










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkRevista mexicana de biodiversidad
On-line version ISSN 2007-8706Print version ISSN 1870-3453
Rev. Mex. Biodiv. vol.83 n.2 México Jun. 2012
Taxonomía y sistemática
Two new species of the bee genus Peponapis, with a key to the North and Central American species (Hymenoptera: Apidae: Eucerini)
Dos especies nuevas de abejas del género Peponapis, con una clave para las especies de América del Norte y Central (Hymenoptera: Apidae: Eucerini)
Ricardo Ayala1* and Terry Griswold2
1 Estación de Biología Chamela, Instituto de Biología, Universidad Nacional Autónoma de México. Apartado Postal 21, 48980 San Patricio, Jalisco, México. *rayala@ibiologia.unam.mx.
2 USDA–ARS, Bee Biology and Systematics Laboratory, Utah State University, Logan, UT 84322–5310, USA.
Recibido: 12 octubre 2011
Aceptado: 20 enero 2012
Abstract
Two new species of squash bees, Peponapis pacifica Ayala and Griswold sp. n. and P. parkeri Griswold and Ayala sp. n., are described and illustrated. Peponapis pacifica is oligolectic on flowers of Schizocarpum longisepalum (Cucurbitaceae) endemic to Mexico, where it is found in the tropical dry forest along the Pacific Coast, between Sonora and Chiapas and in the Balsas River basin; and P. parkeri is known only from the Pacific slope of Costa Rica. A key for the North and Central American species of Peponapis is provided.
Key words: taxonomy, Hymenoptera, Apidae, squash bees, Cucurbitaceae, Schizocarpum.
Resumen
Se describen e ilustran 2 especies nuevas de abejas, Peponapis pacifica Ayala and Griswold sp. n. y P. parkeri Griswold and Ayala sp. n. De éstas, P. pacifica es oligoléctica sobre flores de Schizocarpum longisepalum (Cucurbitaceae), endémica de México, con distribución asociada al bosque tropical caducifolio a lo largo de la vertiente del Pacífico entre Sonora y Chiapas y en la Cuenca del río Balsas; P. parkeri es conocida sólo para la vertiente del Pacífico de Costa Rica. Se incluye una clave para las especies de América del norte y central.
Palabras clave: taxonomía, Hymenoptera, Apidae, abejas de calabazas, Cucurbitaceae, Schizocarpum.
Introduction
Revisionary studies of bees of the genus Peponapis (Hurd and Linsley, 1964, 1966, 1967, 1970) confirm Mexico as the center of Peponapis diversity with numbers diminishing north to Canada and south into southern South America (Michener, 2007). The genus is exclusive to America, with 15 known species (including 2 herein described), of which 11 are present in North America, all in Mexico (Ayala et al., 1996), 7 in Central America with 2 in Costa Rica, and only 3 for South America, 1 reaching Argentina (Table 1; Hurd and Linsley, 1964, 1966, 1967; Moure et al., 2007).
Hurd and Linsley (1970) present data on the importance of the species of Peponapis Robertson and Xenoglossa Smith as specialist pollinators of the flowers of Cucurbita, including cultivated and wild species of squashes (Cucurbita), plants that also have an important center of diversification in Mexico (Merrick and Bates, 1989; Lira–Saade, 1995). The relation between the distribution of Peponapis and Cucurbita, and the analysis of probable original distributions of these bees and plants, previous to the dispersal of squashes by domestication, is presented by Giannini et al. (2010). Most of the known species of Peponapis are crepuscular; their activity occurs at dusk and resumes in the early hours of the morning, in synchrony with the anthesis of Cucurbita flowers. Many of the species of this genus also visit flowers of Ipomoea (Convolvulaceae), plants that may be an important nectar source (Hurd and Linsley, 1964). These bees play a critical role as unmanaged pollinators, providing important ecosystem services (Klein et al., 2007). Concern has been raised that these pollinator services may be in jeopardy (Mayer et al., 2011). Therefore, it is necessary to emphasize their importance and to develop strategies for their conservation.
Here we describe 2 new species and provide a key to the species of North and Central American Peponapis, modified from Hurd and Linsley (1966).
Materials and methods
The specimens from Mexico used in the descriptions were collected during a study of the bee fauna of Chamela (Ayala, 1989, 2004). Subsequently, the PCAM project (Programa Cooperativo sobre la Apifauna Mexicana) provided additional specimens that expanded their distribution in Mexico. Morphological terminology follows Michener (2007). In the description the following abbreviations are used: F, flagellar segment; T, terga (um); S, sterna (um). To illustrate the genitalia, the structure was placed in 10% KOH solution for approximately 24 hrs, then rinsed in distilled water and dehydrated with 90% ethanol, before being transferred to glycerin for study and illustration. Genitalia were stored in glycerin in a microvial attached to the pin of the specimen. Photomicrographs were taken using an Olympus SZH10 microscope and a Keyence® VHX–500F Digital Imaging System.
Peponapis pacifica Ayala and Griswold sp. n. (Figs. 1–6, 7–10)
Peponapis sp. Ayala, 1989: 403, 430
Peponapis sp. n. Ayala, 2004: 201
Diagnosis: Males (Fig. 1) are easily separable from those of other Peponapis species by the following characteristics: labrum with distinct notch on distal margin (Fig. 5) that is lacking in other species; median flagellar segments crenulate (Fig. 3); prominent tuft of dark brown or black hairs basoventrally on hind tibia (Fig. 4); and distinctive shape of S6 with posterior medial projection, rounded distal margin and angular lateral projections (Fig. 7). Females are separable from other North American Peponapis by the almost completely dark pubescence, and the presence of a design of whitish hairs on the mesoscutum (Figs. 2, 6) (other species have mostly brown pubescence).
Description. Holotype male (Fig. 1). Body length, 13.00 mm; integument black except as noted below. Head: width, 4.11 mm; length, 3.05 mm; dorsal interocular distance, 2.21 mm; ventral interocular distance, 2.31 mm; eye length, 2.36 mm; eye maximum width, 1.23 mm; maxillary palpi with 4 segments, decreasing in length from first to last; mandible with 2 apical teeth, inner one wider, deep notch between teeth at 90° angle, internal (superior) margin with third, smaller denticle at three quarters of length of structure, subapical quarter reddish brown, brown spot subapically on external surface, inferior margin with white plumose hairs; labrum wider than long, with shallow notch on apical margin, with abundant decumbent white or yellow, apically directed pubescence, as long as pubescence on inferior margin of mandible, external surface dark brown with large yellow central mark (Fig. 5); clypeus with width almost twice length (1.16 mm: 2.30 mm), strongly produced in lateral view, with wide yellow spot subapically, apical margin light brown, granulate, with marked furrow, superficial punctures large, dense, but not contiguous, pubescence sparse; malar space as wide as clypeocular space; interantennal distance greater than antennocular (0.77 mm: 0.39 mm); scape swollen (0.44 mm), as long as interantennal distance (0.77 mm); F1 much shorter than F2 (0.20 mm: 0.69 mm); F2–F8 with rounded projections on inferior margin that give flagellum a crenulate aspect; facial pubescence yellow; ocellocular space with well defined depression; posterior interocellar distance larger than ocellocular space (0.68 mm: 0.59 mm); pubescence behind ocellus yellow, curved forward; vertex with narrow line of black pubescence interrupted medially, succeeded by abundant yellow pubescence. Mesosoma: pronotum with smooth, shining integument, except with dense, irregular punctures posterolaterally and on pronotal lobe; mesoscutum wider than long (3.59 mm: 2.95 mm), with integument granulate, slightly shining, with superficial punctures dense but separate; pubescence brown, short, with integument visible; scutellum with width slightly more than twice length (2.24 mm: 0.99 mm), punctures deep, closer than those on mesoscutum, pubescence short, brown, with some intermixed black hairs; metanotum with punctures similar to those on mesoscutum; tegula with dark brown integument, almost black; mesepisternum with granulate integument, punctures small, separated, pubescence yellow; propodeum with striate punctures on superior half of propodeal triangle, elsewhere smooth and shining, lateral surface with irregular granular integument, light brown, abundant pubescence. Wings: translucent with black venation; length of forewing, 10.04 mm, breadth, 3.46 mm. Legs: integument dark brown on apices of tibiae and tarsi except basitarsi; fore and mid legs with whitish pubescence on coxae, femora and tibiae; basitarsi with brown pubescence on anterior margins, posterior margin and external surface with pubescence dark brown, with light ends, these longer than hairs present on anterior margin; tarsi with brown pubescence; mid basitarsus almost double length of corresponding tibial spur (1.72 mm: 0.88 mm); hind femur with well defined tuft of dark reddish apically curved setose hairs (Fig. 4); hind tibia with whitish plumose pubescence except darker on extreme base, longer on posterior margin; hind basitarsus with dark brown plumose pubescence on posterior margin; external surface with sparse, plumose, brown pubescence; anterior margin with pubescence simple. Metasoma: terga with small, deep, uniform, punctures; T4 with smooth, shining integument, with apical hair band, width of band slightly less than minimum wide of F2 (0.22 mm: 0.29 mm), anterior surface of T1 with length and color of pubescence as on mesosoma, rest of the surface with short pubescence; T2 with basal pubescence laterally; T3–T5 with medially interrupted subapical bands of whitish pubescence, with some long black hairs; T6 with very narrow apical furrow; pubescence black; T7 with broadly rounded apical truncation, surface longitudinally striate, densely covered with dark pubescence, with black pubescence on lateral margin of pygidial plate; T2–T6 with lateral carinae, those on T5–T6 projecting posteriorly as small teeth; sterna black, with long, light pubescence curved ventrally; S6–S8 as in Figs. 7–9; genital capsule as in Fig. 10.
Female. Body length, 14.00 mm; integument black except as noted below; pubescence black and gray in macroscopic view (Figs. 2, 6). Head: length, 3.24 mm; width, 4.57 mm; dorsal interocular distance, 2.45 mm; ventral interocular distance, 2.70 mm; eye length, 2.61 mm; maximum eye width, 1.43 mm; mandible with 2 apical teeth little prominent, integument reddish medially, distally with orange brown spot extending almost to apex, inferior margin with whitish and dark pubescence; malar area very narrow; labrum with small, dense, superficial punctures; integument slightly shining, granulate, covered with abundant whitish, narrowly plumose hairs; clypeus with dense, superficial punctures, integument slightly shining, granular, apical margin with well defined furrow; face and frontal area with whitish pubescence; scape almost as long as F1 and F2 combined (0.96 mm: 1.02 mm), F1 twice length of F2 (0.68 mm: 0.34 mm); ocellocular distance shorter than distance between lateral ocelli (0.59 mm: 0.77 mm); pubescence of ocellar triangle whitish; vertex with black pubescence, becoming whitish posteriorly. Mesosoma: pronotum with whitish pubescence; mesoscutum wider than long (3.93 mm: 3.15 mm), punctures superficial, integument slightly shining, granular, pubescence black and whitish intermixed, posterolateral tuft and posterior margin with whitish hairs; scutellum with strongly granulated integument, pubescence of disc black, whitish on posterior margin; metanotum with integument more granular than on mesoscutum, with whitish pubescence; tegula dark brown, with pubescence black except white medially; mesepisternum with punctures as on mesoscutum, pubescence black. Wings: wings translucently darkened; length of forewing, 10.15 mm; width, 3.46 mm. Legs: pubescence black, except dark reddish brown on trochanters and bases of femora; fore tibia with long hairs, but shorter than length of fore tibia; mid tibial spur slightly longer than half of mid basitarsus (1.18 mm: 2.07 mm); hind tibia with the pubescence of anterior margin slightly shorter than on posterior margin, where slightly more plumose and with lighter tips; hind basitarsus rhomboid, pubescence on posterior margin long, plumose, on anterior margin shorter, simple. Propodeum with whitish pubescence, but grayish on sides, integument of posterior surface shining smooth. Metasoma: T1–T4 with well defined apical bands; T1 on anterior surface with whitish pubescence of similar length (some specimens with some dark hairs in middle), dorsal anterior area with band of dark pubescence, followed by narrow line of whitish pubescence mainly on carinate angle; T2 with anterolateral tuft of whitish pubescence; T3 and T4 with transverse, medially interrupted bands of primarily whitish pubescence with some long black hairs interspersed; T5 with white tuft posterolaterally, rest of surface with black pubescence, denser towards medial area; T6 with pygidial plate half oval in form, slightly truncated; S1 and S2 brown, S3–S7 with black pubescence.
Variation. In the males, the yellow spot on the clypeus (Fig. 3) varies in size and intensity, from intense in Sonora to almost absent in Chiapas. Females vary in the width of impunctate margin of T2.
Taxonomic summary
Type material. Holotype 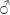 . Mexico. Jalisco, Chamela, 13–X–1985, S. H. Bullock #1942. Paratypes same locality as holotype, 1/8–X–1985, F. D. Parker and T. L. Griswold, 3, 4
. Mexico. Jalisco, Chamela, 13–X–1985, S. H. Bullock #1942. Paratypes same locality as holotype, 1/8–X–1985, F. D. Parker and T. L. Griswold, 3, 4 ; 11–X–1985, R. Ayala (RA 228), 1; 13–X–1985, S.H. Bullock #1942, 1; Melaque 27 km NW, 8–X–1993, L. Carrillo, A. Rodriguez and F. Noguera, 2? The holotype and 6 paratypes in the National Collection of Insects, Instituto de Biología, UNAM; 2 paratypes in USDA Bee Biology and Systematics Laboratory, Utah State University, Logan, UT; 2 paratypes in Essig Museum of Entomology, University of California, Berkeley, CA; and 1 paratype in the Biodiversity Institute, Entomology, University of Kansas, Lawrence, KS.
; 11–X–1985, R. Ayala (RA 228), 1; 13–X–1985, S.H. Bullock #1942, 1; Melaque 27 km NW, 8–X–1993, L. Carrillo, A. Rodriguez and F. Noguera, 2? The holotype and 6 paratypes in the National Collection of Insects, Instituto de Biología, UNAM; 2 paratypes in USDA Bee Biology and Systematics Laboratory, Utah State University, Logan, UT; 2 paratypes in Essig Museum of Entomology, University of California, Berkeley, CA; and 1 paratype in the Biodiversity Institute, Entomology, University of Kansas, Lawrence, KS.
Additional material. Mexico. Chiapas, El Sumidero, 14–IX–1974, G. Bohart and W. Hanson, ex "calabaza", 4. Jalisco, Tapalpa 7 km W, Mirador de Sayula, 6–X–2007, R. Ayala, ex Shizocarpum longisepalum, 1. Michoacán, Los Sabinos, 28 km S Ario de Rosales, 1190 m, 29–X–1987. T. Griswold, 1. Oaxaca, Totolapan 20 mi E., 17–IX–1973, W. J. Hanson and W. H. Haws, ex "squash", 3. 3.5 km NE Puerto Angel, 26–27–X–1989, A. Rodríguez and F. Noguera, 4. Sonora, Alamos, 27–VIII–1976, Hanson and Schwartz, ex "calabaza", 1. Specimens deposited in Estación de Biología Chamela, IBUNAM and USDA–ARS Bee Biology and Systematics Laboratory.
Etymology. The name "pacifica" makes reference to the known distribution of this species in Mexico throughout the coastal slope of the Pacific.
Distribution. Known throughout the coastal slope of the Pacific, between Sonora and Chiapas, and in the Balsas basin, in the state of Michoacán. It has been collected at elevations from sea level to around 1 000 m following the distribution of areas with tropical dry forest and the ecotone with cloud forest or pine and oak forest.
Remarks. Despite extensive collections on flowers of Cucurbita in Mexico, P. D. Hurd and collaborators failed to collect P. pacifica, possibly because they generally searched for bees early in the morning on the large orange flowers of Cucurbita, during the height of activity for the majority of the species of Peponapis. The specimens of P. pacifica from the Chamela biological station were collected on flowers of Schizocarpum longisepalum Jeffrey, a Cucurbitaceae (Lira–Saade, 1995, 1997; Lott, 2002) with yellow greenish flowers and greenish nectar guides that, unlike the orange flowers of Cucurbita, open in the morning and remain open throughout the day. Thus, P. pacifica is active diurnally and presumably primarily visits Schizocarpum, though it at least occasionally visits other Cucurbitaceae; some specimens were collected on flowers of domestic pumpkin. Schizocarpum is a genus with 11 species found in Mexico and Guatemala, apparently associated with the dry deciduous tropical forests of the west slopes, and is not present in the Yucatan Peninsula or the Gulf of Mexico slope (Lira–Saade, 1997). The association of P. pacifica with Schizocarpum suggests the possibility that additional undescribed species of Peponapis exist on flowers of other species of Cucurbitaceae.
At the Chamela field station, the type locality of P. pacifica, and surrounding areas, there are 3 other species of Peponapis, P. utahensis (Cockerell, 1905), P. azteca Hurd and Linsley, 1966 and P. crassidentata (Cockerell, 1949), all matinal and crepuscular visitors to flowers of Cucurbita aryrosperma sororia (L. H. Bailey) Merrick and Bates, as well as cultivated squashes (Cucurbita) and several species of Ipomoea (Lott, 2002; Ayala, 2004).
Peponapis parkeri Griswold and Ayala sp. nov. (Figs. 11–20)
Diagnosis: males can be distinguished from all other Peponapis by the dark pubescence of the labrum. In addition, the combination of narrow impunctate margin on T1 and very short F1 distinguish P. parkeri. The dark hairs of the labrum will separate females from the northern South American P. citrullina (Cockerell) and all other North American Peponapis, except P. atrata Hurd and Linsley. It can be distinguished from P. atrata by the light scopa and the presence of light tergal bands. Additional diagnostic characters include the combination of T1 with distinct, dense punctures on the disc (Fig. 15) and very narrow impunctate margin; complete light hair bands on T3–T5; largely dark haired mesoscutum; and maxillary palpus 5–segmented.
Description. Holotype male (Fig. 11), body length, 10.0 mm; integument black except as noted below. Head: width 3.45 mm; length, 2.64 mm; dorsal interocular distance, 2.18 mm; ventral interocular distance, 2.14 mm; length of ocular orbit, 1.77 mm, maximum width, 0.64 mm; maxillary palpi with 5 segments, decreasing in length from first to last; mandible with 2 apical teeth, internal not wider, shallow notch between teeth at less than 90° angle, internal (superior) margin with third smaller denticle at four fifths length from base, subapical quarter reddish brown, brown spot subapically on external surface, inferior margin with dark plumose hairs; labrum as wide as long, without notch on apical margin, with abundant decumbent dark apically directed pubescence as long as pubescence on inferior margin of mandible, external surface without yellow central mark; clypeus with width less than twice length (1.88 mm: 1.12 mm), strongly produced in lateral view, with small, irregular yellow spot subapically, apical margin dark brown, granulate, with indistinct furrow, superficial punctures large, dense, but not contiguous, pubescence sparse; malar space wider than clypeocular space; interantennal distance greater than antennocular (0.76 mm: 0.39 mm); scape swollen (0.38 mm), shorter than interantennal distance (0.76 mm); F1 much shorter than F2 (0.22 mm: 0.79 mm); flagellomeres without rounded projections on inferior margin, flagellum not appearing crenulate (Fig. 13); facial pubescence intermixed white and dark, with white predominant just above antennal socket; ocellocular space with well defined depression; posterior interocellar distance approximately equal to ocellocular distance (0.65 mm: 0.61 mm); pubescence behind ocellus largely dark; vertex with narrow line of black pubescence interrupted medially, succeeded by abundant white pubescence. Thorax: Pronotum with smooth, shining integument, except for dense, irregular punctures posterolaterally and on lateral lobe; mesoscutum wider than long (2.47 mm: 2.02 mm), with integument granulate, rather shining, with superficial punctures, moderately dense becoming sparse posteromedially; pubescence black except white along margins, long, integument visible; scutellum with width slightly more than twice length (1.78 mm: 0.77 mm), punctures shallow, much closer than on mesoscutum, pubescence short, black; metanotum with punctures similar to those on mesoscutum; tegula with reddish integument; mesepisternum with granulate integument, punctures small, separated, pubescence mixed light and dark; propodeum rugulose in propodeal triangle, lateral surface with irregular granular integument, pubescence long, abundant yellow. Wings: translucent with brown venation; length of forewing 8.58 mm, breadth 2.83 mm. Legs: integument dark brown on tarsi except basitarsi; legs with mixture of light and dark pubescence on tibiae and tarsi; mid basitarsus less than twice length of corresponding tibial spur (1.23 mm: 0.75 mm); hind femur with hairs denser towards base, without tuft of setose hairs (Fig. 14); Metasoma: terga with small, distinct, uniform, punctures; anterior surface of T1 with pubescence length and color as on mesosoma, rest of the surface with short pubescence; T2 with basal light pubescent band, partially covered by T1 and weak subapical band laterally; T3–T5 with uninterrupted subapical bands of whitish pubescence, with scattered long black hairs; T6 with pubescence dark brown; T7 reddish apically, with broadly rounded apical truncation, surface longitudinally striate, sparsely covered with dark pubescence, with black pubescence on lateral margin of pygidial plate; T2–T6 with lateral carinae, not projecting posteriorly as small tooth; sterna with long mixed light and dark, straight pubescence; S6–S8 as in Figs. 17–19; genital capsule as in Fig. 20.
Female. Body length, 11.00 mm; integument black except as noted below; pubescence dark and reddish in macroscopic view (Figs. 12, 15, 16). Head: length, 2.73 mm; width, 4.00 mm; dorsal interocular distance, 2.41 mm; ventral interocular distance, 2.47 mm; eye length, 1.92 mm; maximum eye width, 0.78 mm; mandible with 1 apical tooth, integument reddish medially, distally with orange brown spot extending almost to apex, inferior margin with dark pubescence; malar area narrow; labrum with small, dense, superficial punctures; integument shining, covered with abundant dark, narrowly plumose hairs; clypeus with dense, superficial punctures, integument slightly shining, granular, apical margin with well defined furrow; face with mostly dark pubescence except horizontal supra–antennal band and lower paraocular area; scape shorter than F1 and F2 combined (0.56 mm: 0.76 mm), F1 almost twice length of F2 (0.66 mm: 0.25 mm); ocellocular distance shorter than distance between lateral ocelli (0.66 mm: 0.69 mm); pubescence of ocellar triangle dark; vertex with black pubescence, becoming whitish posterolaterally. Mesosoma: pronotum with dark pubescence; mesoscutum wider than long (2.91 mm: 2.14 mm), punctures superficial, integument shining, slightly granular, pubescence black on disk, yellow anteriorly and posteriorly, yellow along lateral margin (Fig. 16); scutellum with strongly granulated integument, pubescence of disc black, whitish on lateral margin; metanotum with integument as granulate as scutellum, pubescence whitish; tegula reddish, pubescence light; mesepisternum with punctures much finer, sparer than on mesoscutum, pubescence mixed dark and light (Fig. 16). Wings: wings translucently darkened; length of forewing, 8.46 mm; width, 2.73 mm. Legs: pubescence black, except light on apices of fore and mid tibiae, hind femur and hind tibia almost entirely; fore tibia with long hairs, but shorter than length of fore tibia; mid tibial spur slightly longer than half of mid basitarsus (0.94 mm: 1.26 mm); hind tibia with the pubescence of anterior margin slightly shorter than on posterior margin, where more plumose; hind basitarsus rhomboid, pubescence on posterior margin long, plumose, on anterior margin simple, shorter. Propodeum with yellowish pubescence, long laterally. Metasoma: T1 with pubescence of anterior surface long, erect, yellowish, dorsal area with band of dark pubescence, with whitish on lateral edge; T2 with anterolateral tuft of whitish pubescence mostly covered by T1, with weak lateral subapical white band; T3–T5 with well defined uninterrupted subapical bands; T5 with dark apical fimbria; T6 with pygidial plate half oval in form; S1 and S2 brown, S3 and S4 with light margins; S3–S7 with brown, erect pubescence.

Variation. In the males, the yellow spot on clypeus varies in size and intensity, and is absent in one individual.
Taxonomic summary
Type material. Holotype . Costa Rica, Alajuela, Bijagua, 20 km S Upala, 5–18–III–1991, F. D. Parker. Paratypes: same data as holotype, 3, 1; same locality and collector, 20–I–12–II–1991, 2; 19–II–1991, 2; 12–II–5–III–1991, 4; Guanacaste, Finca Montezuma, 3 km SE Río Naranjo, XII–1991, 2; 10/23–I–1992, 1; 19/24–II–1992, 1; 29–I–8–II–1993, 1; 18/21–II–1993, 1; 12/20–III–1993, 1. Holotype deposited in the U. S. National Pollinating Insects Collection, USDA Bee Biology and Systematics Laboratory, Utah State University, Logan, UT; paratypes in the U. S. National Pollinating Insects Collection and INBio.
Etymology. It is a great pleasure to name this bee after Frank Parker, who has been a mentor and friend to both authors.
Distribution. Known throughout the Pacific slope of Costa Rica.
Remarks. This species is most similar to the South American P. citrullina, which ranges as far north as Colombia (Hurd and Linsley, 1967).
Acknowledgments
We especially thank Frank Parker for his aid and support of this work, Charles D. Michener for valuable comments, and Harold Ikerd for the illustrations of Peponapis parkeri.
Literature cited
Ayala, R. 1989. La fauna de abejas silvestres (Hymenoptera; Apoidea) de Chamela, Jalisco. México. Folia Entomológica Mexicana 77:395–493. [ Links ]
Ayala, R. 2004. Fauna de abejas silvestres (Hymenoptera: Apoidea). In Artrópodos de Chamela, A. N. García–Aldrete and R. Ayala (eds.). Instituto de Biología, Universidad Nacional Autónoma de México (UNAM), México, D.F. p. 193–219. [ Links ]
Ayala, R., T. Griswold and D. Yanega. 1996. Apoidea. In Biodiversidad, taxonomía y biogeografía de artrópodos de México, J. E. Llorente, A. N. García Aldrete and E. González (eds.). Universidad Nacional Autónoma de México (UNAM), México, D.F. p. 423–464. [ Links ]
Giannini, T. C, R. Lira–Saade, R. Ayala, A. M. Saraiva and I. Alves–dos–Santo. 2011. Ecological niche similarities of Peponapis bees and non–domesticated Cucurbita species. Ecological Modelling 222:2011–2018 [ Links ]
Hurd, P. D. and E. G. Linsley. 1964. The squash and gourd bee–genera Peponapis Robertson and Xenoglossa Smith inhabiting America north of Mexico (Hymenoptera; Apidae). Hilgardia 35:375–477. [ Links ]
Hurd, P. D. and E. G. Linsley. 1966. The Mexican squash and gourd bees of the genus Peponapis (Hymenoptera: Apoidea). Annals of the Entomological Society of America 59:835–851. [ Links ]
Hurd, P. D. and E. G. Linsley. 1967. South American squash and gourd bees of the genus Peponapis (Hymenoptera: Apoidea). Annals of the Entomological Society of America 60:647–661. [ Links ]
Hurd, P. D. and E. G. Linsley. 1970. The classification of the squash and gourd bees Peponapis and Xenoglossa (Hymenoptera; Apidae). University of California Publications in Entomology 62:1–39. [ Links ]
Klein, A. M., B. E. Vaissiere, J. H. Cane, I. Steffandewenter, S. A. Cunningham, C. Kremen and T. Tscharntke. 2007. Importance of pollinators in changing landscapes for world crops. Royal Society 274:303–313. [ Links ]
Lira–Saade, R. 1995. Estudios taxonómicos y ecogeográficos de las Cucurbitaceae latinoamericanas de importancia económica. Systematic and Ecogeographic Studies on Crop Genepools 9:1–115. [ Links ]
Lira–Saade, R. 1997. Nuevos registros de Cucurbitaceae para varias regiones de México. Acta Botanica Mexicana 41:17–20. [ Links ]
Lott, E. 2002. Listado anotado de las plantas vasculares de Chamela–Cuixmala. In Historia natural de Chamela, F. A. Noguera, J. H. Vega–Rivera, A. N. García– Aldrete and M. Quesada–Avendaño (eds.). Universidad Nacional Autónoma de México, México, D.F. p. 99–142. [ Links ]
Mayer C., L. Adler, W. S. Armbruster, A. Dafni, C. Eardley, S.–Q. Huang, P. G. Kevan, J. Ollerton, L. Packer, A, Ssymank, J. C. Scout and S. G. Potts. 2011. Pollination ecology in the 21st Century: Key questions for future research. Journal of Pollination Ecology 3:8–28. [ Links ]
Merrick, L. C. and D. M. Bates. 1989. Classification and nomenclature of Cucurbita argyrosperma. Baileya 23:98. [ Links ]
Michener, C. D. 2007. The bees of the world. Second edition. The Johns Hopkins University Press, Baltimore and London. 953 p. [ Links ]
Moure, J. S., D. Urban and G. A. R. Melo. 2007. Catalogue of bees (Hymenoptera, Apoidea) in the Neotropical region, V.1. Curitiba, Sociedade Brasileira de Entomologia 1072–1058 p. [ Links ]

