueron relativamente frecuentes. Todas las especies exclusivas de cada método fueron “singletons”, es decir, solo se detectó un individuo de dichas especies en el muestreo.
Los valores de diversidad (expresada por la PEI) obtenidos con el uso de ambos métodos de manera separada y en conjunto se acercaron mucho a 1 (el valor máximo que puede tomar este valor) y fueron muy similares entre sí (Tabla 2), lo que indica una alta equidad en la comunidad y una alta coincidencia entre métodos.
Se encontró una alta concordancia entre las curvas de rango abundancia obtenidas por cada método usado de manera independiente y de forma conjunta, con las mismas especies más abundantes (Fig. 1). Esto indica que, en general, hay consistencia en los datos de abundancias relativas obtenidas con ambos métodos.
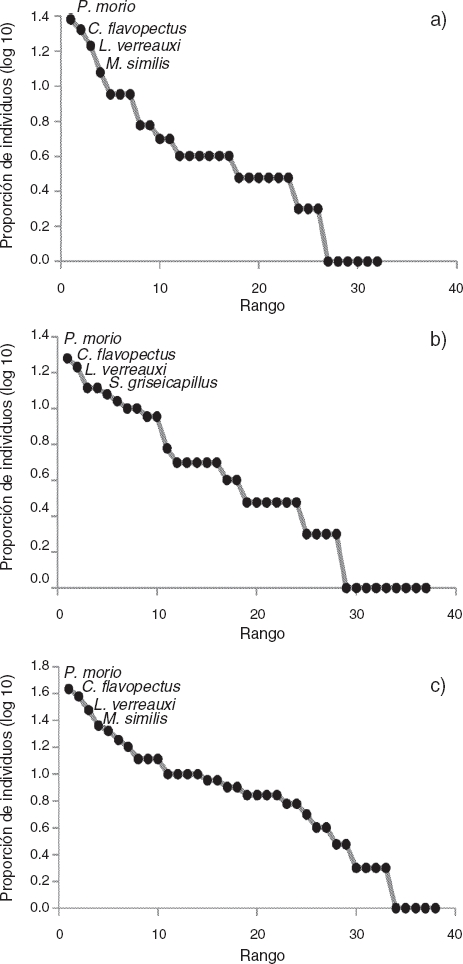
Figura 1 Curvas de rango abundancia. a) Puntos de conteo, b) grabaciones de paisaje sonoro y c) multimétodo. Se muestran las 4 especies más abundantes.
No encontramos diferencia significativa entre el número de especies registrado por cada uno de los métodos en cada punto (W = 322, p > 0.05). El valor del índice de similitud Bray-Curtis obtenido fue de 0.77. Todas las especies registradas por el método de puntos de conteo excepto Colaptes rubiginosus fueron registradas también por el método bioacústico. Las especies exclusivas del método bioacústico fueron Micrastur semitorquatus, Dumetella carolinensis, Picoides scalaris, Campylopterus curvipennis, Cyanocorax yncas y Euphonia elegantissima. Estas 2 últimas especies son gregarias, mientras que las demás son conocidas por sus hábitos solitarios (Howell y Webb, 1995). M. semitorquatus es una especie rara de observar, por lo que es más común que se la detecte por la vocalización.
Las probabilidades de detección (p), así como la proporción de área total ocupada por cada especie (psi) calculadas con ambos métodos por separado y por el muestreo combinado se pueden apreciar en la Tabla 1. Por ejemplo, las especies Leptotila verreauxi, Psilorhinus morio y Chlorospingus flavopectus tuvieron una p relativamente alta, mientras que T. collaris, D. carolinensis y P. rubra tuvieron una p muy cercana a cero. En general, se obtuvieron resultados muy similares tanto de probabilidades de detección como de ocupación mediante el uso de ambas técnicas por separado y con la combinación de métodos. Se registraron 2 especies con categoría “sujeta a protección especial” según la NOM-059-SEMARNAT-2010 (Semarnat, 2010) cuyas probabilidades de detección fueron muy bajas: M. semitorquatus (p = 0.017) y T. collaris (p = 0.035) (Tabla 2).
Discusión
Los resultados demuestran que los métodos de muestreo que examinamos en este trabajo (puntos de conteo y método bioacústico) pueden representar a la comunidad de aves en nuestro sitio de estudio y en temporada reproductiva de manera muy similar, aunque el tipo de información obtenida por cada método varió ligeramente. Por ejemplo, en relación con la completitud del inventario, los puntos de conteo tuvieron un mejor desempeño que el método bioacústico. Esto es similar a lo encontrado por Hutto y Stutzman (2009), en cuyo trabajo los puntos de conteo estimaron una riqueza de especies superior al método bioacústico en bosques templados de Canadá. Sin embargo, el método bioacústico resultó más efectivo para realizar registros adicionales, ya que permitió la detección de 6 especies exclusivas, lo que coincide con lo reportado por Haselmayer y Quinn (2000), quienes pudieron registrar de mejor forma especies muy vocales pero poco observables en los bosques tropicales del Perú. Además, los resultados de este trabajo, al igual que los de Venier et al. (2012) y Zwart et al. (2014), confirmaron la efectividad del método bioacústico en la detección de especies raras. Asimismo, acorde con lo reportado en trabajos anteriores (Acevedo y Villanueva-Rivera, 2006, Alquezar y Machado, 2015, Celis-Murillo et al., 2009, Celis-Murillo et al., 2012), la diversidad, estructura y composición de la comunidad, la probabilidad de detección y ocupación fueron similares entre ambos métodos de muestreo. Cabe destacar que dichos trabajos fueron desarrollados en ambientes tropicales, lo que indica una pauta de complementariedad de métodos en esta región. Los hallazgos de este trabajo indican que, en el estudio de comunidades de aves en bosque mesófilo de montaña y en temporada reproductiva, el desempeño de los puntos de conteo y el método bioacústico es muy similar. Sin embargo, se debe considerar que la escala espacial del muestreo fue pequeña (en total, se cubrieron unas 30 ha), por lo que no sería prudente extrapolar estos resultados a escalas geográficas mayores. Sin embargo, para realizar la comparación de métodos, que fue uno de los fines de este estudio, las escalas espacial y temporal fueron adecuadas, y es de esperar un resultado similar en áreas grandes y en temporadas más largas si se aumenta tanto el número de puntos como el periodo. No obstante, si los muestreos se realizan en temporada de migración se podría tener un problema con el estimador de riqueza, ya que la continua aparición de especies nuevas (transeúntes en la zona) podría elevar el número de “singletons” (Gotelli, 2008) y con ello la imprecisión de tal estimador.
Uno de los objetivos principales de nuestro estudio fue determinar si el uso simultáneo de ambos métodos arrojaría mejores estimaciones de los atributos de la comunidad. En cuanto a la completitud del muestreo, se encontró que la combinación de métodos permitió registrar 38 especies de aves, una más que el método bioacústico y 6 más que los puntos de conteo. En cuanto a diversidad, estructura y composición de la comunidad, probabilidad de detección y ocupación, el uso simultáneo de ambas técnicas tuvo un desempeño similar al de su empleo por separado.
Específicamente, el estimador de riqueza Chao 1 mostró que la mayor completitud de muestreo se realizó por medio de los puntos de conteo y la combinación de métodos. El método bioacústico no resultó tan eficiente en cuanto al porcentaje de completitud, lo que seguramente se debió a la mayor cantidad de “singletons” (especies representadas por un solo individuo en la muestra), que es un parámetro utilizado por el estimador Chao 1 para calcular el número de especies no detectadas (Gotelli, 2008). Seguramente, si se aumentara el tiempo de muestreo, las especies que se registraron como “singletons” y “doubletons” se volverían más abundantes en las muestras. Sin embargo, hay algunas especies como M. semitorquatus, T. collaris y A. brunneinucha que son raras en el sitio. Otras especies “singletons” como D. carolinensis y P. rubra son migratorias y comunes en la región en temporada de invernada. Sin embargo, al momento de muestreo pudieron haber sido raras debido a que en dicha época la mayoría habrían migrado hacia el norte. Para las especies migratorias, sería necesario muestrear en plena temporada de invernada, aunque otros métodos, como el uso de redes niebla, han sido más utilizados para estudiar a este grupo de aves en los trópicos (véase Karr, 1981). Con base en la proporción de especies registradas y estimadas, es probable que con un muestreo más se hubiese obtenido una eficiencia de muestreo muy cercana al 100%.
Los bosques tropicales son ecosistemas con una alta diversidad de especies de aves (Karr, 1981). La comunidad de aves estudiada presentó una alta PEI, lo que corresponde a un valor alto de diversidad. Aunque se esperaba que tanto la combinación de métodos como el método bioacústico (por su gran capacidad de detectar los individuos en campo), o los puntos de conteo (por la inclusión de detecciones tanto vistas como escuchadas) por separado mostraran mayor o menor número de detecciones, estas fueron consistentes en el valor calculado de la PEI y en las curvas de rango abundancia, con las mismas especies más dominantes (P. morio, C. flavopectus y L. verreauxi). Este hallazgo es consistente con lo descrito por Hobson et al. (2002), en el sentido de una alta concordancia entre los valores de abundancias relativas obtenidos con ambas técnicas (aunque cabe resaltar que ellos trabajaron en bosques boreales). Esto es importante porque demuestra que el método bioacústico puede generar datos de abundancia relativa consistentes para estimar la diversidad de las comunidades de aves.
Estudiar comunidades biológicas conlleva la consideración de que la probabilidad de detección de cada especie es distinta (Iknayan et al., 2014). Esto implica que un método de campo determinado podría ser insuficiente para detectar una especie cuando está presente y dar lugar a falsos negativos (o falsas ausencias). Nichols et al. (2008) recomiendan que cuando se utilicen varios métodos para estudiar fauna silvestre se evalúe su efectividad tanto por separado como en conjunto, para conocer su exactitud en la estimación de probabilidad de detección y estimación de ocupación. En este caso, tanto el uso de los métodos separados como en conjunto arrojaron resultados similares en la probabilidad de detección y el valor de ocupación, lo que nos indica que ambos métodos son igual de efectivos para detectar las especies presentes en la comunidad estudiada, es decir, que las especies pueden ser registradas tanto por avistamiento como por señales acústicas. Los valores de ocupación obtenidos pueden servir como punto de referencia para iniciar un programa de seguimiento de las distintas especies, ya que dicho parámetro representa la proporción de área total ocupada por la especie y es una alternativa a la estimación de las abundancias absolutas (Nichols et al., 2008), cuya obtención generalmente es costosa en términos de tiempo y recursos.
Alternativamente al método tradicional de puntos de conteo, que puede tener sesgos en bosques tropicales, se han desarrollado métodos de evaluación biológica rápida (p. ej., Alonso y Naskrecki, 2011, Herzog et al., 2002), los cuales son inventarios de biodiversidad llevados a cabo en tiempos cortos. Para el caso de las aves, los métodos basados en listas de especies (MacKinnon y Phillips, 1993) se han revelado como una alternativa que ofrece inventarios relativamente completos y que, además, son adecuados para regiones tropicales. En nuestro estudio, los métodos utilizados tanto por separado como simultáneamente tuvieron altos desempeños en completitud de inventario y consistencia en la descripción de la comunidad. Algunos autores han señalado que dichos métodos de evaluación rápida deben ser utilizados con precaución (Cavarzere et al., 2012, O’Dea et al., 2004), pues los datos de frecuencia o abundancia relativa generados a partir de estos pueden ser inexactos. Este trabajo propone que tanto el método bioacústico como el de puntos de conteo son buenas alternativas a las listas de especies y pueden funcionar como herramientas de evaluación rápida.
El método bioacústico puede tener un papel muy relevante si no se cuenta con observadores experimentados: este es un problema recurrente en los trópicos, en donde existe poca gente que pueda identificar certeramente a las especies de aves de su región (Haselmayer y Quinn, 2000). Por medio del método bioacústico, incluso personas con poco entrenamiento en ornitología pueden hacer análisis similares en efectividad a los realizados por personas más experimentadas si cuentan con grabaciones de referencia adecuadas (Goyette, Howe, Wolf y Robinson, 2011). El uso del método bioacústico se debe volver general, ya que ofrece mayores posibilidades para el estudio de las aves. Entre sus ventajas se encuentran el hecho de que no es invasivo, que reduce los sesgos de identificación y que deja un registro grabado que puede ser utilizado posteriormente. Ante la continua degradación de los hábitats en el mundo, es necesario implementar programas de monitoreo acústico para el estudio de la biodiversidad y, específicamente, la grabación de paisajes sonoros (Campos-Cerqueira y Aide, 2016).
Para finalizar, cabe resaltar los beneficios de utilizar el método bioacústico. En primer lugar, las grabaciones adquieren un gran valor, ya que pueden ser almacenadas en colecciones con la finalidad de crear las necesarias bibliotecas de sonidos (Brandes, 2008), además de que pueden ser utilizadas con fines de entrenamiento, cuantificación de la frecuencia de canto y análisis del paisaje sonoro (Celis-Murillo et al., 2009). Si son registradas con la información adecuada como fecha y hora, localización, condiciones meteorológicas, así como notas de la biología de las especies, las grabaciones bioacústicas pueden adquirir un alto valor documental y científico (Acevedo y Villanueva-Rivera, 2006). En todo caso, es necesario incluir en cualquier programa de seguimiento tanto el método bioacústico como el de puntos de conteo (debido a que son complementarios) y evaluar el desempeño de ambas técnicas en áreas grandes, en distintos ambientes tropicales y en diferentes épocas del año.

