










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkRevista mexicana de biodiversidad
On-line version ISSN 2007-8706Print version ISSN 1870-3453
Rev. Mex. Biodiv. vol.85 n.3 México Sep. 2014
https://doi.org/10.7550/rmb.43334
Ecología
Ecological evaluation of air and water habitats in the Great Cavern of Santo Tomás, Cuba
Evaluación ecológica de los hábitats de agua y aire en la Gran Caverna de Santo Tomás, Cuba
Janez Mulec1* and Andreea Oarga2
1 Karst Research Institute, Research Centre of the Slovenian Academy of Sciences and Arts, Titov trg 2, 6230 Postojna, Slovenia. * janez.mulec@guest.arnes.si
2 University of Nova Gorica, Vipavska 13, 5000 Nova Gorica, Slovenia. Current address: Ştefan cel Mare University, Universităţii 13, 720229 Suceava, Romania.
Recibido: 11 noviembre 2013
Aceptado: 26 marzo 2014
Abstract
Air and water habitats in the Great Cavern of Santo Tomás were investigated using cultivable indicator microbial groups. In addition, cave pools were sampled to evaluate the biomass and diversity of aquatic meiofauna. In the climatically dynamic entrance part named Cueva de las Antorchas, concentrations of total cultivable airborne bacteria ranged from 2 to 15 CFU/20 cm2/20 min, and of airborne yeasts and moulds from 2 to 23 CFU/20 cm2/20 min. Different pools fed by constant percolation water or occasional underground floods were sampled. Microbial indicators of water habitats (total bacteria, coliforms, Escherichia coli, Enterobacteriaceae) showed that microbial load very likely depended on the sampling position in the vertical transection of the cave system and closeness to the entrance. During the underground flow of Santo Tomás Stream through the cave system, the concentration of isolates typical for E. coli was reduced from 56 CFU/ml at the ponor in Valle de Santo Tomás to a concentration below the detection limit at the spring in Valle de Quemado. In some cave pools, apart from microorganisms, aquatic fauna contribute highly to the total biomass. Investigation for aquatic meiofauna diversity in cave pools revealed relative high abundances of copepods and ostracods.
Keywords: monitoring, microbial indicators, underground water, meiofauna, aerobiology.
Resumen
Se investigaron los hábitats del aire y del agua de la Gran Caverna de Santo Tomás mediante indicadores de grupos microbianos cultivables. Además, las pozas de agua dentro de la cueva fueron muestreados para evaluar la biomasa y la diversidad de meiofauna acuática. La parte de la entrada climáticamente dinámica se llama la Cueva de las Antorchas y las concentraciones totales de bacterias cultivables en el aire de esta sección oscilaron entre 2 y 15 CFU/20 cm2/20 min, y las levaduras y mohos medidos fueron de 2 a 23 CFU/20 cm2/20 min. Se muestrearon diferentes pozas alimentadas por agua de infiltración constante o por inundaciones subterráneas ocasionales. Los indicadores microbianos de hábitats acuáticos (bacterias totales, coliformes, Escherichia coli, enterobacterias) mostraron que la carga microbiana muy probablemente depende de la posición de muestreo relativa al corte transversal vertical del sistema de la cueva y de la cercanía a la entrada. Durante el flujo subterráneo de la corriente de agua de Santo Tomás a través del sistema de la cueva, la concentración de los aislamientos típicos para E. coli se redujo de 56 CFU/ml en el ponor en Valle de Santo Tomás a una concentración por debajo del límite de detección en el ojo de agua de Valle de Quemado. En algunas pozas, además de los microorganismos, existe una contribución de la fauna acuática a la biomasa total. La investigación de la diversidad acuática de meiofauna en las pozas reveló una alta abundancia relativa de copépodos y ostrácodos.
Palabras clave: monitoreo, indicadores microbianos, agua subterránea, meiofauna, aerobiología.
Introduction
In Cuba, karst occupies around 66% of the land surface (Núñez-Jiménez, 1984; Molerio-Leόn and Parise, 2009; Febles-González et al., 2012), and in some regions karst aquifers represent an important source of water for human consumption and agriculture (Molerio-León and Gutiérrez-Díaz, 1999). The semi-tropical climate of the Caribbean region, accompanied by high organic input in karst underground, is reflected in high turnover rate of organic matter. The entrance sections and occasionally deeper parts of such caves, frequently shelter different trogloxenic animals such as: snakes, scorpions, frogs, crabs, crickets and spiders. In the western part of Cuba, the Great Cavern of Santo Tomás is a complex cave system from which local people use water and other natural resources for their needs (Núñez-Jiménez, 1990) and it harbours an obligate subterranean biota (Pérez-González and Yager, 2001).
Air and water were sampled in The Great Cavern of Santo Tomás. Atmospheric conditions and airborne microbes were evaluated in the climatically dynamic entrance section (Cueva de las Antorchas and Cueva de la Incógnita). Cave aerobiology investigation was conducted in this cave system because air has been poorly investigated in the Caribbean region, to evaluate conditions and organic load in this habitat, and to emphasize air as a vehicle for transport of airborne microbial pathogens. In Cuban caves spores of the human and bat pathogen Histoplasma capsulatum are frequently encountered (Font D'Escoubet and Macola Olano, 1976; Fernández Andreu, 1988; Erkens et al., 2002; Craven, 2013).
The same microbial indicators used for the air study were applied for water habitats. Microbiological indicator groups, such as cultivable heterotrophic aerobic bacteria, Escherichia coli, coliform bacteria, enterobacteria, and fungi are of great help in assessing the environmental health of underground ecosystems (Mulec et al., 2012a). Information on these parameters is even more significant in warm caves in tropical and subtropical zones that are rich in organic material. Microbes related to the environmental health of underground habitats are airborne and originate from cave sediments, guano and external air, and waterborne which originate from surface/underground river and/or percolating water (Borda et al., in press; Mulec et al., 2012a). Water samples were collected from pools and active river flow to observe eventual differences related to the complex water pathway due to the multi-level nature of the Great Cavern of Santo Tomás. To get a better overview of biomass in pools containing captured water after floods and in pools with percolation water, in addition to concentration of bacterial indicators, the abundance of aquatic fauna was also estimated. In comparison to temperate caves, higher turnover rates of organic matter and more dynamic relations between biota are expected in tropical and subtropical caves due to generally higher temperatures. Results on water quality are important, as the local population uses water for drinking from karst underground. This study represents an incentive for wider use of cave monitoring in the Caribbean region, because it is generally poorly studied (Day and Koenig, 2002; Parise and Valdes-Suarez, 2005).
Material and methods
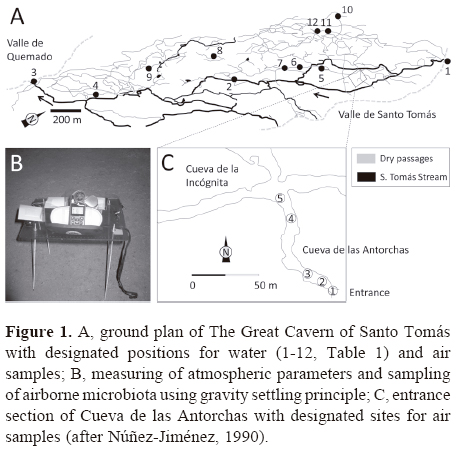
Description of the cave. The great cavern of Santo Tomás in Sierra de Quemado, Cuba (in Spanish literature found under the following names: Caverna de Santo Tomás, Cueva de Santo Tomás and La Gran Caverna de Santo Tomás, Fig. 1A), extends for approximately 46 km of dry and flooded subterranean passages at 7 main levels (I-VII) and 65 m of vertical distance in a mogote, a residual karst hill. Since 1994 a part of the cave system named Cueva de las Avispas and Cueva de las Perlas (VI level) was equipped for tourist visits, but without electric lighting. Local people use water from the cave for their domestic needs, and bat guano to fertilize tobacco fields in the foothills of surrounding mogotes. This complex karst system has been recognized as a Cuban National Monument due to its natural, historical, paleontological and archaeological importance (Núñez-Jiménez, 1990; Parise and Valdes-Suárez, 2005). Sampling was limited to the unrestricted part of the cave system.
Microbiological media. RIDA®COUNT plates were used to enumerate different microbial groups, RIDA®COUNT Total Aerobic Count for total heterotrophic aerobic bacteria, RIDA®COUNT E. coli/Coliform for total coliform bacteria with discernment of E. coli colonies, RIDA®COUNT Enterobacteriaceae for enterobacteria, and for yeasts and moulds RIDA®COUNT Yeast&Mold Rapid. For airborne microbiota, approximately 2 hours before caving, RIDA®COUNT media sheets were activated by adding 1 millilitre of sterile physiological saline (R-Biopharm AG, Germany). Isolates that exhibited ß-D-glucuronidase and ß-D-galactosidase biochemical activities on RIDA®COUNT E. coli/Coliform plates were considered indicative of E. coli (R-Biopharm AG, Germany). After inoculation of plates in a cave, plates for bacteria were placed in an incubator at 37° C for 24 hours, and plates for yeasts and moulds were incubated at room temperature (~25° C) for 48 hours. After cultivation, microbial colonies grown from water samples were counted and expressed as Colony-Forming Units (CFU) per millilitre, and from the air samples as CFU/20 cm2/20 min ( Mulec et al., 2012a, 2012b).
Air samples. Airborne microbes were sampled using a depositional sedimentation principle by exposing open plates to the cave atmosphere for 20 minutes. Sampling of airborne microbiota was done using a 30 cm high rack above the cave floor to avoid direct contact with cave sediment and dust formation by the research team, especially in the dry part of the cave (Fig. 1B). The biggest changes in air conditions were expected in climatically diverse cave parts, e.g. at the entrance and inside in narrow and ramified passages. Air samples were collected at various distances from the cave entrance: in Cueva de las Antorchas (III) approximately at 5, 15, 25, and 100 m depths in the cave, and at the beginning of Cueva de la Incógnita (III) at 120 m from the entrance (Fig. 1C). 20-minute samplings of airborne microbiota (22 December 2011) were done at 5 sites progressively (12:15-12:35, 12:43-13:03, 13:11-13:31, 13:38-13:58, 14:08-14:28) from the site closest to the entrance towards the interior of the cave. Simultaneously with microbiota sampling, temperature, relative humidity (RH) and wind were recorded by a Kestrel 4500 Pocket Weather Tracker datalogger (USA).
Water samples. For microbial analyses of water samples, plates were aseptically inoculated in the cave with 1 millilitre of water. Due to cave morphology, different parts of the cave system were selected for water samples than those for air samples. At 4 levels (I-IV), altogether 12 water bodies were sampled in December 2011. Different types of water bodies were selected to estimate microbial load: i) Santo Tomás Stream (Arroyo de Santo Tomás) at the ponor at Valle de Santo Tomás (sample no. 1), in the cave (no. 2) and at the spring at Valle de Quemado (no. 3); ii) pools containing captured water after floods caused by heavy rains in Cueva de las Represas (II), and iii) pools with percolation water in Cueva de la Incógnita (III), Cueva de Rivero (III), Cueva del Salón (III), Cueva Escarlata (IV), and Cueva Increíble (IV) (Table 1).
Temperature and specific electric conductivity (SEC) were measured with a WTW MultiLine P4 (Germany) and pH was estimated with pH indicator strips (EMD Chemicals, Germany).
Samples for meiofauna were taken from all selected pools after microbiological sampling was finished. Water (4-8 litres) was filtered through a bottle equipped with 60 μm mesh on 2 sides which allowed water to pass through, but retained organisms (Pipan, 2005; Oarga, 2006; Iepure and Oarga, 2011). The volume of filtered water from pools was used to estimate abundance of individuals per sample (Table 1). Samples of organisms were preserved in 80% ethanol. Animals were sorted and identified up to the class and/or order level for Crustacea and up to the order level for Rotifera under a stereomicroscope Nikon SMZ800 (USA).
Results
Air. Measured atmospheric parameters in the entrance part of the Great Cavern of Santo Tomás in Cueva de las Antorchas showed changing conditions related to cave morphology and clearly indicated impact from the surface. The highest temperature and the lowest RH were measured at the site closest to the entrance (Figs. 2A, C). Later successive measurements toward the interior showed that cave temperature was progressively lower, and RH increased. At the last sampling site (14:08-14:28) approximately 120 m from the entrance, RH reached 95% and temperature 22.7° C. This temperature is very close to the average temperature of the measured water bodies, 21.5° C (Table 1).
Inflow of external air at the cave entrance dried the cave air. In narrow parts, air currents reached velocities of up to 2.5 m/s (Fig. 2B, site no. 3). Deeper in the cave, air flow was less strong, and at the branching of cave passages after Cueva de las Antorchas the wind was diminished (Fig. 2B).
At 3 sampling sites close to the cave entrance, the total concentrations of fungi were higher compared to total bacterial concentrations, but at the 2 sites deeper in the cave we found the opposite trend (Fig. 2D). No airborne coliforms, E. coli and enterobacteria were detected on differential and selective media, respectively.
Water. SEC of Santo Tomás Stream before sinking (271 µS/cm) and during its underground water course (275 µS/cm) was lower than at the spring (398 µS/cm). Captured water in a pool had a comparable value of SEC (220 µS/cm) to the water from pools that were fed by active drips (199 to 330 µS/cm). pH of water samples was neutral to somewhat alkaline (7.0-7.5), and temperature was in the range of 20.6 to 22° C (Table 1).
Santo Tomás Stream (I) sank into the Great Cavern of Santo Tomás with rather high concentrations of total cultivable bacteria (910 CFU/ml) which were reduced during underground water course approximately 4 fold (217 CFU/ml) at the spring at Valle de Quemado (Fig. 3). The concentration of E. coli, an indicator of fresh faecal contamination, in Santo Tomás Stream was reduced from 56 CFU/ml at the ponor to a concentration below the detection limit at the spring (Fig. 3). All investigated pools in the cave system were negative for the presence of E. coli, except a pool with captured water in Cueva de las Represas (sample no. 4) with 1 isolate indicative of E. coli. In 2 pools (no. 5 and no. 6) in Cueva de la Incógnita (III) low concentrations of total bacteria were found, 70 and 54 CFU/ml, respectively, and no coliforms; but in another pool (no. 7), concentration of total heterotrophic bacteria was much higher (276 CFU/ml) and, in addition, this sample contained 6 CFU/ml of coliforms. The pool from Cueva de Rivero (no. 8) had 57 CFU/ml of total bacteria and 10 CFU/ml of coliforms. The pool from Cueva del Salón (no. 9) contained similar concentrations, 41 CFU/ml of total bacteria and 18 CFU/ml of coliforms. In water samples from pools from Cueva Escarlata (IV) and Cueva Increíble (IV) no cultivable bacteria were detected. When detected, the concentrations of coliform bacteria were comparable to the concentrations of Enterobacteriaceae (Fig. 3).
In 9 pools sampled for meiofauna, the dominant taxonomic group was Ostracoda (Crustacea), while Cyclopoida (Crustacea, Copepoda) were present in 3 samples, and Harpacticoida (Crustacea, Copepoda) and Rotifera (Bdelloida) (Miloslav Devetter, personal communication) in one sample (sample no. 8). Sample no. 8 –Cueva de Rivero (III)– was located approximately 10 m from one of the entrances into the cave system and was evidently enriched with cave sediment. In addition to this site, copepods with numerous nauplii were found in pools in Cueva de las Represas (II) and Cueva del Salón (III). Ostracoda, as the only faunal representative was identified in 4 sites in Cueva de la Incógnita (III) and Cueva Increíble (IV). Two sampled pools in Cueva Escarlata (IV), and Cueva Increíble (IV) contained no fauna and no cultivable bacteria in 1 millilitre of samples (Figs. 3, 4).
Pearson correlation analyses were applied among faunal concentration and temperature, SEC and microbial indicator groups, and between total bacterial concentrations and physical parameters of water bodies. Relations between faunal concentrations and all parameters showed a negative trend (r< 0) and no statistical significance (p> 0.05). r2 for faunal abundance vs. SEC was 0.061, vs. temperature was 0.301, vs. pH was 0.092, vs. total bacteria was 0.030, vs. E. coli was 0.035, vs. coliforms was 0.005, and finally, faunal abundance vs. enterobacteria was 0.009. A similar trend (r< 0, p> 0.05) was observed for the relationship between total bacterial counts and measured parameters; r2 for total bacterial counts vs. SEC was 0.007, vs. temperature was 0.023, and total bacterial counts vs. pH was 0.207.
Discussion
Air. Climate in caves is very much subject to external influences (Sánchez-Cañete et al., 2013) and is very pronounced in the Great Cavern of Santo Tomás which is characterized by a complex multi-levelled and ramified cave system. One can expect that wind velocities may exceed 2.5 m/s which were measured in the cave during warm and sunny weather. Wind mixes cave air, and is a carrier for airborne external and cave-originating minute particles (Mulec et al., 2012c). By drying the cave atmosphere at the entrance, and mixing airborne particles, wind is important also for aerosolization of biological particles. This is important especially for aerosolization of H. capsulatum and other pathogenic microbes which can be inhaled during a cave visit. H. capsulatum is a pathogenic fungus that inhabits sediments and guano in tropical caves, and is a causative agent of histoplasmosis which can be manifested as acute benign pulmonary, acute disseminated, chronic pulmonary, chronic localised and chronic disseminated infections (Richardson and Warnock, 2003). Few unreported cases of histoplasmosis among cavers were attributed to speleological activities in this cave (Galliano Bressan, pers. comm.). A case of histoplasmosis probably acquired during this caving expedition was linked to high concentration of minute particles in the entrance and improper personal protection (Nicolò Zuffi, pers. comm.). However, wearing protection masks in the Cuban caves may prevent histoplasmosis even with high infectious doses (Erkens et al., 2002). Proper personal protection, e.g. masks, avoiding dust formation, proper disposal of caving equipment and sanitation, should be seriously considered, especially in those parts of the cave where morphology enhances wind formations and deposits of organic sediments and bat guano are present. Disturbance or mixing of organic material in caves, especially guano should be avoided as it was recently shown using RIDA®COUNT plates that up to 3.35% of fungi and up to 0.10% of bacteria can be directly aerosolized from guano (Borda et al., in press).
The ratio of total bacterial counts vs. total fungal counts was between 0.25 and 0.39 during the first 3 measurement campaigns, but the fourth (13:38-13:58) had 2.50 and the last (14:08-14:28) 2.00. Relatively higher concentrations of bacteria compared to fungi might be attributed to freshly introduced organic material, for example by human and/or animal excrement. Generally, in caves with no evident fresh organic material, counts of cultivable airborne fungi exceed those of bacteria (Mulec et al., 2012a). Bacteria/fungi ratio during the first 3 measurements, when strong external influence prevailed, was comparable to the ratios from moderate eutrophic continental caves from Slovakia (0.18-0.33) (Mulec et al., 2012b). Concentration of airborne bacteria and fungi in the Great Cavern of Santo Tomás were similar in range as in temperate Slovenian and Slovakian caves (Mulec et al., 2012a, 2012b); however due to strong wind in the entrance section, the concentration of airborne microbes very likely did not reveal the true values as the wind caused deviation from normal vertical sedimentation route of air particles and pushed minute particles deeper into the cave. Wind in this and other Caribbean caves should be considered as a potential vehicle for transmission of pathogenic microbes, e.g. H. capsulatum. This is one reason why tourist visits should be carefully planned for each individual cave sector.
The constant input, mixing, circulation, and final sedimentation of cave allochthonous and autochthonous material from the air represent an important and continuous "rain" of organic carbon for cave fauna (Mulec et al., 2012c). Apart from dripping water and river flow, this is another important way to introduce energy source and microbes in caves.
Water. Water pathways in the Great Cavern of Santo Tomás are complex, as vertical water flow is intercepted by many levels. Screening of water quality in pools fed by percolating and captured water by using microbial indicators showed that microbial load diminished from the lower towards the intermediate levels of the cave. Detection of organic load measured as concentration of heterotrophic microbes within the system depended on the sampling position in the vertical transection and distance from the exterior. Water samples from pools that were fed solely by percolating water contained no E. coli. A pool which contained captured water after rain close to one of the entrances in Cueva de las Represas in the vicinity of a walking path (sample no. 4) did contain E. coli. During the underground flow of Santo Tomás Stream, the concentration of E. coli was reduced (Fig. 3). Due to the presence of isolates which indicated E. coli and Enterobacteriaceae on RIDA®COUNT plates, direct uptake of water for drinking from the Santo Tomás Stream represents a biohazard. The ratio of coliform bacteria vs. total bacteria in Santo Tomás Stream dropped after approximately 2.0 km of underground flow from 0.37 to 0.22. This value is comparable to the ratio of coliforms vs. total bacteria after 9 km of underground river flow of the Pivka River in the cave system Postojna Cave in the temperate zone in Slovenia (Mulec et al., 2012b). Using microbial indicator groups for monitoring of water bodies, in particular those of the Santo Tomás Stream, is recommended, especially if water is to be used for human consumption. Higher temperature in the underground river might cause higher organic matter turnover compared to underground rivers in moderate climate, nevertheless one has to take into account also eventual dilution in the underground.
When studying microbiology in the underground water bodies, one should also consider the fauna. Microbes are a food source for meiofauna. An analysis of microbial and faunal abundance from different sampling sites obtained during one expedition in the Great Cavern of Santo Tomás indicated that high abundance of fauna was accompanied by relative lower microbial concentrations (r= -0.172, Fig. 4). Relationship between concentrations of microbiota and meiofauna abundance in pools is not expected to be simply linear and directed only by faunal grazing on microbes, as there are many factors which define conditions in pools, e.g. presence of organic matter, hydrodynamics, and various ecological interactions. Continuous input into pools and other underground voids with microbes and organic matter is an important source of nutrients. For example, in the temperate Škocjan Caves, Slovenia (~12° C), with an assumed constant dripping rate from only one trickle, approximately 4.3 × 106 bacterial CFU/year is introduced (Gerič et al., 2004). Organic load measured solely by the microbial indicator groups in underground pools may mask the true situation. A more realistic estimation of organic load should also include other macronutrients such as C, N, P and S.
In addition to microbes, aquatic fauna contributes to the total biomass of cave pools to a greater or lesser extent (Fig. 4). Although the samples did not show high faunal richness at superior taxonomic levels, a high diversity of species level can be expected for such a big and heterogeneous cave system. The relative high number of nauplii in samples no. 4 (Cueva de las Represas, II) and no. 9 (Cueva del Salón, III) indicated favourable conditions for reproduction of copepods. Copepods can be used as an important parameter to characterize conditions in the epikarst, its spatial connectivity and responses to hydrological events (Pipan and Culver, 2007a, 2007b). Pools that are fed by dripping and seeping water from cave ceilings form natural traps for epikarst fauna, of which copepods are most abundant (Pipan, 2005). Ostracods are also common cave-dwellers in the vadose zone; however due to morphological characteristics and locomotion behaviour they prefer standing water pools or interstitial sediments of subterranean rivers (Peterson et al., 2013). Identification at the species level has not been done yet; it is likely that the crustaceans found (Copepoda, Ostracoda) can be highly diverse as was indicated by previous attempts on Cuban cave crustacean diversity (Botosaneanu, 1970; Pleşa, 1981; Iepure and Defaye, 2003). Spatially isolated mogotes might play an important role in development of novel lineages of strictly obligate cave organisms.
In pools in the Great Cavern of Santo Tomás other groups were also found including those that have been washed from the surface and soil. For example, in the level VI (Cueva de las Perlas) of the cave system in 2009 a sample of water pool contained individuals of the polychaete group determined as Aeolosoma sp., Aeolosomatidae (Christer Erséus, pers. comm.).
Results of the study showed that due to the high complexity of the Great Cavern of Santo Tomás cave system, different monitoring plans related to protection and conservation should be undertaken for different parts. The approach presented in this study for air and water habitats can serve as an additional example of a low-cost monitoring plan to estimate human and natural impact in Cuban and other Caribbean caves, especially those that serve as water sources for people and as attractions for tourists. For a better understanding of trophic levels in the subterranean environment in (sub) tropical zones, studies should include microbiological, faunistic and environmental data.
Acknowledgements
The study was supported by the Research Programme P6-0119 "Karst Research" and project "Promotion of Slovenian Science" MU-PROM/2011-2-003 financed by the Slovenian Research Agency. Authors are grateful to Galliano Bressan, Héctor Pérez Jiménez and members of Sociedad Espeleológica de Cuba and Escuela Nacional de Espeleología "Antonio Núñez Jiménez" for help during field work; to Christer Erséus for identification of Aeolosoma, Miloslav Devetter for identification of Bdelloida, Sanda Iepure for useful comments on earlier version of the manuscript, and Trevor Shaw and Laura Rosales Lagarde for language assistance.
Literature cited
Borda, D. R., R. M. Năstase-Bucur, M. Spînu, R. Uricariu and J. Mulec. In press. Aerosolized microbes from organic rich materials: case study of bat guano from caves in Romania, Journal of Cave and Karst Studies.
Botosaneanu, L. 1970. L'expédition biospéologique à Cuba (mars-juin 1969) organisée par les Académies des Sciences de Cuba et de Roumanie: presentation sommaire des stations explores pur le prélèvement d'échantillons de faune aquatique souterraine. Travaux de L'Insitut de Speologie "Emile Racovitza" 9:81-95. [ Links ]
Craven, S. A. 2013. The epidemiology of acute benign pulmonary histoplasmosis (= cave disease). Cave and Karst Science 40:133-140. [ Links ]
Day, M. and S. Koenig. 2002. Cave monitoring priorities in Central America and the Caribbean. Acta Carsologica 31:123-134. [ Links ]
Erkens, K., M. Lademann, K. Tintelnot, M. Lafrenz, U. Kaben and E. C. Reisinger. 2002. Histoplasmosis group disease in bat researchers returning from Cuba. Deutsche Medizinische Wochenschrift 127:21-5. [ Links ]
Febles-González, J. M., M. B.Vega-Carreño, A. Tolón-Becerra and X. Lastra-Bravo. 2012. Assessment of soil erosion in karst regions of Havana, Cuba. Land Degradation and Development 23:465-474. [ Links ]
Fernández-Andreu, C. M. 1988. Isolation of Histoplasma capsulatum in bats in Cuba. Revista Cubana de Medicina Tropical 40:36-43. [ Links ]
Font D'Escoubet, E. and S. Macola Olano. 1976. Histoplasma capsulatum: isolation from bats in Cuba. Revista Cubana de Medicina Tropical 28:119-25. [ Links ]
Gerič, B., T. Pipan and J. Mulec. 2004. Diversity of culturable bacteria and meiofauna in the epikarst of Škocjanske jame Caves (Slovenia). Acta Carsologica 33:301-309. [ Links ]
Iepure, S. and D. Defaye. 2003. Redescription of Teratocyclops cubensis Pleşa, 1981 (Copepoda, Cyclopoida, Cyclopidae) from Cuba. Crustaceana 76:591-604. [ Links ]
Iepure, S. and A. Oarga. 2011. A new Acanthocyclops Kiefer, 1927 (Copepoda: Cyclopidae) from caves in Apuseni Mountains (North-Western Romania). Annales Zoologici 61:427-438. [ Links ]
Molerio-León, L. M. and J. Gutiérrez-Díaz. 1999. Agricultural impacts on Cuban karstic aquifers. In Karst hydrogeology and human activities, D. Drew and H. Hötzl (eds.). Balkema, Rotterdam. p. 76-78. [ Links ]
Molerio-Leόn, L. and M. Parise. 2009. Managing environmental problems in Cuban karstic aquifers. Environmental Geology 58:275-283. [ Links ]
Mulec, J., V. Krištůfek and A. Chroňáková. 2012a. Monitoring of microbial indicator groups in caves through the use of RIDA®COUNT kits. Acta Carsologica 41:287-296. [ Links ]
Mulec, J., V. Krištůfek and A. Chroňáková. 2012b. Comparative microbial sampling from eutrophic caves in Slovenia and Slovakia using RIDA®COUNT test kits. International Journal of Speleology 41:1-8. [ Links ]
Mulec, J., J. Vaupotič and J. Walochnik. 2012c. Prokaryotic and eukaryotic airborne microorganisms as tracers of microclimatic changes in the underground (Postojna Cave, Slovenia). Microbial Ecology 64:654-67. [ Links ]
Núñez-Jiménez, A. 1984. Cuevas y Carsos. Editora Militar, La Habana. 431 p. [ Links ]
Núñez-Jiménez, A. 1990. La Gran Caverna de Santo Tomás: Monumento Nacional. Plaza Vieja, La Habana. 160 p. [ Links ]
Oarga, A. 2006. Groundwater faunal assemblages in different habitats of Padurea Craiului Mountais (NW Romania). MSc thesis, Babes-Bolyai University. Cluj-Napoca, 63 p. [ Links ]
Parise, M. and M. Valdes-Suarez. 2005. The show cave at "Gran caverna de Santo Tomás" (Pinar del Rio province, Cuba). Acta Carsologica 34:135-149. [ Links ]
Pérez-González, A. and J. Yager. 2001. The Cuban troglobites. In Karst Water Institute Special Publication 6, D. C. Culver, L. Deharveng, J. Gibert and I. D. Sasowsky (eds.). Charles Town, WV p. 61-75. [ Links ]
Peterson, D. E., K. L. Finger, S. Iepure, S. Mariani, A. Montanari and T. Namiotko. 2013. Ostracod assemblages in the Frasassi Caves and adjacent sulphidic spring and Sentino River in the Northeastern Apennines of Italy. Journal of Cave and Karst Studies 75:11-27. [ Links ]
Pipan, T. 2005. Epikarst - a Promising Habitat: Copepod Fauna, its Diversity and Ecology: a Case Study from Slovenia (Europe). ZRC Publishing, Ljubljana. 101 p. [ Links ]
Pipan, T. and D. Culver. 2007a. Copepod distribution as an indicator of epikarst system connectivity. Hydrogeology Journal 15:817-822. [ Links ]
Pipan, T. and D. Culver. 2007b. Epikarst communities: biodiversity hotspots and potential water tracers. Environmental Geology 53:265-269. [ Links ]
Pleşa, C. 1981. Résultats des expeditions biospéleologique Cubano-Roumaines à Cuba III: Cyclopides (Crustaceda, Copepoda) de Cuba. Editura Academiei Republicii Socialiste Romania 3:17-34. [ Links ]
Richardson, M. and D. Warnock. 2003. Fungal Infection: Diagnosis and Management. Blackwell Publishing, Oxford. 366 p. [ Links ]
Sánchez-Cañete, E. P., P. Serrano-Ortiz, F. Domingo and A. S. Kowalski. 2013. Cave ventilation is influenced by variations in the CO2-dependent virtual temperature. International Journal of Speleology 42:1-8. [ Links ]

