











 text new page (beta)
text new page (beta) Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO 
 Permalink
PermalinkIntroducción
Los fondos blandos son considerados el hábitat bentónico más extenso sobre la plataforma continental, pendientes y cuencas marinas. Estos se encuentran constituidos por sedimentos arenosos y fangosos, los cuales difieren en el tamaño de grano, proporcionando así hábitats distintos para diferentes especies de organismos (Schiff, Allen, Zeng y Bay, 2000). El tipo de sedimento, incluyendo el tamaño de grano, porosidad, contenido orgánico y productos orgánicos que estos pudieron haber absorbido, se encuentra estrechamente relacionado con la distribución de los organismos que constituyen las comunidades. Los anélidos poliquetos son considerados un grupo dominante en dichas comunidades debido a su diversidad, abundancia y funcionalidad ecológica, lo que les permite una alta adaptabilidad a los diferentes hábitats (Díaz-Castañeda y Harris, 2004). Asimismo, los poliquetos forman parte de los grupos pioneros en las áreas en donde existe alguna perturbación ambiental de cualquier tipo (Díaz-Castañeda y Almeda-Jauregui, 1999; Heilskov y Holmer, 2001). Estos organismos son una parte esencial en las cadenas alimenticias, sirviendo, además, como importantes indicadores de las condiciones del medio ambiente (Díaz-Castañeda, de León y Solana, 2005) e intervienen directamente en el proceso de sedimentación, proporcionando un enlace entre la meiofauna y la macrofauna, permitiendo así el buen funcionamiento del medio (Cacabelos, Moreira y Troncoso, 2006). Considerando la importancia de los poliquetos en la estructura de los ecosistemas acuáticos, se han realizado diversos estudios en la plataforma continental de la costa occidental de la península de Baja California (De León-González, 1988, 1990, 1992, 1994a,b, 1998; Fauchald, 1968, 1970, 1972, 1982a,b; Hartman, 1939a,b, 1940, 1941, 1944a,b,c, 1950, 1956, 1957 y Hartman, 1961; Salazar-Vallejo, 1987; asimismo de De León-González, Hernández-Guevara y Rodríguez-Valencia, 2006; De León-González y Díaz-Castañeda, 1998; De León-González y Rodríguez-Valencia, 1996; De León-González y Solís-Weiss, 1998, 2000, 2001; Díaz-Castañeda, de León-González y Solana-Arellano, 2014; Díaz-Castañeda y de León-González, 2007; Díaz-Castañeda y Harris, 2004; Díaz-Castañeda y San Martín, 2001; Díaz-Castañeda et al., 2005). En el presente estudio se determina la composición taxonómica de la fauna de poliquetos, los cambios espaciales a lo largo de la plataforma continental de la zona de estudio, así como las variaciones espaciales de la densidad y las afinidades faunísticas entre las localidades de muestreo, evaluando la diversidad alfa y determinando sus variaciones geográficas.
Materiales y métodos
En octubre de 1995, dentro del marco del proyecto SIMSUP VIII «Sistemas Marinos de Surgencias y su relación con Pelágicos menores» realizado en la costa occidental de la península de Baja California a bordo del buque oceanográfico «El Puma», fueron recolectadas muestras de fondos blandos en 60 estaciones por medio de una draga Smith-McIntyre (0.1 m2). Las estaciones se dividieron en zona norte: desde punta Colonet hasta punta San Antonio; zona centro: desde punta Eugenia hasta San Hipólito, y zona sur: en la boca de bahía Magdalena (fig. 1). En la tabla 1 se muestran las coordenadas de las 30 estaciones positivas referentes en este trabajo. Cada transecto se dividió batimétricamente en 4 intervalos de profundidad: de 0-49 m, de 50-99 m, de 100-149 m y de 150-200 m, tal como ha sido sugerido por Hernández-Alcántara (2002); McConnaughey (1978) y Ross (1977). La cuantificación de organismos recolectados se llevó a cabo por especie y su registro en las tablas respectivas se presenta en términos de densidad (orgs/0.1 m2), que es el área de muestreo de la draga Smith-McIntyre utilizada en la toma de muestras. La representación y análisis de la distribución espacial de la densidad y el número de especies se realizó por medio de mapas de distribución. Para esto, se incluyeron diversos índices y tratamientos estadísticos tales como: diversidad alfa, equitatividad de Pielou y dominancia de Simpson. Con la finalidad de analizar las variaciones norte-sur de la fauna de poliquetos y comparar las diferencias en la composición de especies a lo largo del margen peninsular, se evaluó la diversidad Beta (Gray, 2000). La utilización de la diversidad Beta en la costa occidental de la península de Baja California permitió detectar cambios bióticos en sentido norte-sur, por medio del índice propuesto por Wilson y Shmida (1984). Para seleccionar adecuadamente las especies representativas de una comunidad se determinaron las especies dominantes tanto en las localidades de muestreo como en las diferentes regiones geográficas mediante la dominancia específica.
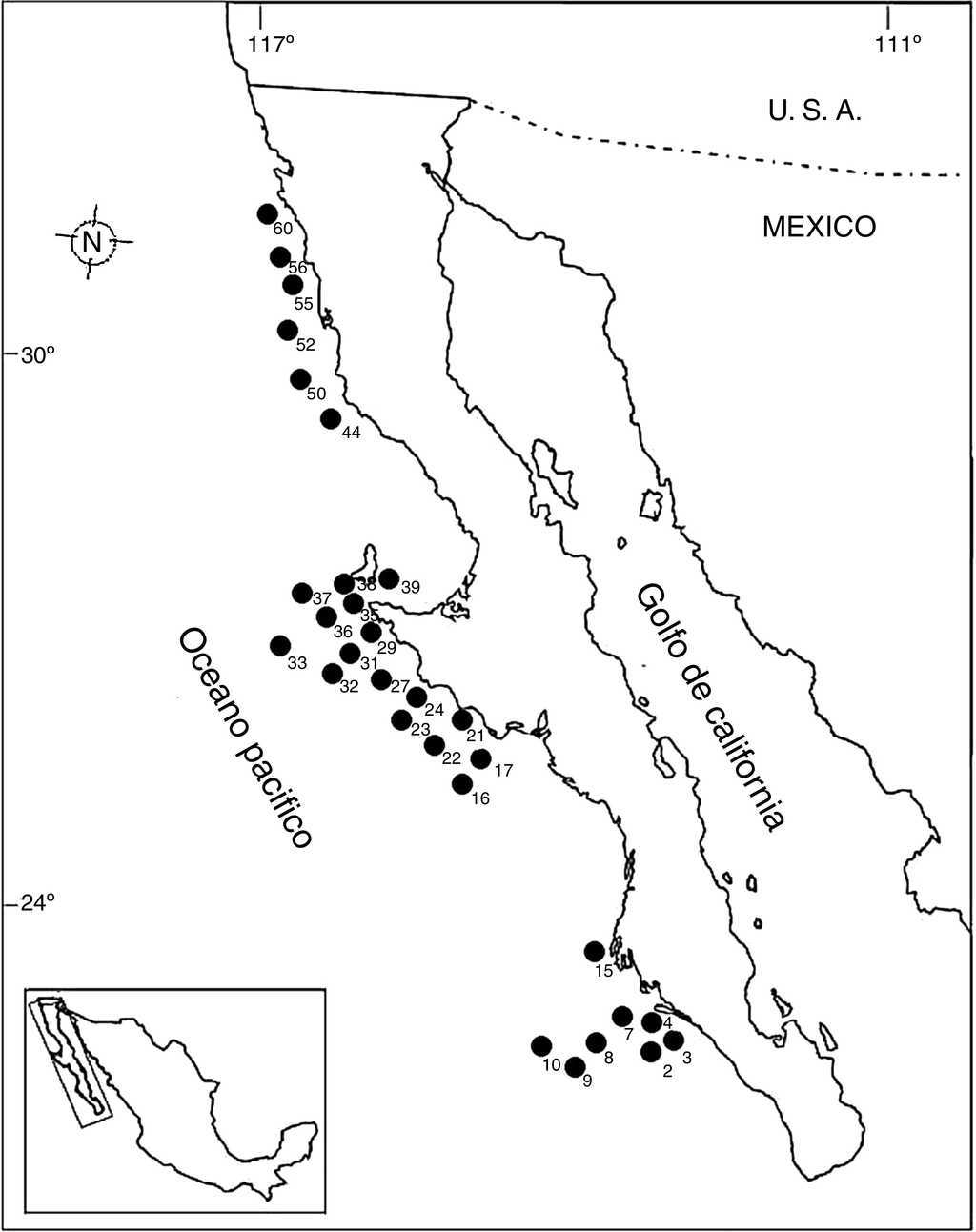
Figura 1 Área de estudio (punta Colonet a boca de bahía Magdalena) en el que se distinguen las 3 zonas de muestreo.
Tabla 1 Estaciones y posición geográfica establecidas dentro del margen del muestreo.
| Zona | Estación | Latitud N | Longitud O |
|---|---|---|---|
| 2 | 23°57′00.00″ | 111°49′00.01″ | |
| 3 | 24°10′00.01″ | 111°40′00.01″ | |
| 4 | 24°16′59.99″ | 111°54′00.00″ | |
| Sur | 7 | 24°25′59.99″ | 112°06′18.00″ |
| 8 | 24°15′00.00″ | 112°13′30.00″ | |
| 9 | 24°04′00.01″ | 112°22′00.01″ | |
| 10 | 24°10′59.99″ | 112°33′18.00″ | |
| 15 | 24°48′00.00″ | 112°25′00.01″ | |
| 16 | 26°16′00.01″ | 114°05′17.99″ | |
| 17 | 26°28′18.01″ | 113°55′00.01″ | |
| 21 | 26°52′00.01″ | 113°55′00.01″ | |
| 22 | 26°38′17.99″ | 114°02′17.99″ | |
| 23 | 26°40′59.99″ | 114°25′00.01″ | |
| 24 | 26°51′00.00″ | 114°28′59.99″ | |
| 27 | 27°01′00.01″ | 114°43′00.01″ | |
| Centro | 29 | 27°27′00.00″ | 114°43′00.01″ |
| 31 | 27°00′00.00″ | 115°07′00.01″ | |
| 32 | 27°12′00.00″ | 115°17′26.99″ | |
| 33 | 27°34′41.28″ | 115°43′33.39″ | |
| 35 | 27°49′18.01″ | 115°09′00.00″ | |
| 36 | 27°37′00.01″ | 115°19′59.99″ | |
| 37 | 27°49′18.01″ | 115°30′00.00″ | |
| 38 | 28°00′00.00″ | 115°19′59.99″ | |
| 39 | 28°00′00.00″ | 115°03′00.00″ | |
| 44 | 29°46′00.01″ | 115°12′01.80″ | |
| 50 | 29°55′00.01″ | 116°10′59.99″ | |
| 52 | 30°19′59.99″ | 116°06′00.00″ | |
| Norte | 55 | 30°32′17.99″ | 116°10′59.99″ |
| 56 | 30°46′59.99″ | 116°16′18.01″ | |
| 60 | 31°16′00.01″ | 116°31′00.01″ |
Resultados
Análisis global de la riqueza y abundancia específica
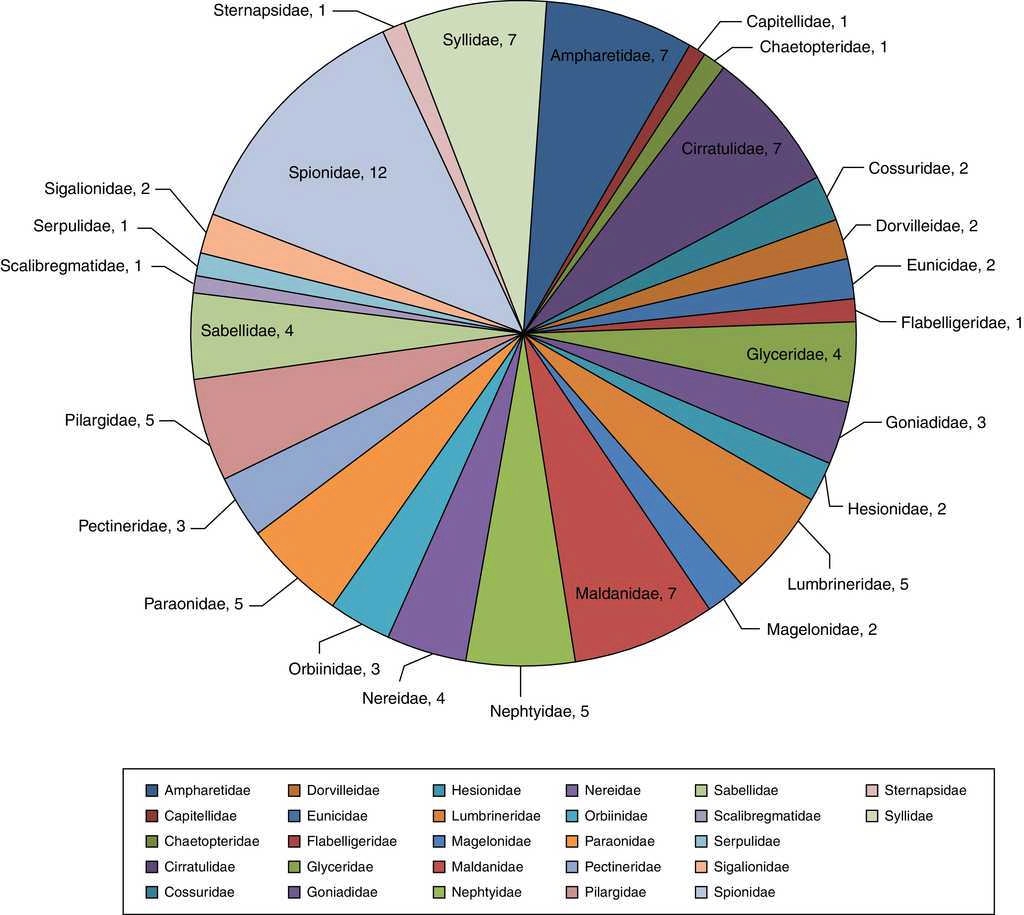
Se examinaron un total de 3,709 organismos incluidos en 62 géneros y 99 especies (Anexo 1). La familia Spionidae presentó la mayor riqueza, constituida por 12 especies y 4 géneros: Minuspio cirrifera, Minuspio perkinsi, Paraprionospio alata, Paraprionospio pinnata, Prionospio cristata, Prionospio dubia, Prionospio jubata, Prionospio malmgreni, Prionospio vermillionensis, Spiophanes bombyx, Spiophanes kroeyeri y Spiophanes wigleyi, las mismas que componen el 12.2% de la diversidad global y el 42.7% de la abundancia relativa (1,583 organismos). Las familias Syllidae (Exogone dispar, Opisthosyllis brunnea, Opisthosyllis japonica, Syllis caeca, Syllis gracilis, Trypanosyllis aeolis y Trypanosyllis zebra), Ampharetidae (Amphicteis scaphobranchiata, Eclysippe trilobata, Lysippe labiata, Melinna oculata, Melinna tentaculata, Melinnampharete gracilis y Paralysippe annectens), Cirratulidae (Aphelochaeta monilaris, Aphelochaeta parvus, Caulleriella gracilis, Chaetozone corona, Chaetozone setosa, Monticellina serratiseta y Monticellina tesselata) y Maldanidae (Clymenella complanata, Clymenella mucosa, Clymenura gracilis, Maldane sarsi, Praxillella gracilis, Praxillella pacifica y Rhodine bitorquata) agrupan el 28.2% del total de especies y 24.0% de abundancia —889 organismos— (fig. 2). Las familias que mostraron la menor diversidad específica fueron Capitellidae (Capitella sp.), Chaetopteridae (Spiochaetopterus costarum), Flabelligeridae (Pherusa neopapillata), Scalibregmatidae (Scalibregma inflatum), Serpulidae (Hydroides gracilis), Sternapsidae (Sternaspis fossor), todas con una sola especie, englobando el 7.7% (287) de abundancia del total de individuos.
Análisis de la riqueza específica en la zona sur
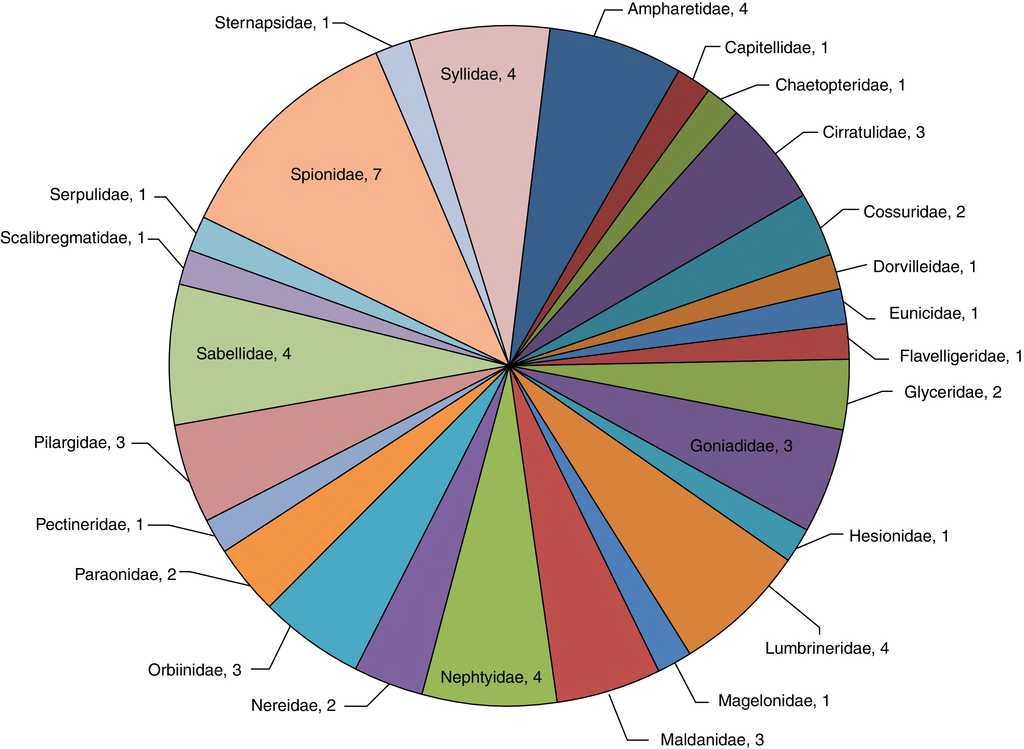
De las muestras obtenidas en esta zona se identificaron 788 organismos (21.2%), representando 26 familias y 61 especies (61.6%), donde 7 de estas corresponden a Spionidae (M. cirrifera, M. perkinsi, P. alata, P. pinnata, P. malmgreni, S. bombyx y S. kroeyeri), constituyendo el 11.5% de la diversidad específica y el 35.9% de abundancia relativa —283 organismos— de la zona, mientras que Syllidae (E. dispar, S. caeca, S. gracilis y T. aeolis), Ampharetidae (E. trilobata, M. tentaculata, M. gracilis y P. annectens), Lumbrineridae (Lumbrinereis crassidentata, Lumbrinereis erecta, Lumbrinereis latreilli y Ninoe longibranchiata) y Sabellidae (Euchone incolor, Megalomma pigmentum, Paradialychone ecaudata y Pseudopotamilla reniformis) equivalen al 26.2% de la riqueza en la zona y 17.9% de abundancia relativa —141 organismos—. Asimismo, Cirratulidae (A. parvus, C. setosa y M. tesselata), Goniadidae (Glycinde polygnatha, Goniada brunnea y Goniada maculata), Maldanidae (M. sarsi, P. gracilis y P. pacifica), Orbiinidae (Leitoscoloplos kerguelensis, Leitoscoloplos pugettensis y Scoloplos treadwelli) y Pilargidae (Ancistrosyllis jonesi, Sigambra bassi y Sigambra tentaculata) constituyen el 24.6% en el número de especies y 22.7% de abundancia relativa —179 organismos—. Sternapsidae (S. fossor), Capitellidae (Capitella sp.), Chaetopteridae (S. costarum), Dorvilleidae (Schistomeringos annulata), Eunicidae (Eunice vittata), Flabelligareidae (P. neopapillata), Hesionidae (Hesione intertexta), Magelonidae (Magelona pacifica), Pectineridae (Amphictene auricoma), Scalibregmatidae (S. inflatum) y Serpulidae (H. gracilis) corresponden al 17.9% de la riqueza específica y 13.1% del número de organismos —103 organismos— (fig. 3).
Análisis de la riqueza específica en la zona norte
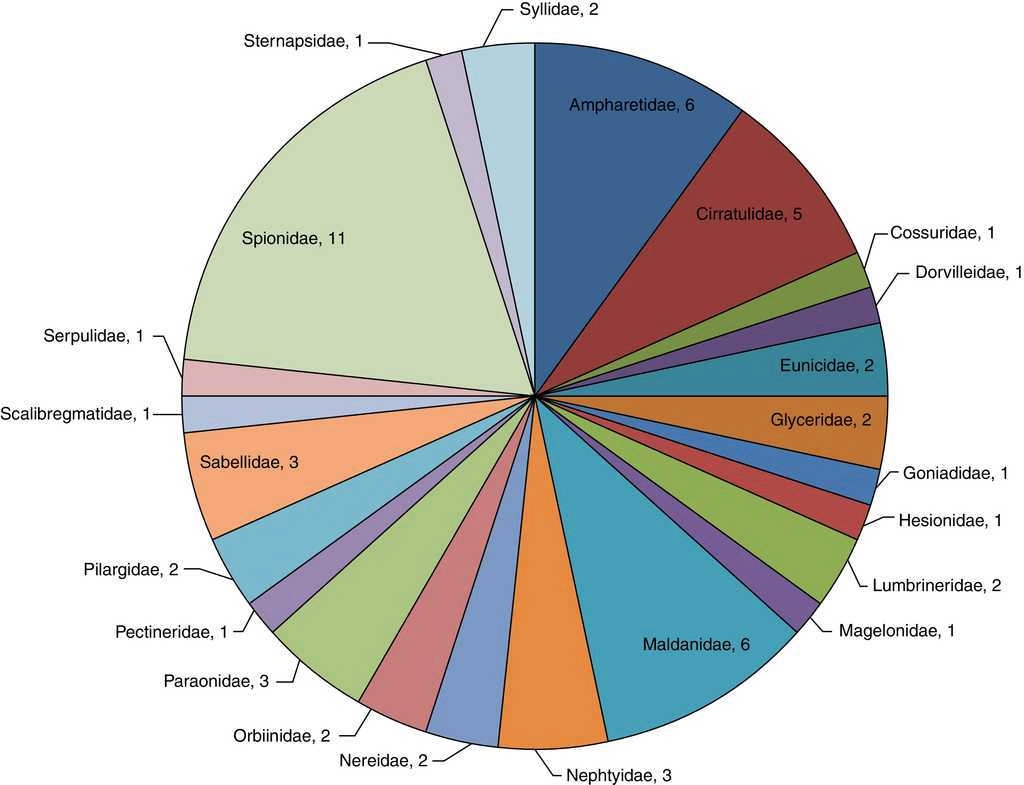
En la zona norte se identificaron un total de 592 organismos (16%) pertenecientes a 23 familias y 60 especies, donde la familia Spionidae representa el 18.3% de la riqueza específica de la zona (M. cirrifera, P. alata, P. pinnata, P. cristata, P. dubia, P. jubata, P. malmgreni, P. vermillionensis, S. bombyx, S. kroeyeri y S. wigleyi). Ampharetidae (A. scaphobranchiata, E. trilobata, L. labiata, M. oculata, M. gracilis y P. annectens) y Maldanidae (C. complanata, C. mucosa, C. gracilis, M. sarsi, P. pacifica y R. bitorquata), concentran el 20.0% de la riqueza y el 11.3% de la abundancia —67 organismos—.La familia Cirratulidae engloba 5 especies (A. monilaris, A. parvus, C. gracilis, C. setosa y M. tesselata), significando el 8.3% de la riqueza y el 10.8% de la abundancia —64 organismos—. Sternapsidae (S. fossor), Dorvilleidae (S. annulata), Hesionidae (Podarkeopsis glabra), Magelonidae (Magelona suculata), Pectineridae (Pectinaria californiensis), Scalibregmatidae (S. inflatum), Cossuridae (Cossura brunnea), Goniadidae (G. maculata) y Serpulidae (H. gracilis) resultaron con una sola especie presente en esta zona, lo que representa el 14.94% de la riqueza y el 17.1% de la abundancia —101 organismos— (fig. 4).
Análisis de la riqueza específica en la zona centro
De esta zona se identificaron un total de 2,329 organismos (62.8%) pertenecientes a 25 familias y 82 especies (82.8%). La familia Spionidae, igual que en las otras zonas, fue la mejor representada con 9 especies (M. cirrifera, M. perkinsi, P. alata, P. pinnata, P. malmgreni, P. vermillionensis, S. bombyx, S. kroeyeri y S. wigleyi) equivalentes al 11.0% de la riqueza total de la zona y el 46.8% de la abundancia —1,089 organismos—. Syllidae (E. dispar, O. brunnea, O. japonica, S. caeca, S. gracilis, T. aeolis y T. zebra) y Cirratulidae (A. monilaris, A. parvus, C. gracilis, C. corona, C. setosa, M. serratiseta y M. tesselata) están constituidas por 7 especies cada una, ambas equivalentes al 17.1% de la riqueza y 15.2% de la abundancia —354 organismos—. Maldanidae con 6 especies (C. complanata, C. mucosa, C. gracilis, M. sarsi, P. pacifica y R. bitorquata), lo que representa 7.3% de la riqueza y 7.5% de la abundancia —174 organismos—. Las familias Sternapsidae (S. fossor), Flabelligeridae (P. neopapillata), Scalibregmatidae (S. inflatum) y Serpulidae (H. gracilis) resultaron con una sola especie presente en esta zona, lo que representa 4.9% del total de la riqueza y 9.1% de la abundancia —212 organismos— (fig. 5).

Estimación global de índices con importancia biológica específica
Los resultados indican que la familia Maldanidae presentó la mayor diversidad de Shannon-Wiener con un valor de 1.76, mismo que contrasta con la obtención de un dato bajo de dominancia (0.194) y corroborado por un alto valor de equitatividad correspondiente a 0.9032. Otras familias donde también se observan datos sobresalientes de diversidad y equitatividad son Cirratulidae, Spionidae, Nephtyidae y Sabellidae. De la misma forma, las familias con valores bajos de diversidad y equitatividad, así como una alta dominancia son Dorvilleidae y Eunicidae, tal como se observa en la (fig. 6).
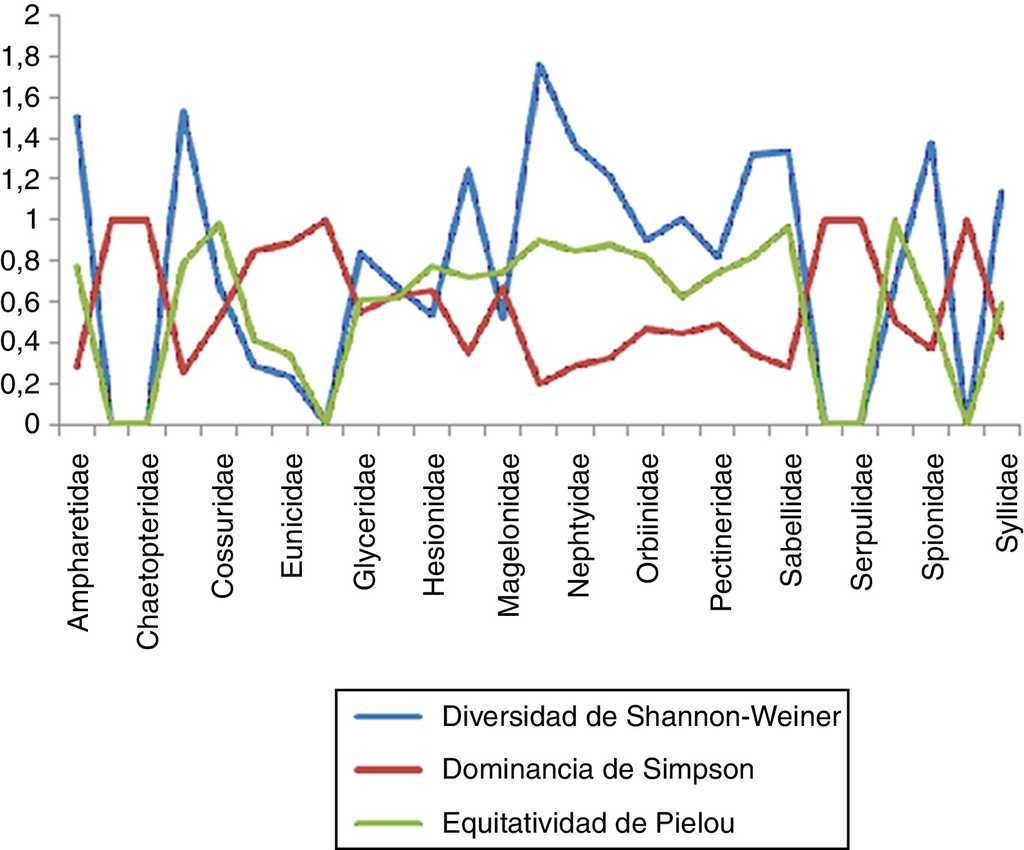
Estimación de índices con importancia biológica en la zona sur
En la zona sur se recolectaron un total de 26 familias, de las cuales Spionidae resultó la más diversa, con un total de 7 especies, y presentó un valor de 1.652, lo que nos indica una equitatividad elevada (0.849) en las 7 estaciones en las que estuvo presente y una dominancia menor (0.214). En el caso de la familia Sabellidae comprendió un total de 4 especies en 4 estaciones, evidenciando que es la segunda familia con valores de diversidad y equitatividad mayores (1.316 y 0.949, respectivamente) y una dominancia menor (0.286). Dentro de las familias que presentaron la menor diversidad se encuentran Nereididae con 2 especies en 3 estaciones, con un valor de 0.673 y una equitatividad igualmente baja (0.52); sin embargo, esto nos muestra que existe una alta dominancia entre las especies presentes (0.917). En el caso de Capitellidae, Chaetopteridae, Dorvilleidae, Eunicidae, Hesionidae, Magelonidae, Pectineridae, Scalibregmatidae, Serpulidae y Sternapsidae se obtuvieron niveles tanto de diversidad como de equitatividad iguales a 0 y una dominancia igual a 1, lo que nos indica que solo se cuenta con una sola especie para cada caso (fig. 7).

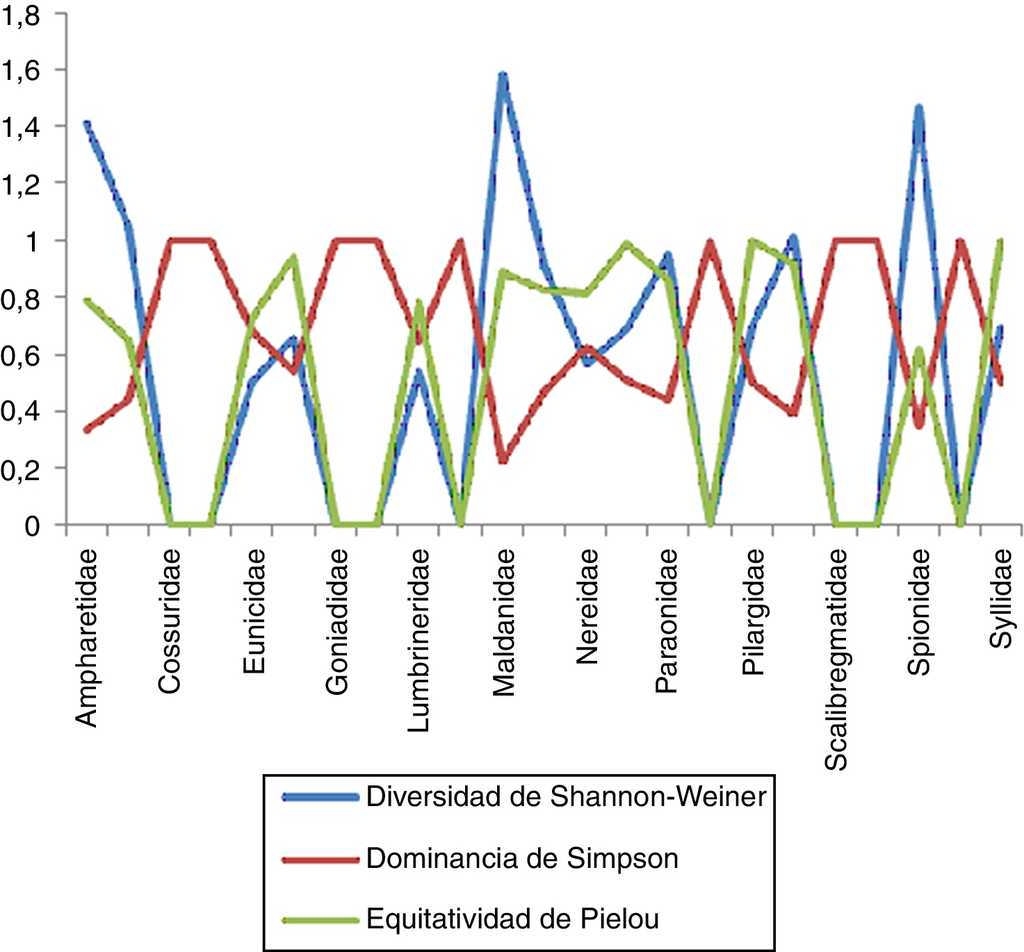
Estimación de índices con importancia biológica en la zona centro
En la zona centro se encontraron un total de 25 familias, de las cuales Cirratulidae resultó la más diversa con un valor de 1.537 y un total de 7 especies en 9 estaciones, lo que nos indica una equitatividad elevada (0.79) y una dominancia menor (0.266). En el caso de Maldanidae presentó un total de 6 especies en 8 estaciones para esta zona, mostrando que es la segunda familia con valores de diversidad y equitatividad mayores (1.421 y 0.793, respectivamente) y una dominancia menor (0.284). Dentro de las familias que presentaron la menor diversidad se encuentran Eunicidae con 2 especies en 5 estaciones, con un valor de 1.732 y una equitatividad igualmente baja (0.249); sin embargo, esto nos muestra que existe una alta dominancia entre las especies presentes (0.9201), ya que una de sus 2 especies presenta una mayor cantidad de organismos en esta zona (fig. 8). En el caso de Flabelligeridae, Scalibregmatidae, Serpulidae y Sternapsidae se obtuvieron niveles tanto de diversidad como de equitatividad iguales a 0 y una dominancia igual a 1, lo que nos indica que solo se cuenta con una sola especie de organismo para cada caso.
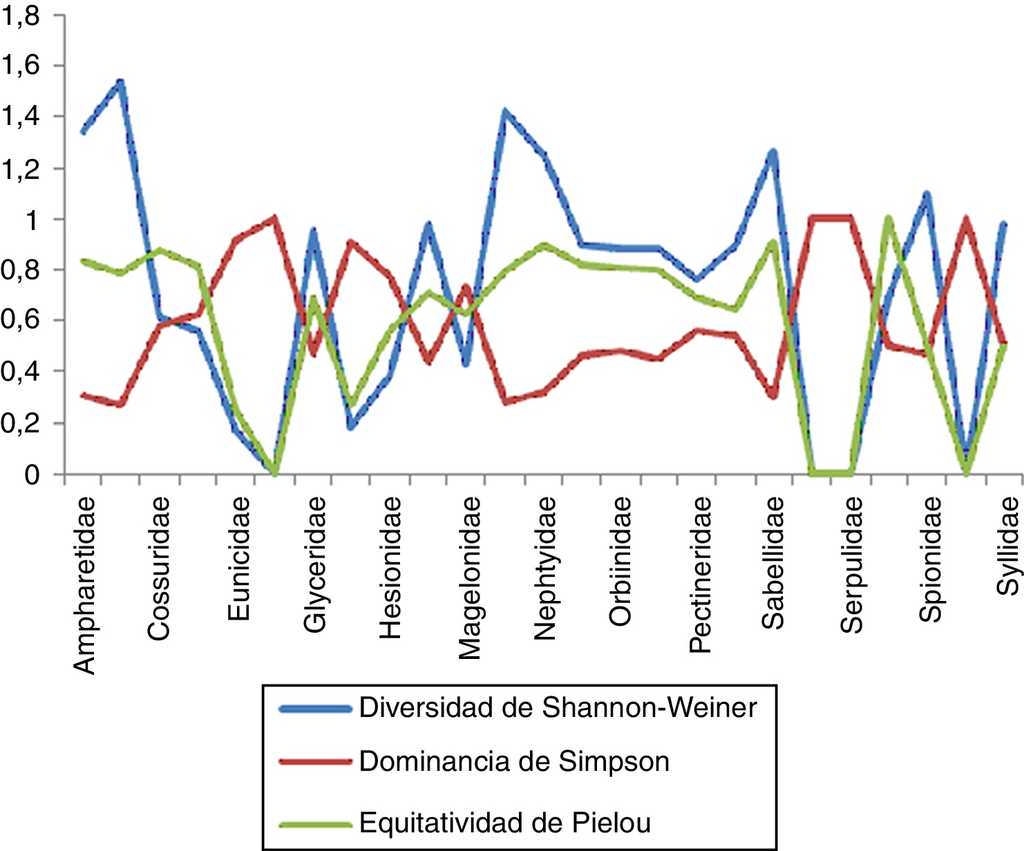
Estimación de índices con importancia biológica en la zona norte
En la zona norte se encontraron un total de 23 familias, de las cuales Maldanidae resultó la más diversa con un valor de 1.589 y un total de 6 especies en 4 estaciones, lo que nos indica una equitatividad elevada (0.886) y una dominancia menor (0.218). En el caso de Spionidae, arrojó un total de 11 especies en 5 estaciones para esta zona, mostrando que es la segunda familia con valores de diversidad y equitatividad mayores (1.466 y 0.611, respectivamente) y una dominancia menor (0.341), debido a que 3 de estas especies presentan una mayor abundancia con respecto al resto. Dentro de las familias que presentaron la menor diversidad se encuentran Eunicidae con 2 especies en 2 estaciones, con un valor de 0.500 y una equitatividad alta (0.721); sin embargo, esto nos muestra que existe una alta dominancia entre las especies presentes (0.680). En el caso de Cossuridae, Dorvilleidae, Goniadidae, Hesionidae, Magelonidae, Pectineridae, Scalibregmatidae, Serpulidae y Sternapsidae se obtuvieron niveles tanto de diversidad como de equitatividad iguales a 0 y una dominancia igual a 1, lo que nos indica que solo se cuenta con una sola especie de organismo para cada caso (fig. 9).
Análisis batimétrico con base en la estimación de índices de importancia biológica
De las 60 estaciones de muestreo durante el crucero SIMSUP VIII, en 30 se obtuvo por lo menos un ejemplar de poliqueto, donde el 53.3% corresponden a la zona centro, el 26.7% a la sur y el 20% a la norte. Asimismo, la mayor riqueza (85.9%) y abundancia (62.8%) fueron producto de recolectas realizadas en la zona centro. Cabe señalar que estas diferencias con respecto a las otras 2 zonas podrían atribuirse a que el 56% de las estaciones muestreadas fueron en esta área. En el estudio realizado por Rodríguez-Valencia (1994), equivalente a la zona centro y comparado con el de Hernández-Guevara (1994), correspondiente a la zona norte, también fue superior en riqueza específica y abundancia, tal como sucedió en el presente estudio (tabla 2). Batimétricamente, el intervalo 50-99 m para el área total y para las 3 zonas presentó la mayor riqueza y abundancia específica (tabla 3), así como la mayor diversidad estimada con base en el índice Shannon-Wiener y Simpson.
Tabla 2 Análisis batimétrico con base en la estimación de índices de importancia biológica.
| Profundidad | Especies | Número de individuos | Dominancia de Simpson | Diversidad de Shannon-Weiner | Diversidad de Simpson | Equitatividad de Pielou |
|---|---|---|---|---|---|---|
| 0-49 | 68 | 386 | 0.03801 | 3.703 | 0.962 | 0.8777 |
| 50-99 | 92 | 2,690 | 0.05317 | 3.55 | 0.9468 | 0.785 |
| 100-149 | 2 | 3 | 0.5556 | 0.6365 | 0.4444 | 0.9183 |
| 150-200 | 34 | 630 | 0.4622 | 1.599 | 0.5378 | 0.4534 |
Tabla 3 Análisis batimétrico por zona con base en la estimación de índices de importancia biológica.
| Zona | Profundidad | Especies | Número de individuos | Dominancia de Simpson | Diversidad de Shannon-Weiner | Diversidad de Simpson | Equitatividad de Pielou |
|---|---|---|---|---|---|---|---|
| 0-49 | 37 | 177 | 0.06566 | 3.107 | 0.9343 | 0.8603 | |
| Sur | 50-99 | 51 | 604 | 0.05127 | 3.352 | 0.9487 | 0.8524 |
| 100-149 | 1 | 1 | 1 | 0 | 0 | 0 | |
| 0-49 | 42 | 107 | 0.04096 | 3.447 | 0.959 | 0.9222 | |
| Centro | 50-99 | 73 | 1,592 | 0.05494 | 3.407 | 0.9451 | 0.794 |
| 100-149 | |||||||
| 150-200 | 34 | 630 | 0.4622 | 1.599 | 0.5378 | 0.4534 | |
| 0-49 | 29 | 102 | 0.09035 | 2.831 | 0.9097 | 0.8406 | |
| Norte | 50-99 | 51 | 494 | 0.09086 | 3.027 | 0.9091 | 0.7699 |
| 100-149 | 1 | 2 | 1 | 0 | 0 | 0 | |
| 150-200 |
Discusión
La costa occidental de Baja California hasta mediados de 1980 concentraba aproximadamente 236 especies de poliquetos (Salazar-Vallejo, de León-González y Salaices-Polanco, 1989). Barbosa-López (2008) llevó a cabo la revisión de las bases de datos de los registros realizados entre 1901 y 2004 en la plataforma continental de la costa occidental de Baja California, registrando un total de 491 especies, por lo que el listado elaborado en el presente estudio corresponde al 20.2% de tales especies (Anexo 1). En estudios de ciclos cortos, Rodríguez-Valencia (1994) identificó un total de 20 familias en el área correspondiente a bahía de San Juanico, en la costa occidental de Baja California Sur, de las cuales el 85% de ellas son las mismas a las encontradas en el presente trabajo. Asimismo, Hernández-Guevara (1994) identificó 23 familias de poliquetos en la bahía Sebastián Vizcaíno, donde el 88% también están presentes en esta investigación. Con el trabajo de De León-González (1994a) se obtuvo un traslape del 85% de las familias documentadas en ambos estudios.
El 64.5% de las especies de poliquetos identificadas en el presente estudio pertenecen a 10 familias: Spionidae, Maldanidae, Ampharetidae, Syllidae, Cirratulidae, Nephtyidae, Paraonidae, Pilargiidae, Lumbrineridae y Nereididae, las cuales, en su mayoría, son dominantes en sustratos blandos. Rodríguez-Valencia (1994) obtiene una estimación del 62.2% en las 10 familias con mayor riqueza, donde 7 de ellas aparecen en ambos listados, y Hernández-Guevara (1994) obtiene el 67.3% con 5 familias muestreadas en ambos trabajos. En la misma área de estudio, Barbosa-López (2008) registra que Spionidae, Paraonidae, Ampharetidae, Cirratulidae, Nereidae y Syllidae contribuyen con el 32.8% de las especies. Hernández-Alcántara (2002), en el golfo de California, también señala dentro de las primeras 10 familias con la mayor riqueza y número de organismos a Spionidae, Lumbrineridae, Nereididae, Paraonidae y Cirratulidae, mismas que representan el 27.6% de las especies.
De las 27 familias, Capitellidae, Sternapsidae, Serpulidae, Flabelligeridae, Chaetopteridae y Scalibregmatidae estuvieron representadas por una sola especie; sin embargo, Sternapsidae presentó 198 organismos y el resto, menos de 20. Comparativamente, otros estudios llevados a cabo por Rodríguez-Valencia (1994) arrojaron un total de 55% de similitud, lo que equivale a 11 familias con una sola especie. Hernández-Guevara (1994) mostró una similitud del 60%, lo equivalente a 15 familias, las cuales solo presentaron una especie. De León-González (1994a) documenta para el área de punta Eugenia hasta la punta de Cabo Falso una similitud de 39.4% o lo equivalente a 13 familias. Hernández-Alcántara (2002) igualmente registra a Sternapsidae, Serpulidae y Chaetopteridae con una sola especie para el golfo de California.
Uno de los resultados preponderantes del análisis de la poliquetofauna fueron los datos sobresalientes de riqueza y abundancia que representa la familia Spionidae, debido a que constituye el 12.2% de las 99 especies identificadas y el 42.7% de los 3,709 organismos. Información similar fue documentada por De León-González (1994a), contabilizando 10.6 y 37.8% de riqueza y abundancia, respectivamente. Igualmente, Hernández-Guevara (1994) identificó 5 especies (10.9%) de 171 organismos de esta familia (25.8%). Asimismo, Rodríguez-Valencia (1994) examinó 712 ejemplares (66.4%) pertenecientes a 4 especies. Cabe destacar que la mayor presencia de esta familia está fundamentada en la diversidad de hábitat que ocupa tanto latitudinal como batimétricamente en la plataforma continental, dominando ampliamente los fondos blandos de aguas someras y profundas, tal como es señalado por Hernández-Valdez (2008).
P. pinnata es la especie más abundante, frecuente y dominante entre las familias de poliquetos obtenidas en este estudio y representa el 24.2% del número total de organismos. En los estudios realizados por De León-González (1994a), Donath-Hernández y Loya-Salinas (1989), Encalada-Fleites y Millán-Núñez (1990) y Rodríguez-Valencia (1994), también se identificó a P. pinnata como la especie más abundante.
Del total de especies identificadas en la zona sur, la familia Spionidae fue la más diversa, así como en el trabajo de De León-González (1994a) que menciona a estos organismos sedimentívoros subsuperficiales como los más abundantes y con mayor riqueza de especies en la plataforma continental de la costa oeste de Baja California, sobre todo en sustratos blandos. La zona centro comprendió un total de 85 especies de Spionidae; nuevamente es la mejor representada, seguida por Cirratulidae. Eunicidae en esta área resultó la familia con una menor diversidad de especies. Rodríguez-Valencia (1994) menciona que esta zona presenta una acumulación de materia orgánica formando fondos blandos en su mayor parte, haciéndola homogénea y logrando el enriquecimiento del sustrato, favoreciendo así el desarrollo de un mayor número de especies con hábitos sedentarios, las cuales van desapareciendo en los estratos más profundos. En la zona norte se identificaron un total de 60 especies, de las cuales 11 pertenecen a la familia Spionidae; esta presenta la mayor diversidad al igual que en el estudio realizado por Hernández-Guevara (1994) para el área de punta María a punta Eugenia, quien menciona que el indicador de la magnitud en que los sustratos son favorables a una especie o de la adaptación de la especie a ese sustrato es la densidad de organismos por unidad de muestreo.


