











 text new page (beta)
text new page (beta) English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO 
 Permalink
Permalink1. Introduction
Two small Ophthalmoplax specimens were collected from the upper portion of the Lodolitas de Aguacaliente Formation (Guerrero and Sarmiento, 1996), within outcrops of the eastern margin of the Cordillera Oriental de Colombia (Fig. 1). The specimens represent the oldest record of the genus in America. Their small size may be related with an anoxic event during the late Campanian in Colombia (Föllmi et al , 1992; Martinez, 1995; Guerrero et al , 2000; Guerrero, 2002), and they represent a new species of the genus, named O. andina new species.

Figure 1 Location map of the Llanos foothills, central region of Colombia, with position of fossil locality.
Feldmann et al (1999) reported Ophthalmoplax spinosus Fedmann, Villamil and Kauffman, 1999 from the Turonian San Rafael Formation, near Villa de Leyva, Boyacá department; but subsequently, Vega et al (2007) assigned this specimen to Cenomanocarcinus vanstraeleni Stenzel, 1945. Later, Feldmann and Villamil (2002) described Ophthalmoplax triambonatus Feldmann and Villamil, 2002 from the Puerto Romero Formation, near the town of the same name, west of Boyacá department; latest Maastrichtian age is now accepted for the Puerto Romero Formation, since this unit forms at the top of the Cretaceous sequence of the Middle Magdalena Basin (Vásquez-Parra et al , 2000). Vega et al (2013) illustrated another specimen of Ophthalmoplax (of possible latest Maastrichtian age) from the Tolima department. According to Vega et al (2013), these two specimens, of relative large size, correspond to Ophthalmoplax brasiliana (Maury, 1930), a taxon originally described based on large-sized specimens from the latest Maastrichtian Gramame Formation, Eastern Brazil (Maury, 1930; Beurlen, 1958). Vega et al (2013) suggested that Ophthalmoplax is monospecific in America (O. brasiliana ), with a stratigraphic range throughout the Maastrichtian and a geographic distribution from Brazil to North Carolina, with O. stephensoni Rathbun, 1935 and O. triambonatus as synonyms of O. brasiliana O. minimus Ossó-Morales, Artal and Vega, 2010 (Ossó-Morales et al , 2010) from the late Campanian of Morocco is the other valid species for the genus.

Figure 2 Stratigraphic section of Lodolitas de Aguacaliente Formation (modified from Guerrero and Sarmiento, 1996), showing position of segment containing crustacean specimens, sequence stratigraphy surfaces, system tracts and paleoenvironments
2. Geographic and geological setting
The study area is located 100 Km east of Bogotá D.C., at the southern margin of the Boyacá department, on approximately 5 km west of the San Luis de Gaceno county, eastern margin of the Cordillera Oriental de Colombia. The fossiliferous level belongs to the Lodolitas de Aguacaliente Formation, middle portion of Guadalupe Group, it thus forms part of the Upper Cretaceous sequence of the Piedemonte Llanero area. Exposures are along San Antonio creek, a tributary of Lengupá River (Fig. 1).
3. Stratigraphy
Available biostratigraphic data suggest a late Campanian age for Lodolitas de Aguacaliente Formation. This is based on the presence of: benthic foraminifers Bolivinoides decoratus decoratus Jones in Wright, 1886 and Haplophragmoides cf. perexplicatus ; a rich dinoflagellate association including Andalusiella polymorpha polymorpha Malloy, 1972 and A. polymorpha aegyptiaca Schrank, 1988; the abundance and posterior disappearance of Trichodinium castanea Deflandre 1935; and gradual disappearance in number of species and specimens of Dinogymnium Evitt, Clarke and Verdier, 1967 (Guerrero and Sarmiento, 1996; Tchegliakova et al , 1997). In the center of the basin, the transgressive strata of fully marine correlative unit Plaeners Formation, shows the presence of ammonoids Nostoceras (Nostoceras ) liratum sp., and Exiteloceras jenneyi Whitfield, 1877, and the buliminid foraminifera Siphogenerinoides ewaldi Karsten, 1858, that also indicates a late Campanian age (Föllmi et al , 1992; Martinez, 1995).
The Lodolitas de Aguacaliente Formation conformably overlies the Arenitas de San Antonio Formation (lower Campanian). The latter composed, at the top, by medium to coarse-grained quartz arenites, deposited in a lower shoreface environment at the eastern margin of the Cretaceous Colombian Basin (Guerrero and Sarmiento, 1996). The Lodolitas de Aguacaliente Formation (160 m thick), is composed dominantly by mudstone lithofacies, although diagenetic chert, quartz arenites (some glauconitic), and thin-bedded phosphorites are also frequent. Ellipsoidal concretions, some pyritized, and highly bioturbated levels are also present. Fecal pellets, fish scales and occasional bivalves, gastropods and lingulid brachiopods are common.
The contact between Lodolitas de Aguacaliente and Arenitas de San Antonio formations is marked by the change from a coarsening-upward to a fining-upward trend (shallowing to deepening) and also a variation in the parasequences stacking pattern from progradational to retrogradational; this change represents a maximum regressive surface (MRS sensu Embry, 2009) or transgressive surface, that separates the Arenitas de San Antonio Formation regressive strata (representing a Regressive systems tract - RST), from the base of Lodolitas de Aguacaliente Formation (which represents a Transgressive systems tract - TST).
The specimen of Ophthalmoplax here described was discovered in layers that represent the uppermost parasequences with the retrogradational stacking pattern, close to the maximum flooding surface (MFS) it was detect to the end of the Upper Campanian.
4. Material
Two specimens is deposited in the Paleontological Collection of the Universidad Nacional de Colombia, Facultad de Ciencias, Departamento de Geociencias, Bogotá D.C., Colombia, under catalog numbers UN-DG-CR-021 and UN-DG-CR-022.
5. Systematic Paleontology
Order Decapoda Latreille, 1802
Infraorder Brachyura Latreille, 1802
Superfamily Portunoidea Rafinesque, 1815
Family Macropipidae Stephenson and Campbell, 1960
Genus Ophthalmoplax Rathbun, 1935
Type species. Zanthopsis brasiliana Maury, 1930, by subsequent designation of Vega et al , 2013.
Ophthalmoplax andina new species Figs. 3-5A

Figure 3 Ophthalmoplax andina n. sp., Lodolitas de Aguacaliente Formation, late Campanian. A) Dorsal carapace morphology, holotype UN-DG-CR-021. B) Dorsal carapace morphology, paratype UN-DG-CR-022. Scale bars = 1cm.
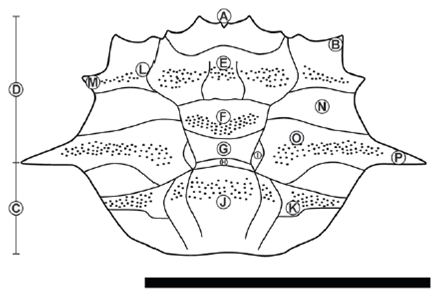
Figure 4 Line drawing of Ophthalmoplax andina n. sp., based on holotype UN-DG-CR-021, sp. A) Bifid rostrum, B) Orbital spines, C) Posterolateral margin, D) Anterolateral margin, E) Protogastric region, F) Mesogastric region, G) Metagastric region, H) Urogastric region, I) Branchial lobes, J) Cardiac region, K) Metabranchial region, L) Hepatic region, M) Anterolateral spine, N) Epibranchial region, O) Mesobrachial region, P) Lateral spine. Scale bar = 1cm.

Figure 5 Ophthalmoplax spp. from different localities and ages. A) Ophthalmoplax andina n. sp., holotype UN-DG-CR-021, upper Campanian, Colombia. B) O. minimus , upper Campanian, ~76 Ma, Morocco. C-H) O. brasiliana , Upper Cretaceous of the Americas. C) Lower Maastrichtian, ~ 70 Ma, Mississippi, USA. D) Lower Maastrichtian, ~ 70 Ma, North Carolina, USA. E) Upper Maastrichtian, ~ 66.8 Ma, Colombia. F) Upper Maastrichtian, ~ 66.8 Ma, North Carolina, USA. G) Uppermost Maastrichtian, ~ 66.2 Ma, Mexico. H) Uppermost Maastrichtian, ~ 66.2 Ma, Mississippi, USA. Scale bars = 1cm.
Diagnosis. Carapace small, wider than long, subquadrate; rostrum bifid; orbital spines sharp and short. Dorsal carapace surface mostly defined by sharp transverse ridges and grooves, deep grooves separate this area from mesobranchial region; mesobranchial region with wide transverse ridges with fine tubercles on crests, connected to long, sharp lateral spine.
Description. Carapace small, wider than long, subquadrate, maximum width at mesobranchial region; dorsal surface longitudinally convex; rostrum bifid; orbito-frontal margin wide, with deep fissure separating rostrum from orbital spines; orbital spines sharp and short; anterolateral margin 60 % the maximum carapace length, slightly concave; posterolateral margin convex, longer than anterolateral margin; posterior margin convex, slightly shorter than orbito-frontal margin. Dorsal carapace surface mostly defined by sharp transverse ridges and grooves; protogastric region swollen, with sharp transverse ridge bearing tubercles on crest, separated from hepatic region by sharp longitudinal groove; mesogastric region subovate, with transverse ridge bearing fine tubercles on crest, delimited from epibranchial region by deep longitudinal grooves; smooth mesogastric region inverted subtrapezoidal, deep grooves separate this area from mesobranchial region; urogastric region forms a shallow depression, narrow and smooth, subrectangular in shape, bounded by narrow, well-defined elongate branchial lobes, separated by deep grooves from adjacent branchial region; cardiac region subtrapezoidal, with sharp, median transverse ridge with fine tubercles on crest; intestinal region undifferentiated; hepatic region with narrow transverse ridge, connected laterally with sharp and long anterolateral spine; epibranchial region with slightly convex subovate lobes, bounded by grooves and lobes of adjacent regions; mesobranchial region with wide transverse ridges with fine tubercles on crests, connected to the long, sharp lateral spines; metabranchial region undifferentiated; no ventral region nor appendages are preserved.
Etymology. The name of the new species is related to the Andean region of Colombia.
Material. Two small carapaces, holotype UN-DG-CR-021; paratype UN-DG-CR-022.
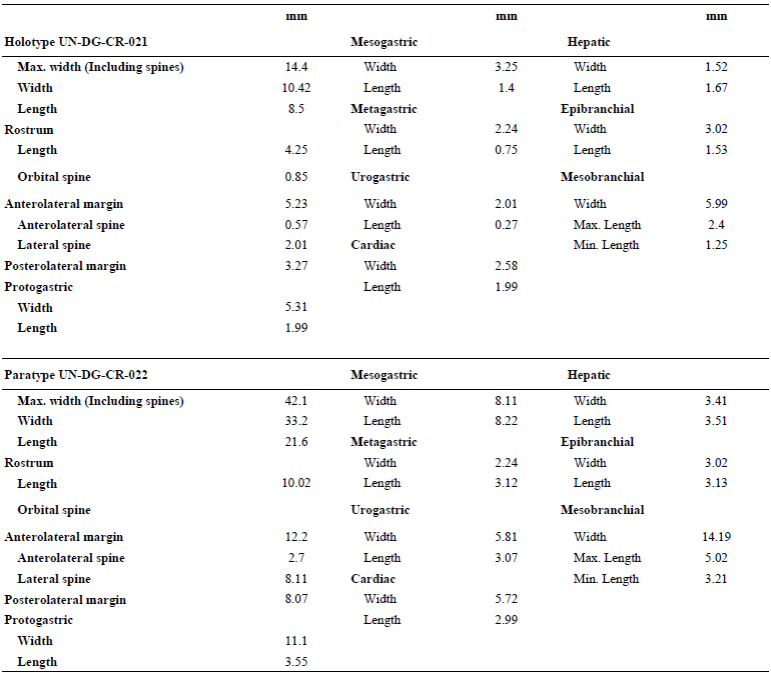
Measurements. Measurements of the specimens are given in Table 1.
Table 1 Measurements (in mm) of holotype UN-DG-CR-021 and paratype UN-DG-CR-XXX of Ophthalmoplax andina n. sp.
Discussion. Maury (1930) reported Ophthalmoplax brasiliana from the Upper Maastrichtian Gramame Formation, Brazil (Table 2). Beurlen (1958), based upon vague morphological differences, suggested concordance of O. brasiliana with O. stephensoni Rathbun, 1935. Feldmann and Villamil (2002) described O. triambonatus from Colombia; however, the single specimen is very similar to O. brasiliana and they both are latest Maastrichtian in age. Rathbun (1935) and Schweitzer et al (2007) in describing specimens of O. stephensoni , presented identical characteristics to those observed in specimens of O. brasiliana Aguilera et al (2010) described O. triambonatus from the late Maastrichtian of Venezuela but again that species has identical characteristics to those observed in specimens of O. brasiliana Finally, Vega et al (2013) described Ophthalmoplax sp. from the Maastrichtian of Tolima department and it too has identical characteristics to those observed in specimens of O. brasiliana According to Vega et al (2013), O. stephensoni is a synonym of O. brasiliana (Vega et al , 2013). Moreover, Mascaranada difuntaensis described by Vega and Feldmann (1991) from the early Maastrichtian Potrerillos Formation, NE Mexico, corresponds to O. brasiliana Ophthalmoplax spinosus from the San Rafael Formation (Turonian), described by Feldmann et al (1999) was redefined as Cenomanocarcinus vanstraeleni by Vega et al (2007).
Table 2 Stratigraphic and geographic distribution of genus Ophthalmoplax , including original definition and subsequent redefinition* (see in text).

Ophthalmoplax has been documented from North Africa, where the genus is present in Upper Cretaceous rocks of the Moyenne Mouluya region of northeast Morocco. It was here described as O. minimus and represents the first record for Opthalmoplax in the west Tethyan Realm. 87Sr/86Sr isotopic data obtained from the well-preserved cuticle of O. minimus suggests an absolute age of 76 Ma, corresponding to the base of the late Campanian (Ossó-Morales et al , 2010). Vega et al (2013) proposed that Ophthalmoplax was represented by only one species in America (O. brasiliana ) of early to latest Maastrichtian age, which was distributed from Brazil to North Carolina, thus invalidating O. stephensoni and O. triambonatus (Fig. 5). To date (in the Americas), the oldest record of Ophthalmoplax corresponded to the Early Maastrichtian (O. brasiliana ). The new Colombian Ophthalmoplax sp. from late Campanian Lodolitas de Aguacaliente Formation of Cordillera Oriental of Colombia, is here considered to be older and may represent the oldest record for the genus in America.
The new species of Ophthalmoplax is similar to O. brasiliana , but is different for the presence of extremely long lateral spines and size (even smaller than O. minimus ).
The small size may be related to temperature fluctuations. Based on calcareous nannofossil abundance (Bice et al. , 2006; Forster et al , 2007; Voigt et al , 2004; Wilson et al , 2002; Linnert et al , 2011), it is known that between Turonian to Maastrichtian times, there was a continuous cooling of seawater, preceded by one of the warmest periods of the Phanerozoic (early Cenomanian - Turonian). In this scenario, global temperatures decreased between 4 - 6 °C during the late Campanian (73 - 70 Ma) (Li and Keller, 1999). According to Keller (2008), low temperatures enhance diversity in relation to increased nutrient cycling and the stratification of the water masses (Fig. 6).
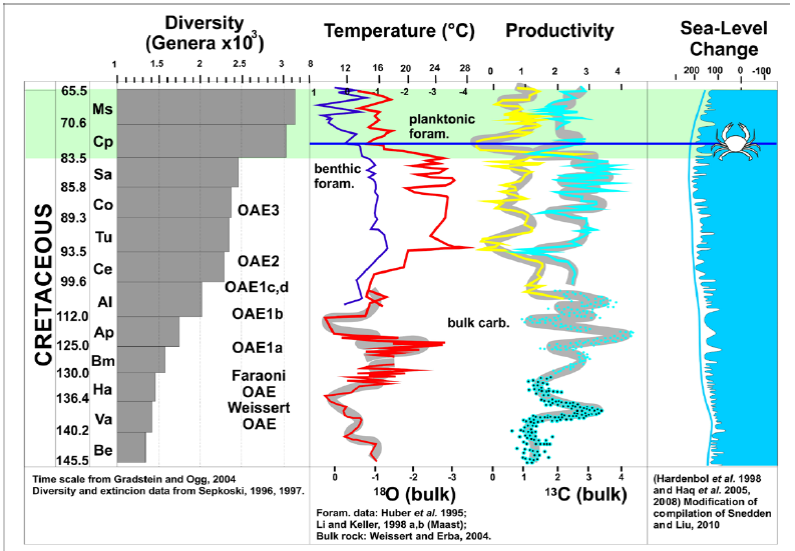
Figure 6 Cretaceous temperature and carbon isotopic (productivity) trends compared with diversity and relative sea level change. Modified from Keller (2008) and Snedden and Liu (2010). Note position of Ophthalmoplax andina n. sp. in a high sea-level period, cool climate and low values of productivity related to increased nutrient cycling and watermass stratification.
The stratigraphic and bioestratigraphic evidences present in the Lodolitas de Aguacaliente Formation suggest that the unit was deposited during a phase of relative sea-level high (Guerrero and Sarmiento, 1996; Tchegliakova et al , 1997). Specifically, the horizon containing Ophthalmoplax andina n. sp. is located towards the end of a transgressive period (TST), at the late Campanian MFS, which can be associated with poor oxygenation and widespread anoxia throughout the Cretaceous Colombian Basin, according to observed variations between benthic foraminifera of the Upper Cretaceous Guadalupe Group (Fig. 7). When populations were grouped into infaunal and epifaunal preferences, it was found that the available oxygen levels at the sea bottom could have controlled their distribution. The evidence suggests that the associations dominated by the epifaunal morphogroup indicate low oxygen levels (Tchegliakova et al , 1997), as observed also in the O. andina n. sp. level of the Lodolitas de Aguacaliente Formation. The latter scenario prevailed during the time of maximum transgression of the middle-upper part of Lodolitas de Aguacaliente Formation, which is a correlative unit with the Plaeners Formation (central sector of the Colombian Cretaceous Basin), where in addition to anoxia, high acidity is evidenced (Cruz et al , 2011) based on chemical characterization of foraminifera tests.
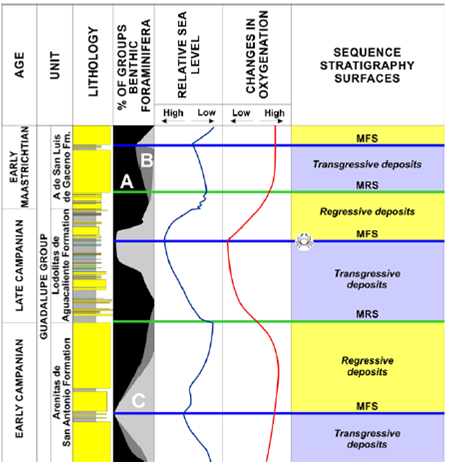
Figure 7 Stratigraphy, paleoenvironments, relative sea level, oxygen level changes and sequence stratigraphy surfaces at Guadalupe Group section (Campanian-Maastrichtian). Modified from Tchegliakova et al (1997). A= Detritivorous benthic foraminifera with elongated test morphotypes, and preferential infaunal micro-habitat; B= Detritivorous/omnivorous/ herbivorous benthic foraminifera with wide test morphotypes, with preferential epifaunal to infaunal micro-habitat; C= Suspension feeder benthic foraminifera with tubular or branches unilocular test morphotypes, and preferential epifaunal micro-habitat.
Sea-level changes, presence of an epeiric sea, stratification of water column, possible vulcanism which increased water acidity, coupled with an increase and diversification of life in the oceans, greatly influenced anoxic conditions in many basins worldwide, and appear to be directly related to the small size of the Campanian Ophthalmoplax andina n. sp. in South America and North Africa. O. brasiliana reached maximum sizes (over 12 cm in carapace width) by late Maastrichtian (66.8 Ma) times, when sea temperatures were much lower and had influence on the size of these portunoid crabs (Vega et al , 2013). Along with low temperatures for early to late Maastrichtian sea water, different environmental conditions in oceans with greater mobility and increased oxygenation prevailed for the Middle and Late Maastrichtian (Douglas and Savin, 1973; Arthur et al , 1985; D'Hondt and Lindinger, 1994; Barrera et al , 1997; Frank and Arthur, 1999; Friedrich et al , 2004, among others).
6. Conclusions
Ophthalmoplax andina n. sp. from the Upper Campanian Lodolitas de Aguacaliente Formation of Colombia represents the stratigraphically oldest report for the genus in America, as well as the smallest Ophthalmoplax reported to date. The size reduction may be related to anoxic conditions that prevailed during a period of maximum marine flooding (MFS) at the end of late Campanian TST. We suggest that the small size resulted from environmental adaptative pressures, related to anoxic conditions.

