











 text new page (beta)
text new page (beta) English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO 
 Permalink
Permalink1. Introduction
Fossil marine mammals, especially whales and dolphins (Cetacea), have been reported widely from around the Pacific margin. Many species have been named from localities in western North America from Alaska to California, Peru, New Zealand, eastern Australia, and Japan. In Mexico Oligocene marine mammals from Baja California Sur have been reported since the 1940s, and most of the identified Oligocene fossils are cetacean, but desmostylids are known and some fragmentary bones could belong to sirenians and pinnipeds. These occurrences are important in providing insights into the poorly understood marine mammals of Oligocene age from warm temperate to subtropical settings.
Amongst the early records, Vanderhoof (1942) and Durham (1950) reported remains of the desmostylian Cornwallius sookensis (Cornwall, 1922), and other fossil bones from Punta San Telmo (Tembabichi Member, locality “miembro Punta San Carlos” in Bahía San Carlos). Other marine mammal fossils were reported in the 1970s, from Punta San Telmo, San Hilario, El Cien, and San Juan de la Costa (Applegate and Wilson, 1976). One skull of a baleen whale from the El Cien locality was considered similar to the Oligocene-Miocene mysticete Mauicetus parki (Benham, 1937) from New Zealand (Downs and Morris, 1974).
Reports from the 1980s include fragments of odontocete mandibles from the Aguajito area and squalodontid teeth are known from the phosphatic sandstones at the locality “Diez minutos” (near El Cien village). Mysticete skulls are known from localities north of La Fortuna and in the vicinity of Tembabichi (Applegate, 1986). The whereabouts of specimens collected or reported during the 1940s to early 1990s are unknown but some may be in the Universidad Autónoma de México (UNAM) collections (see Barnes, 1998).
Around 16 species of cetaceans were reported during the 1990s from outcrops of San Juan Member, El Cien Formation (in the village of San Juan de la Costa, El Cien and San Hilario), based on field observations and uncatalogued specimens from the collections of the Museo de Historia Natural de la Universidad Autónoma de Baja California Sur (MHN-UABCS) and material from the Museo Regional de Antropología e Historia de Baja California Sur (Barnes, 1998). Other unpublished specimens from the San Juan Member include a skull of a putative agorophiid odontocete (Cruz-Marín, 1997) and a skull of an archaic toothed mysticete (Olivares-Bañuelos, 2001).
The Oligocene cetacean fossil record from Baja California Sur is restricted to the San Gregorio Formation and El Cien Formation (Figure 1). Most of the specimens are from the San Juan Member (El Cien Formation), from localities near the village of San Juan de la Costa. However, it is known that the distribution of fossils is broader, with abundant material in places like El Aguajito, El Cien, Cerro Colorado, Tembabichi (Tembabichi Member, El Cien Formation) and La Purísima (San Gregorio Formation).
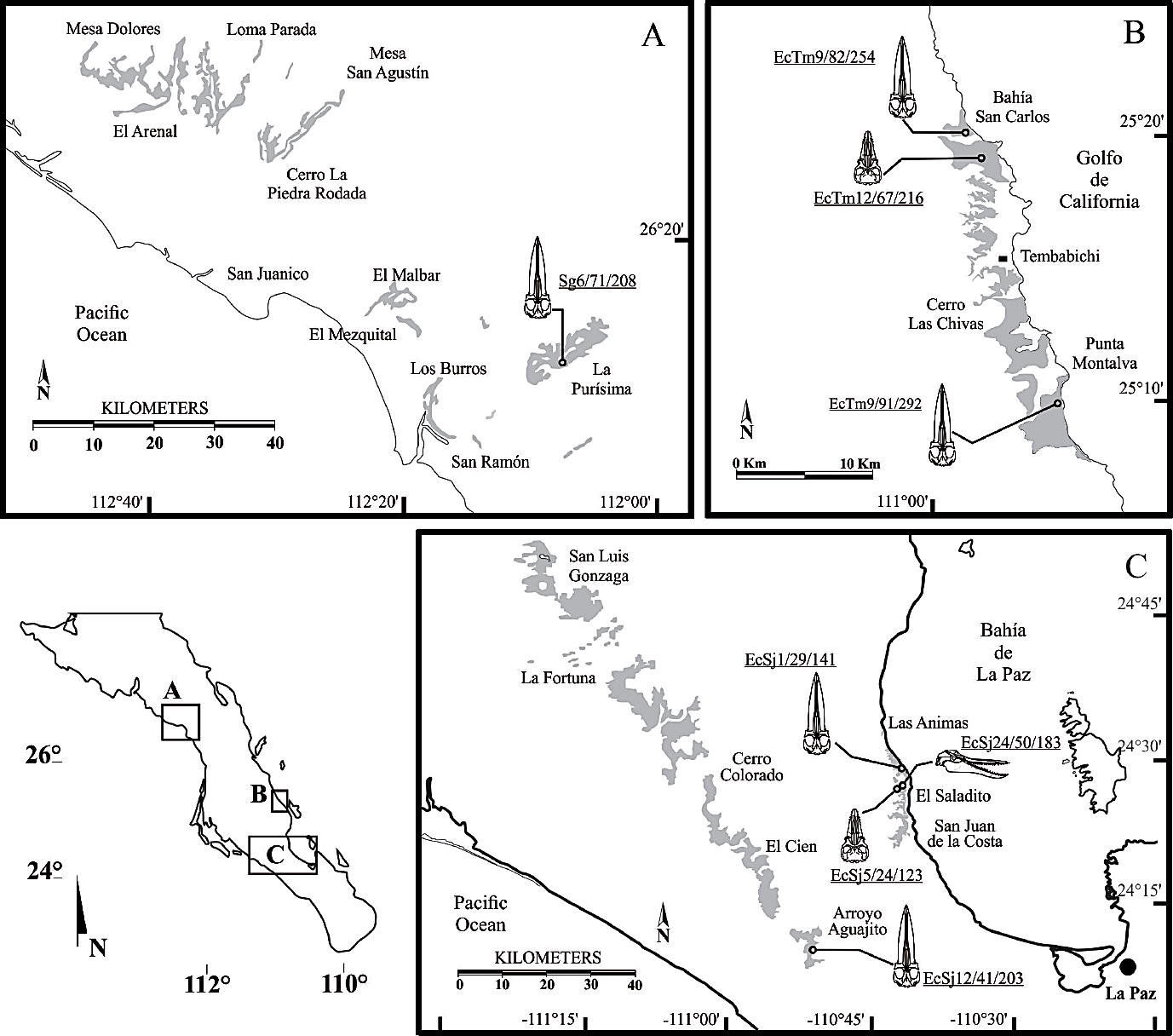
Figure 1 Geologic map of Oligocene outcrops in Baja California Sur, and some representative specimens collected from the localities. San Gregorio Formation (A), Tembabichi Member of the El Cien Formation, (B), and San Juan Member of the El Cien Formation (C), Geological distribution extracted from the map of the Servicio Geológico Mexicano (Alexadri-Rionda et al., 2008).
Since the 2000s more cetacean fossils have been collected from the Oligocene outcrops in Baja California Sur, leading to recent published mentions and some provisional identification (González-Barba, 2007; Hernández-Cisneros and González-Barba, 2013). In summary, the fossil record is diverse at species to family level, but identifications are provisional because most of the specimens need detailed preparation, and the specimens are largely unpublished. Nevertheless, the Oligocene cetaceans from Baja California Sur are potentially important to understand the initial evolutionary radiation of the Neoceti - the two extant clades of Mysticeti (filter-feeding baleen whales) and Odontoceti (echolocating dolphins and toothed whales). The aim here is to review Oligocene cetacean assemblages from Baja California Sur, and add details of specimens that have previously been mentioned only in unpublished theses or abstracts. The geographic distribution and stratigraphy of the main source horizons are summarized. Fossils of Oligocene cetaceans recently cataloged in MHN-UABCS collections improve knowledge of Oligocene cetacean diversity in Baja California Sur (Hernández-Cisneros, 2012).
2. Study area and geological setting
Fossiliferous Oligocene rocks in Baja California Sur include: the San Gregorio Formation, located around La Purísima ~250 km north of La Paz near the Pacific coast (Fischer et al., 1995); the Tembabichi Member, El Cien Formation, on the coast of the Gulf of California, approximately 50 km north of the Bahía de La Paz (Fischer et al., 1995; Plata-Hernández, 2002); and the San Juan Member, El Cien Formation, which lies between the bay in the East and the State Road 1 in the west (Fischer et al., 1995) ~40 km north of La Paz (Figure 1).
The localities of the San Gregorio Formation are small outcrops along the banks and stream bottoms of the largest arroyos (McLean et al., 1987), unlike the more extensive distribution of the Tembabichi and San Juan members. Oligocene outcrops are discontinuous and reflect different depositional environments with relatively similar stratigraphy that include phosphatic sandstone beds, mudstone, diatomite, coquina and conglomerates (Hausback, 1984; Fischer et al., 1995).
The San Gregorio Formation is characterized by dark finely to very finely laminated biosiliceous sediments -white diatomite, diatomaceous clay, porcelanite, silty porcelanite and chert (Grimm and Föllmi, 1994), with several alternating sequences of phosphatic layers, siliceous siltstone, phosphatic mudstone with fish scales, and rhyolitic tuffs (McLean et al., 1987). The Tembabichi Member consists of siltstones, sandstones, mudstones, coquina, phosphatic horizons, and dolomitic beds and it is considered correlative to the San Juan Member as a lateral variation of the same basin deposit (Plata-Hernandez and Schwennicke, 2000; Plata-Hernandez, 2002). The San Juan Member is mainly composed of laminated, partly tuffaceous, siliceous, diatomaceous or phosphatic mudstones, partly phosphatic silt- and sandstones, granular phosphorite, tuff, and conglomerates (Fischer et al., 1995).
The depositional environment for the San Gregorio Formation is interpreted as deposits from water depths between 500 and 2500 m based on planktic foraminifers and trace fossils (McLean et al., 1987; Grimm and Föllmi, 1994). In contrast, sediments of El Cien Formation were deposited no deeper than 200 m, suggesting shelf environments, coastal lagoons, estuaries and deltas (Schwennicke, 1994). Past studies of diatoms (Kim and Barron, 1986), planktonic foraminifera (Kim, 1987), and calcareous nannofossils (McLean et al., 1987; Schwennicke, 1992), shark and ray faunas (González-Barba, 2008), macroinvertebrates -e. g. mollusks and crabs (Smith, 1991; Grimm and Föllmi, 1994; Schweitzer et al., 2002; Vera-Dimas, 2013), phosphorites (Galli-Olivier et al., 1990; Galli-Olivier, 1993; Schwennicke, 1994; Schwennicke and Vázquez-García, 1996), and sedimentological studies (Grimm, 1992) have elucidated the paleobiological and paleoceanographic history of the region during the Late Oligocene and indicate a subtropical environment characterized by high primary productivity.
The age for the Oligocene sequences range ~30 to 23 Ma. The thickness for the main exposed section of San Gregorio Formation is ~75 m (Ganz, 1996), and has been dated as ~27 to 22 Ma (Late Oligocene-Early Miocene) based on K/Ar, calcareous nannofossils (Discoaster ) and diatoms -Rocella gelida (Bukry, 1978) (Kim and Barron, 1986; McLean et al., 1987). For the Tembabichi Member, the type section is ~135 m thick (Grimm, 1992), with an age of ~28 to 25. 9 Ma dated with K/Ar and Rb/Sr using tuff beds (Hausback, 1984; Plata-Hernández, 2002). Thickness of the San Juan Member varies from 75 m at Aguajito locality to 130 m at San Juan de la Costa locality (Schwennicke and Vázquez-García, 1996). Its age is ~30 to 23 Ma based on K/Ar dates for biotite from a tuff bed (Hausback, 1984), diatom biostratigraphy -Rocella vigilans Fenner, 1982, Bogorovia veniamini, Jousé, 1973, R. gelia (Kim and Barron, 1986), foraminifera representing zones P21 and P22 (Kim, 1987) and calcareous nannofossils representing the NP24/NP25 zones (Schwennicke, 1992).
3. Methods and Materials
This study involved field exploration, preparation, identification, and catalog of Oligocene cetacean fossils from MHN-UABCS collection. We reviewed the available literature, outcrops and fossils of the San Gregorio and El Cien formations. Field collections of fossil material for MHN-UABCS began in 1988, and were recorded by Gerardo González Barba, from 1991 to present. New observations (Table 1) are derived from Hernández-Cisneros (2012, 2014, 2015), Fordyce et al., (2012), Hernández-Cisneros and González-Barba (2013), and Hernández-Cisneros et al., (2014). References for taxa mostly cite the original author, using the author-comma-date convention (with parentheses as appropriate) of the International Code of Zoological Nomenclature.
Table 1 Representatives list of Late Oligocene cetaceans from Baja California Sur.

* Specimens in process of description with the collaboration of Robert Ewan Fordyce from the University of Otago, Dunedin, New Zealand.
Institutional abbreviations. MHN-UABCS (MU), Museo de Historia Natural de la Universidad Autónoma de Baja California Sur, La Paz, Baja California Sur, Mexico; OU, specimen curated in Geology Museum, University of Otago, Dunedin, New Zealand; USNM, specimen curated in the National Museum of Natural History, Smithsonian Institution, Washington D. C.
4. Results
Cetacean fossils are sporadic to common in Late Oligocene outcrops of El Cien Formation and San Gregorio Formation. Generally the fossils occur within phosphatic sandstone near the top of the sequence (Appendix Figure A1). In contrast, the Tembabichi area has abundant fossils which are associated with siltstones and phosphatic sandstones, and appear in the section close to the top, in the middle part (mostly allochthonous in phosphatic beds), and close to the base -above ~30-35 m in the stratigraphic column (Appendix Figure A2). The fossils are in concretions or are scattered fragments on the surface. In the San Juan Member, the fossil distribution is similar to Tembabichi Member. The top of the San Juan Member is capped by a conglomerate with sporadic marine mammal fossils in some localities to the north of El Saladito. In the middle part of the exposed stratigraphic section in San Juan de la Costa complete skeletons occur mainly in the zones where the phosphorite mine work is active. More cetacean bones can be found close to the base of the San Juan Member, ~20 m above the lower limit at Aguajito locality (Appendix Figure A3).
Fossil cetaceans from Baja California Sur are noteworthy in terms of number of species and individuals, which represent two major taxonomic groups: the paraphyletic Archaeoceti or stem-Cetacea, and the monophyletic Neoceti (Mysticeti and Odontoceti) or crown-Cetacea. Specimens generally show enough distinctive features to be separated at species level, thus allowing minimum counts for species, but few are preserved or prepared well enough to justify naming at this stage of research.
Some Mysticeti specimens, as MHN-UABCS_EcSj5/24/123 (Figure 2A) exhibit synapomorphies of the family Aetiocetidae, including a premaxilla that overhangs the maxilla adjacent to the bony naris (Fitzgerald, 2010) and a short overlap of the jugal and the zygomatic process of the squamosal (Deméré and Berta, 2008) - from here MU will be used for MHN-UABCS in the series-collection to give an easy read (see the complete specimen references in Table 1).

Figure 2 Late Oligocene cetacean fossils from Baja California Sur, México. (A) Dorsal view, skull of an unnamed aetiocetid, MU EcSj5/24/123. (B) Left, dorsal view, partial skull, MU EcSj9/08/36, ?Chonecetus sp. , and right, concretion with the partial skull in ventral view. Continuous lines show anatomical details, and dashed lines mark missing anatomy. Shaded white areas show sediment matrix. (C) Dorsal view, juvenile skull of an unnamed eomysticetid, MU EcSj1/29/141, and reconstruction scheme, right.
Well preserved toothless mysticetes (Figure 2C, 3) share similar cranial features with eomysticetids like Eomysticetus and Micromysticetus (Sanders and Barnes, 2002a, 2002b), Yamatocetus (Okazaki, 1995, 2012) and Tohoraata, Waharoa and Tokarahia (Boessenecker and Fordyce, 2014, 2015a, 2015b). In general these specimens have a long, narrow skull, including an elongate and narrow intertemporal region, elongate nasals, a large temporal fossa, an elongate zygomatic process of the squamosal that extends beyond the apex of the supraoccipital, and a long-based and high coronoid process. The family identities of other fossil mysticetes are unclear.

Figure 3 Dorsal view, skull of an unnamed eomysticetid, MU EcSj1/29/142; reconstruction scheme, right, red lines show the premaxillae as the distinctive feature. Continuous lines show anatomical details, dashed lines mark missing anatomy, and diagonal lines mark broken areas.
Few Oligocene Odontoceti skulls have been identified, but it is likely that the number of specimens will increase with more field work, given the success of recent prospecting. Fordyce et al. (2012) presented preliminary details of a new group of archaic odontocetes that are structurally similar to New Zealand Oligocene “dalpiazinid” dolphins, but more derived in some cranial features. These specimens, which are undescribed, include MU EcSj24/50/183 (Figure 5B). The latter Baja California Sur specimen is phylogenetically close to USNM205491 from the Alsea Formation of Oregon, ≤ 30 Ma (Snavely et al., 1975).

Figure 4 (A-B) Dorsal view, partial skulls, unnamed mysticetes, MU EcSj5/06/31, left; and MU EcSj5/18/95, right. (C) Left periotic bone in situ of MU EcSj5/06/31, ventral view. (D-D’) broken left periotic of MU EcSj5/18/95, medial view, left, and lateral view, right.

Figure 5 (A) Dorsal view, skull of unnamed archaic odontocete, MU EcSj5/12/48. (B) Dorsal view, skull of an unnamed archaic odontocete, MU EcSj24/50/183 (Fordyceet al . , 2012). Continuous lines show anatomical details and diagonal lines mark broken areas. Shaded white areas show sediment matrix.
Probable kekenodontids are represented by: a periotic, MU EcSj5/16/267 (Figure 6A); a left frontal fragment, MU EcSj5/16/72 (Figure 6B); and unprepared skulls, MU EcSj1/45/214, EcSj1/94/296, and EcSj1/38/155 (Hernández-Cisneros, 2015). These specimens are similar to an undescribed specimen (OU 22294) from the Kokoamu Greensand-basal Otekaike Limestone of the Waitaki Valley region (26-28+ Ma, Late Oligocene; Fordyce, 2004) from New Zealand.

Figure 6 (A) Medial view, left periotic, ?kekenodontid-like animal, MU EcSj5/16/267. (B) Dorsal view of a frontal fragment, ?kekenodontid-like animal, specimen MU EcSj5/16/72, dashed lines mark missing anatomy.
4. 1. Systematic Paleontology
4. 1. 1. Suborder Mysticeti-Aetiocetidae
Cetacea Brisson, 1762
Neoceti Fordyce & de Muizon, 2001
ysticeti Gray, 1864
Aetiocetidae Emlong, 1966
?Aetiocetus Emlong, 1966
?Aetiocetus sp.
Material. MHN-UABCS_EcSj5/24/123, skull (Figure 2A).
Locality. El Saladito, “Mesa El Saladito”, San Juan de la Costa (24. 43929 N, 110. 70042 W), La Paz, Baja California Sur, México. Collected by Gerardo González Barba and Raúl Agundez Arce. March 1996.
Formation. El Cien Formation.
Age -28 to 25 Ma approximately, Late Oligocene (Fischer et al., 1995).
Description. The specimen (Figure 2A) consists of a skull with a condylobasal length of ~760 mm, a bizygomatic width of ~320 mm and an estimated body length of ~3. 03 m derived from the bizygomatic width (following Pyenson and Sponberg, 2011). The inferred age class is a subadult individual, as indicated by the nature of the cranial sutures which are mainly closed but not fused or obliterated. The occipital condyles, ear bones, and postcranial elements generally lack fine details. Features of note include: elongate and narrow nasal bones 5-6 % longer than in Aetiocetus weltoni (see Barnes et al., 1995), a wide and short mesorostral groove, thick and elongate zygomatic process of squamosal without the thin dorsoventral shape in its middle portion, and a wide temporal fossa. These features contrast notably with Fucaia goedertorum (Barnes et al., 1995), A. weltoni, and Aetiocetus polydentatus both of Barnes et al., 1995, and Aetiocetus cotylalveus Emlong, 1966. The features differ in part from those of Aetiocetus tomitai Barnes et al., 1995, Morawanocetus yabukii Barnes et al., 1995, Chonecetus sookensis Russell, 1968 (see also Barnes et al., 1995), Fucaia buelli Marx et al., 2105 and Willungacetus aldingensis Pledge, 2005 (see also Fitzgerald, 2010). Other notable features include multiple dorsal infraorbital foramina, thin and elongate lacrimal with lobed end, intertemporal region without sagittal crest, a slightly convex supraoccipital shield with a semicircular-triangular outline and a weak external occipital crest on middle line. The specimen is placed in Aetiocetus because it has some synapomorphies for the group (Fitzgerald, 2010), such as an incompletely open mesorostral groove partially roofed by the premaxilla, and the anterior edge of the nasals is forward of the anterior edge of the supraorbital process of frontal.
?Chonecetus Russell, 1968
?Chonecetus sp.
Material. MHN-UABCS_EcSj9/08/36, partial skull, tympanic bullae, few teeth and jaw fragments. (Figure 2B).
Locality. Mina “Las Crucecitas”, San Juan de la Costa (24. 40859 N, 110. 69608 W), La Paz, Baja California Sur, México. Collected by Gerardo González Barba and Ruben Castro Arvizá. August 1992.
Formation. El Cien Formation.
Age. 28 to 25 Ma approximately, Late Oligocene (Fischer et al., 1995).
Description. The specimen (Figure 2B) consists of a partial skull with both tympanic bullae, and jaw fragments with few teeth; its preserved length is 400 mm, without occipital condyles, bizygomatic width is estimated at ~300 mm, and the calculated body length is ~2. 86 m (following Pyenson and Sponberg, 2011). The age class is adult, as indicated by generally closed cranial sutures. Features of note are: a fine suture in the middle line of the frontal; a convex, soft and tenuous sagittal crest on the intertemporal region; a broad intertemporal region (~59 mm wide) with a U upside down shape, and a total width between postorbital processes of ~240 mm. These features contrast with the type species of Chonecetus, C. sookensisRussell, 1968 and F. goedertorum (Barnes et al., 1995). Nevertheless, the specimen is placed in Chonecetus because it shows some synapomorphies that Fitzgerald (2010) recognized for that genus: dorsal braincase edge low and flat, with a profile less of 10° to the lateral edge of the rostrum; possibly a V-shape frontoparietal suture, and a low nuchal crest of the supraoccipital that apparently does not project laterally or dorsolaterally. Conversely, MU EcSj9/08/36, contrasts with the recently named Fucaia buelli (see Marx et al., 2015) in the anteroposterior extension of supraoccipital and thickness of the sagittal crest.
4. 1. 2. Suborder Mysticeti-Eomysticetidae
Chaeomysticeti Mitchell, 1989
Eomysticetidae Sanders & Barnes, 2002b
Eomysticetidae gen. et. sp. indet. 1
Material. MHN-UABCS_EcSj1/29/141, skull, four thoracic vertebrae, one caudal vertebra, ear bone fragments, and bone fragments (Figure 2C).
Locality. Mesa Tesoro, San Juan de la Costa (24. 47366 N, 110. 69646 W), La Paz, Baja California Sur, México. Collected by Gerardo González Barba; Lawrence G. Barnes; James L. Goedert, and two anonymous collectors (Luisa and Memo), November 1999.
Formation. El Cien Formation.
Age. 28 to 25 Ma approximately, Late Oligocene (Fischer et al., 1995).
Description. The specimen (Figure 2C) consists of a partial skull of a juvenile mysticete, and associated elements. The age class is indicated by cranial sutures which are moderately but not fully closed, e. g. the junction of the squamosal and parietal is still quite apparent. Estimated condylobasal length is ~620 mm; bizygomatic width is ~320 mm, and body length is ~3. 03 m (following Pyenson and Sponberg, 2011). Features include: elongate, triangular, nasal bones; a wide frontal similar to Yamatocetus canaliculatusOkazaki, 2012 (see also Okazaki, 1995); a short and relatively thick intertemporal region with a fine sagittal crest, a supraoccipital shield with a semicircular outline similar to mammalodontids and archaeocetes; an external occipital crest, vertical nuchal crests; and a squamosal prominence like a fingertip on the posterolateral part of squamosal (Hernández-Cisneros et al., 2014). The specimen is placed within Eomysticetidae because it shows: rigid, firmly sutured nasals and premaxillae; posteriorly elevated nuchal crest as the highest point of the skull; anteroposteriorly oriented zygomatic process; and longitudinally twisted zygomatic process that lacks a supramastoid crest (features discussed by Boessenecker and Fordyce, 2015a, 2015b).
Eomysticetidae gen. et. sp. indet. 2
Material. MHN-UABCS_EcSj1/29/142, partial skull, four cervical vertebrae, eight thoracic vertebrae, four lumbar vertebrae, and fragmentary bones (Figure 3).
Locality. Mesa Tesoro, San Juan de la Costa (24. 47366° N, 110. 69646° W), La Paz, Baja California Sur, México. Collected by Gerardo González Barba; Lawrence G. Barnes; James L. Goedert, and two anonymous collectors (Luisa and Memo), November 1999.
Formation. El Cien Formation.
Age. 28 to 25 Ma approximately, Late Oligocene (Fischer et al., 1995).
Description. The specimen (Figure 3) includes of a partial skull, lacking the condyles and rostrum apex, with a preserved total length of 1100 mm. Assuming proportions similar to those in Eomysticetus whitmoreiSanders and Barnes, 2002 the complete condylobasal length could have reached ~1600 mm. The vertebrae are unprepared. The age class is subadult as indicated by cranial sutures which are well closed, but distinct, and not completely fused. Features of note include: very narrow elongate nasal bones; a wide mesorostral groove; a narial process of the frontals; and an extended ascending process of the premaxilla which forms a broad posterior apex that covers part of the frontal. Posteriorly, behind the level of the orbit, a serrate suture runs obliquely anterolaterally toward the region of the lacrimal and/or antorbital notch. This suture forms the posterior margin of the ascending process of the maxilla with an uncertain contribution from the premaxilla. This specimen differs in the latter structure of the premaxilla from Yamatocetus, Eomysticetus, the New Zealand eomysticetids, and other known eomysticetids (Hernández-Cisneros, unpublished data). The specimen is placed in Eomysticetidae because the cranial morphology is phenetically similar to other eomysticetids in features including: a narrow and long rostrum, firmly or rigidly sutured nasals and premaxillae; supraorbital process of frontal medially elongated and transversely narrow; intertemporal region exposed (parietal + frontal); and a large temporal fossa (characters reviewed by Boessenecker and Fordyce, 2015a, 2015b).
4. 1. 3. Suborder Mysticeti-Chaeomysticeti
Family indet. gen. et. sp. indet.
Material. MHN-UABCS_EcSj5/06/31, partial skull with left periotic; and MHN-UABCS_EcSj5/18/95, partial skull with a left bulla and periotic (Figure 4).
Locality. San Juan de la Costa, El Saladito, mine spoils (24. 43916 N, 110. 69512 W). La Paz, Baja California Sur, México. Collected by Gerardo González Barba, December 1991 and May 1994.
Formation. El Cien Formation.
Age -28 to 25 Ma approximately, Late Oligocene (Fischer et al., 1995).
Description. The specimens (Figure 4) consist of two partial skulls which differ from each other in dorsal cranial morphology, but share a similar periotic bone. The condylobasal length is lost for both, but other size estimates are possible. MU EcSj5/06/31 has an estimated bizygomatic width of ~750 mm based on the dimension from the midline at the condyle to the margin of the zygomatic process. The condyle width is ~126 mm, and its calculated size body is 6. 65 to 7. 70 m using the equations for stem Mysticeti and stem Balaenopteroidea in Pyenson and Sponberg (2011). MU EcSj5/18/95 is fragmentary, comparable in size with MU EcSj5/06/31, but with no meaningful standard dimensions preserved. Both specimens are subadults as indicated by cranial sutures that are not completely fused. Features of note include: a periotic bone with very flat (transversely compressed) anterior process; and compound posterior process with a marked facial nerve sulcus, similar to the narrower and more elongated process of balaenopterids (Ekdale et al., 2011). Notable primitive characters of the periotic include: a superior process; and an internal acoustic meatus with a single large aperture without a prominenttransverse crest (Hernández-Cisneros et al., 2014). The fossils are too incomplete to assess possible synapomorphies for Chaeomysticeti, such as absence of teeth (Mitchell, 1989) or an orbital crest well positioned anteriorly on the dorsal surface of the frontal (Geisler and Sanders, 2003). But, other features indicate that the fossils are a species of chaeomysticetes: compound posterior process of the periotic; transversally widened intertemporal constriction; triangular shape of the supraoccipital, and a postglenoid process of the squamosal posteriorly directed in lateral view (see Marx and Fordyce, 2015, on these features). The specimens MU EcSj5/06/31 and MU EcSj5/18/95 are quite different from Horopeta umarere Tsaiand Fordyce, 2015, Whakakai waipata Tsai and Fordyce, 2016, and M. parki Benham,1937 (see also Fordyce, 2005) mainly in the ear bone morphology and other cranial features (Hernández-Cisneros unpublished data).
4. 1. 4. Suborder Odontoceti-Archaic Odontocetes
Odontoceti Flower, 1867
Family indet. gen. et. sp. indet.
Material. MHN-UABCS_EcSj5/12/48, skull, right forelimb (humerus, radio, ulna), periotics, right bulla, cervical vertebrae, and teeth (Figure 5A).
Locality. San Juan de la Costa, El Saladito, mine spoils (24. 44249 N, 110. 69106 W), La Paz, Baja California Sur, México. Collected by Miguel Angel Meza Ortega; Shelton P. Applegate, Gerardo González Barba, Andreas Knieth and Lawrence G. Barnes February 1993.
Formation. El Cien Formation.
Age. 28 to 25 Ma approximately, Late Oligocene (Fischer et al., 1995).
Description. The specimen (Figure 5A) consists of a partial skull with a preserved length of ~400 mm (substantially less than condylobasal lengthbecause of loss of the rostrum apex), bizygomatic width estimated at ~200 mm, and estimated body size of ~1. 41 m (following the equation for stem Odontoceti in Pyenson and Sponberg, 2011). The inferred age class is adult, as indicated by completely closed cranial sutures, a developed nuchal crest, and epiphyses fused to the cervical vertebra body (on significance of fusion, see Perrin, 1975). MU EcSj5/12/48 was earlier referred to as “Agorophius ” sp. (Cruz-Marín, 1997), but its morphology is quite different to Agorophiidae (Fordyce, 1981, Godfrey et al., 2016) and closer to Simocetus rayi Fordyce, 2002. Amongst features of note, the specimen is small compared with both S. rayi (condylobasal length: 449+ mm; bizygomatic width: ~238 mm, Fordyce, 2002) and Agorophius pygmaeus (Müller, 1849) (condylobasal length: ~450-500 mm; bizygomatic width: ~200+ mm). The skull has a shortened rostrum with a narrow snout and short mesorostral groove, and a supraoccipital shield with a semicircular outline behind the anterior margin of the squamosal fossa. The intertemporal region is prominent. The small teeth are heterodont but not polydont. The maxilla rises posteriorly over the frontal, extended near to the frontoparietal suture; laterally overlaps part of the supraorbital process, but does not reach the supraorbital margin, leaving the rest of the frontal exposed dorsally above the elevated roof of the orbit. The premaxillary sac fossa and associated foramina and sulci lie well forward on the rostrum, anterior to the antorbital notch. Many of the latter are plesiomorphic features shared with S. rayi and A. pygmaeus, and for now they do not help with taxonomy. Systematic questions should be resolved by a detailed morphological description and phylogenetic analysis.
Family indet. gen. et. sp. indet.
Material. MNH-UABCS_EcSj24/50/183, skull (Figure 5B).
Locality. San Juan de la Costa, Arroyo Tarabillas, mine spoils (24. 44249° N, 110. 69106° W) La Paz, Baja California Sur, México. Collected by Robert Ewan Fordyce; Gerardo González Barba; Priscila Morales Ortega; Ricardo Mirsha Martín Mata Cruz, and Atzcalli Ehécatl Hernández Cisneros. February 2008.
Formation. El Cien Formation.
Age. 28 to 25 Ma approximately, Late Oligocene (Fischer et al., 1995).
Description. The specimen (Figure 5B) consists of a partial skull with a preserved length of ~305 mm but judging from comparable specimens (USNM 205491, below) its condylobasal length could have been double the preserved length. The inferred age class is adult, as indicated by cranial sutures which are relatively closed and tight. Features of note include the prominent narial “snout”, and archaic features like parietals exposed at a prominent intertemporal constriction, plus premaxilla without a posterolateral fold but with a narrow and elongate premaxillary sac fossa (Fordyce et al., 2012). The skull is more dorsoventrally flat than S. rayi Fordyce (2002), Albertocetus meffordorum Uhen (2010), Waipatiidae (Fordyce, 1994; Tanaka and Fordyce, 2014, 2015a, 2015b), Patriocetus ehrlichi (Van Beneden, 1865), Patriocetus kazakhstanicus, Dubrovo and Sanders, 2000, or the unnamed specimen USNM 205491 from Oregon (Fordyce et al., 2012). The rostrum is less concave and, the premaxillary sac fossae are more anterior and shallow than S. rayi. Other features include: the narrow and relatively long nasals, an elongate and narrow postorbital process of the frontal; and the flat, wide and slightly concave palatine. Fordyce et al. (2012) identified the specimen as a possible Dalpiazinidae, but the typology of that family is now considered uncertain, and the specimen is not allocated to the family.
4. 1. 5. Suborder ‘Archaeoceti’ ?Kekenodontidae
Archaeoceti Flower, 1883
?Kekenodontidae Mitchell, 1989
?Kekenodontidae gen. et. sp. indet.
Material. MHN-UABCS_EcSj5/16/267, left periotic (Figure 6A).
Locality. San Juan de la Costa, El Saladito, mine spoils (24. 44249° N, 110. 69106° W), La Paz, Baja California Sur, México. Collected by Gerardo González Barba; Lawrence G. Barnes, and Arturo Cruz Marin. March 1993.
Formation. El Cien Formation.
Age. ~27 Ma approximately, Late Oligocene (Fischer et al., 1995).
Description. The specimen (Figure 6A) is a left periotic without the posterior process, and with a shape similar to that of basilosaurids (Kellogg, 1936; Luo and Gingerich, 1999; Uhen, 2004; Fitzgerald, 2010), and contrast widely with the morphology present in mysticetes and odontocetes (Hernández-Cisneros and Tsai, 2016). The periotic has a maximum preserved length of 51 mm. Features of note include the elongate body which is transversely narrow in ventral view, like Kekenodon onamata Hector, 1881 (see Hector’s Fig. 10), and is quite similar to the undescribed specimen OU 22294 (Fordyce 2004; Clementz et al., 2014) from New Zealand. The suprameatal area is ovoid; the superior process is low compared with the high structure seen in Zygorhiza kochii (Reichenbach, 1847), Dorudon atrox (Andrews, 1906), or Basilosaurus isis (Andrews, 1904). In lateral view the shape is rectangular, the superior process has a concave profile and the anterior process is trapezoidal-rectangular in medial view. The periotic, MU EcSj5/16/267, is similar to the elements of the unprepared specimen, MU EcSj1/45/214, but it is not clear whether these are the same species; detailed description and comparison are needed. Other unprepared specimens, such as MU EcSj5/16/72 (partial frontal, Figure 6B); MU EcSj1/94/296 and MU EcSj1/38/155 (partial skulls), are phenetically similar to the undescribed specimen OU 22294 (Clementz et al., 2014: Fig. 2) in a narrow and longest intertemporal region, short zygomatic process, and a flat braincase (Hernández-Cisneros, 2015).
5. Discussion
Cetaceans from Baja California Sur contribute towards the continually expanding Late Oligocene history of Cetacea around the Pacific Basin. Barnes (1998) briefly discussed apparent differences of the Oligocene cetaceans from Mexico with other areas of the Pacific margin, remarking upon similarity with the fossil cetacean assemblage from the Olympic Peninsula in Washington (U. S. A.) and differences with Oligocene cetaceans from Hokkaido (Japan), and New Zealand. Given the rapidly expanding diversity of Oligocene Cetacea from Baja California Sur and New Zealand, comparisons may become outdated unless species are formally described.
At present, the reported Oligocene cetaceans from the Pacific Basin include several formally named species (Appendix, Table A1). Many recent advances in cetacean paleontology have expanded the framework of Oligocene cetacean evolution and its diversity, e. g. Fitzgerald (2010); Boessenecker and Fordyce (2014, 2015a, 2015b); Tsai and Fordyce (2015); Tanaka and Fordyce (2014, 2015a, 2015b), and others. The nature of the specimens means that many of the latter articles are detailed morphological accounts with cladistic analyses of large character matrices. In spite of such advances, there is a much greater known diversity -as revealed by undescribed museum collections- than is apparent from the literature. Thus, there is a considerable information gap about the Oligocene cetaceans (Uhen and Pyenson, 2007), which include a critical link between archaeocetes and the Neogene cetacean groups (Fordyce et al., 1995). The Late Oligocene cetacean fossils from Baja California Sur will elucidate evolution of the Cetacea including morphological transitions, functional and ecological shifts, and changing biogeography.
6. Conclusion
Baja California Sur has produced some important Cenozoic fossils assemblages distributed along the Peninsula, including Late Oligocene marine mammals (mostly cetaceans and a few desmostylians) reported briefly in the last 70 years. Published details are sparse, and some appear only in theses or abstracts. These fossils represent the southern-most records of Late Oligocene cetaceans from North America. There is a great expectation to increase the knowledge of cetacean diversity as fieldwork continues and the specimens collected in museums are prepared. Recent studies indicate the presence of hitherto unreported archaic odontocetes, mysticetes (Aetiocetidae and Chaeomysticeti), and kekenodontid-like cetaceans, mostly from the El Cien Formation (Oligocene-Early Miocene) and San Gregorio Formation (Late Oligocene). As with new fossil cetaceans from elsewhere in the world, the materials will expand understanding of diverse aspects of cetacean evolution.





