










Services on Demand
Journal
Article
 text in
text in  English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkAgrociencia
On-line version ISSN 2521-9766Print version ISSN 1405-3195
Agrociencia vol.52 n.8 Texcoco Nov./Dec. 2018
Animal Science
Productive performance of Lotus corniculatus L. dependent on harvest strategy
1Recursos Naturales Renovables. Universidad Autónoma Agraria Antonio Narro. Calzada Antonio Narro 1923, Buenavista, Saltillo Coahuila.
2Producción de Semillas. Campus Montecillo. Colegio de Postgraduados. 56230. Montecillo, Estado de México.
3Desarrollo Agrícola Regional. Campus Puebla. Colegio de Postgraduados. 72760. Puebla, Puebla, México.
4Ganadería. Campus Montecillo. Colegio de Postgraduados. 56230. Montecillo, Estado de México.
Harvest efficiency is one of the main factors that define productivity of a grassland. In Lotus species, productive response has not been studied in function of the amount of intercepted light. This study was conducted in the Colegio de Postgraduados, Montecillo, Texcoco, Mexico, in the 2014-2015 and 2015-2016 crop cycles with the objective of determining the production of L. corniculatus in function of the percentage (90, 95 and 100 %) of light intercepted (LI) and a fixed cutting (CF) defined seasonally. The hypothesis was that higher production of forage would be obtained with 95 % LI. The experimental design was random blocks in divided plots, and the treatments were LI and CF with three replications. Annual yield of forage showed differences among treatments only in the second year. Forage yield of CF was lower (22 844 kg MS ha-1) than the average with 90, 95 and 100 % LI (32 173 kg MS ha-1). Seasonal yield was higher in spring (10 461 kg MS ha-1) relative to the fall and winter average (5750 kg MS ha-1). Weeds contributed (4 %) to annual yield and leaves (58 %) in spring with 95 % LI. With 95 and 100 % LI greater heights (23 cm) were recorded, and in CF the heights were lower (19 cm) in both years. Of the seasons, taller plants (24 cm) grew in the spring and shorter plants in autumn and winter (19 cm). In the second year, there was a higher mean annual leaf: stem ratio in CF (2.8:1), mainly in autumn (3.3:1). The best productive performance of L. corniculatus was with 90, 95 and 100 % LI. The highest contribution of leaf was with 95 % LI, mainly in spring. In conclusion, although there were not differences between the percentages of intercepted light, at 95% the largest amount of leaf was produced, and thus we consider it to be the best management strategy. Nevertheless, because it is a species recently characterized in Mexico, it is necessary to conduct more research in different environment types.
Keywords: Lotus corniculatus L.; forage yield; productive performance; intercepted light; cutting interval
La eficiencia de la cosecha es uno de los principales factores que definen la productividad de una pradera. En especies de Lotus no se ha estudiado la respuesta productiva en función de la cantidad de luz interceptada. Este estudio se realizó en el Colegio de Postgraduados, Montecillo, Texcoco, México, en los ciclos agrícolas 2014-2015 y 2015-2016, y el objetivo fue determinar la producción de L. corniculatus, en función del porcentaje (90, 95 y 100 %) de luz interceptada (LI) y un corte fijo (CF) definido estacionalmente. La hipótesis fue que la mayor producción de forraje se presenta con 95 % de LI. El diseño experimental fue bloques al azar en parcelas divididas, y los tratamientos fueron LI y CF con tres repeticiones. El rendimiento anual de forraje solo mostró diferencias entre tratamientos en el segundo año. El rendimiento de forraje del CF fue menor (22 844 kg MS ha-1) al promedio con 90, 95 y 100 % LI (32 173 kg MS ha-1). El rendimiento promedio estacional de forraje fue mayor en primavera (10 461 kg MS ha-1) respecto al promedio de otoño e invierno (5750 kg MS ha-1). La maleza aportó al rendimiento anual (4 %) y la hoja el (58 %) en primavera, con 95 % de LI. Con 95 y 100 % de LI se registraron las alturas mayores (23 cm) y en el CF las alturas menores (19 cm), en ambos años. Entre estaciones, las alturas mayores (24 cm) se presentaron en primavera y las menores en otoño e invierno (19 cm). En el segundo año se presentó la mayor relación media anual hoja: tallo en el CF (2.8:1), principalmente en otoño (3.3:1). El mayor comportamiento productivo de L. corniculatus fue con 90, 95 y 100 % de LI, con aporte mayor de hoja con 95 %, principalmente en primavera. En conclusión, a pesar de, no haber diferencia entre las intercepciones luminosas, al 95 % se produjo la mayor cantidad de hoja, por lo que, podríamos considerarla como la mejor estrategia de manejo, no obstante, siendo una especie recién caracterizada en México, es necesario realizar más investigación en diferentes tipos de ambientes.
Palabras clave: Lotus corniculatus L.; rendimiento de forraje; comportamiento productivo; luz interceptada; intervalo de corte
Introduction
The genus Lotus comprises around 200 annual and perennial species. The European Mediterranean region has the highest diversity (Lagler, 2003), and L. corniculatus is the most important forage species (Castro, 2006). It is distributed in temperate regions of Europe, the Near East, and North and South America (Gunn et al., 1992). This leguminous plant grows in temperate climates, and its yield and nutritional quality (18 to 22 % crude protein) is comparable to Medicago sativa L. and Trifolium repens L. (Marley et al., 2006). Moreover, it contains less cellulose and more nonstructural carbohydrates (Grant, 2009) and does not produce bloating in grazing animals (Beuselinck and Grant, 1995).
Productivity and persistence of pastures is in function of their capacity to recover and maintain the remaining leaf area. Different harvest strategies have an impact on the plants and determine their growth rate, productivity and persistence (Nabinger and Pontes, 2001). For this reason, harvest efficiency is one of the main factors that define productivity of a pasture (Giacomini et al., 2009), which is affected by environmental conditions such as the amount of light (Lemare and Agnusdei, 2000) that can increase competition among plants. In the periods of regrowth, the quantity and quality of light in pastures decrease (Da Silva and Nascimento, 2007). One of the criteria for estimating the optimum point for harvesting is when the pasture reaches 95 % intercepted light (Montagner et al., 2012). Consequently, good management of the amount of light intercepted by the pasture assures better productivity (Difante et al., 2008). Information on L. cornicultatus is scarce, but in other temperate legumes there are high correlations between forage yield and the amount of intercepted light (Rojas et al., 2016). Therefore, the objective of the present study was to evaluate the productive performance of the genotype 202700 of L. corniculatus in function of the amount of intercepted light and a fixed cutting interval defined seasonally.
Materials and methods
Field conditions
The experiment was conducted at the Colegio de Postgraduados, Campus Montecillo, Texcoco, México (19° 29’ N and 98° 53’ W), altitude 2,250 masl, during the 20142015 and 2015-2016 crop cycles. The climate is sub-humid, with and mean temperature of 15 °C and annual accumulated precipitation of 636 mm (García, 2004). Soil is sandy loam with pH 7.1. Climate data were obtained from the meteorological station of the Universidad Autónoma Chapingo, 2 km from the study site. Accumulated annual precipitation was 1043 mm in the first year and 877 mm in the second. Higher precipitations and temperatures were recorded during the spring and summer than in fall and winter in both years (Figure 1).
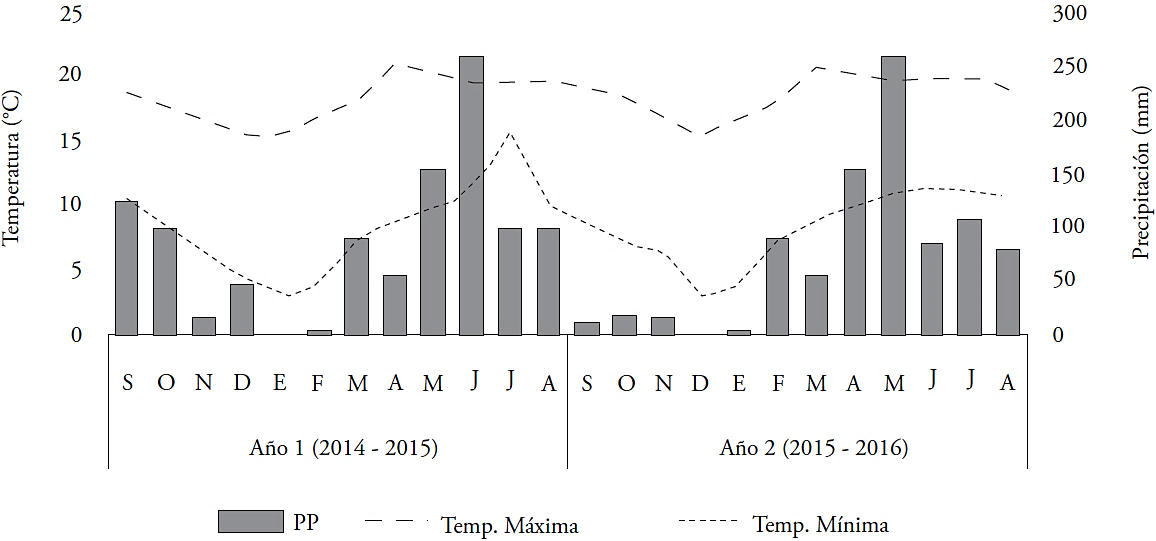
Figure 1 Mean monthly high and low temperatures and monthly accumulated rainfall (PP) in the two crop cycles.
The pasture of L. corniculatus was established by transplanting on March 14, 2014; density was 9 plants m-2 and distance between plants was 33 cm. The plants were obtained from seedlings grown in a greenhouse in a substrate of soil typical to the study site with no fertilization. Before beginning the experiment, sheep were grazed 13 weeks after establishment and uniformizing cutting was done at the beginning of the experiment (September 5, 2014) to 7 cm from the soil. The plots were irrigated every 15 d to field capacity in the dry season; no fertilizer was applied.
The experimental area was divided into 12 sub-plots with an area of 4 m2 with 36 plants per experimental unit. Cuttings were treatments and were determined when the pastures reached percentages of intercepted light of 90, 95 and 100 %, plus a fixed cutting defined seasonally (spring-summer: every 28 d, fall: every 35 d, and winter: every 42 d). LI was monitored at soil level (six measurements) using a canopy analyzer, AccuPAR Linear PAR/ LAI ceptometer Modelo PAR 80 (Decagon devices, USA).
Forage yield (kg DM ha-1) was assessed when biomass was harvested in two fixed squares (0.25 m2) by repetition, established at the beginning of the experiment. This forage was dried at 60 °C to constant weight in a forced-air oven (Felisa, Mod. FE-243A). Botanical and morphological composition (CBM) was calculated by taking a 10 % sample of the harvested forage, separating it into leaf, stem and dead matter (senescent material) and weeds. The contribution to yield of each component (kg MS ha-1) was calculated. Plant height (cm) was measured in 12 random readings per replication using a 50 cm graduated ruler. The equivalence in yield per cm of plant height was calculated by dividing yield by the cm of plant height by the number of cuttings. The CBM data for leaf and stem were used to determine the leaf/stem ratio (R:L/S: 1), dividing leaf weight by stem weight.
The treatments were assigned to experimental units according to a design of random blocks in divided plots with three replications, where the large plot was the genotype and the small plot the treatment. The data obtained per cutting were organized by season and analyzed with PROC GLM of SAS (SAS Institute, 2009); averages were compared with the Tukey test (p≤0.05).
Results and discussion
Forage yield
Differences (p≤0.05) in annual forage accumulation were found only in the second year (Table 1), and the lowest yield corresponded to the fixed cutting with 29 % less than the average of the three percentages of intercepted light (32,173 kg MS ha-1), indicating less adaptation of the species to fixed cutting management, relative to the cuttings that depended on the percentage of intercepted light. Regardless of the treatment, the highest mean seasonal yields, in both years, were obtained in spring with 10,461 kg MS ha-1 and the lowest in fall (5,915 kg MS ha-1) and winter (5,585 kg MS ha-1). This behavior can be explained by the direct and close relationship between forage yield and optimal temperatures (22 °C) for growth of the species in field conditions (García et al., 2014). Optimal temperatures occurred in spring (Figure 1) benefitting L. corniculatus growth and productivity. In our study, seasonal behavior is similar to forage yield reported by Scheffer et al. (2011) of five populations of L. corniculatus, but unlike the behavior with fixed cutting established in our study, these authors observed that a high cutting frequency increases yield. Sbrissia et al. (2010) attribute these changes in yield to environmental conditions in each season of the year. Díaz et al. (1996) found a seasonal distribution of L. corniculatus forage production of 49, 25, 14 and 12 % for spring, summer, winter and fall, respectively, with 72% in spring-summer. According to Santiñaque and Battista (2003), there is an increase in yield of 45% at harvest intervals of 44 d, relative to that at 22-d intervals. For 45-d cutting intervals, García et al. (2014) observed an average forage yield of 7700 kg MS ha-1, and the genotype 202700 was the most productive of the 12 L. corniculatus genotypes evaluated. They also pointed out that accumulated production can be influenced by climate, management and the growing habit of the genotype.
Table 1 Accumulated forage yield of L. corniculatus, genotype 202700, in function of intercepted light (LI) and a fixed cutting defined seasonally.
| LI (%) | Otoño | Invierno | Primavera | Verano | Anual acumulado | EEM† |
| Año 1 (2014 - 2015) | ||||||
| 90 | 4881 Ad | 6447 Ac | 10396 ABa | 8593 ABb | 30317 A | 516 |
| 95 | 5533 Ad | 6757 Ac | 10806 Aa | 9177 Ab | 32274 A | 393 |
| 100 | 5785 Ab | 6453 Ab | 11116 Aa | 9880 Aa | 33234 A | 823 |
| Corte fijo§ | 4896 Ac | 6568 Ab | 8708 Ba | 7509 Bb | 27680 A | 373 |
| Promedio estacional | 5274 d | 6556 c | 10256 a | 8790 b | 30876 | 369 |
| EEM† | 721 | 864 | 673 | 546 | 2303 | |
| Año 2 (2015 - 2016) | ||||||
| 90 | 5695 ABc | 5729 Ac | 11063 Aa | 8797 Ab | 31283 A | 549 |
| 95 | 6567 Ac | 6293 Ac | 12068 Aa | 8773 Ab | 33701 A | 265 |
| 100 | 5398 ABc | 5688 Ac | 11433 Aa | 9015 Ab | 31534 A | 762 |
| Corte fijo§ | 4934 Bb | 4377 Bb | 8222 Ba | 5311 Bb | 22844 B | 562 |
| Promedio estacional | 5648 c | 5522 c | 10696 a | 7974 b | 29841 | 259 |
| EEM† | 531 | 421 | 694 | 555 | 1003 | |
AB,abcdAverages with different uppercase letters in a column and different lower case letters in a row are statistically different (p≤0.05).
†Standard error of the mean. §Fixed cutting = fall: 35 d, winter: 42 d, and spring-summer 28 d between cuttings.
Accumulated forage yield of L. corniculatus in function of the percentages of LI were not different (p>0.05), unlike the findings of Da Silva and Nascimento, (2007), Da Silva and Hernández, (2010). Mattera et al. (2013) indicated that forage species should be harvested at 95 % intercepted light, but only when plant density is competitive. In this respect, Rojas et al. (2016) in alfalfa (M. sativa L.) recorded 95 % intercepted light only in summer; this was explained by the low plant density due to the time of establishment of the pasture. Mattera et al. (2013) also found that 95 % intercepted light in alfalfa is achieved in spring when combining distances between plants of 10, 15, 20, 25 and 30 cm, while in summer and winter only densities of 10 and 15 cm are present.
Botanical and morphological composition
The percentage of weeds in the pasture was low, 4 % annual average, in both years (Table 2). In this respect, Teixeira et al. (2007) mentioned that the presence of weeds in a pasture can increase over time as an effect of cutting frequency. Among the morphological components, leaves contributed most to yield with an annual average of 17 247 kg MS ha-1 (58 %), followed by stems (31%) and dead matter (7 %). This behavior can result from the accelerated rhythm of leaf and stem growth favored by environmental conditions such as photoperiod, temperature and humidity (Difante et al., 2009).
Table 2 Annual forage yield, by botanical and morphological component, of L. corniculatus, genotype 202700, in function of percentages of intercepted light (LI) and one fixed cutting defined seasonally.
| LI (%) | Hoja | Tallo | Material muerto | Maleza | EEM† |
| Año 1 (2014 - 2015) | |||||
| 90 | 16435 BCa | 9389 ABb | 3161 Ab | 1333 Ad | 477 |
| 95 | 18858 Aa | 10418 Ab | 1977 Ac | 1020 Ac | 691 |
| 100 | 17335 Aba | 10919 Ab | 3598 Ac | 1383 Ad | 749 |
| Corte fijo § | 14504 Ca | 8386 Bb | 2208 Ac | 2582 Ac | 1042 |
| Promedio | 16783 a | 9778 b | 2736 c | 1579 c | 430 |
| EEM† | 847 | 669 | 790 | 593 | |
| Año 2 (2015 - 2016) | |||||
| 90 | 18788 Aba | 10387 Ab | 1976 ABc | 132 Ad | 458 |
| 95 | 19622 Aa | 12091 Ab | 1178 Bc | 809 Ac | 1010 |
| 100 | 17674 Ba | 10816 Ab | 2833 Ac | 211 Ad | 477 |
| Corte fijo§ | 14763 Ca | 5794 Bb | 979 Bc | 1308 Ac | 574 |
| Promedio | 17712 a | 9772 b | 1741 c | 615 d | |
| EEM† | 470 | 762 | 379 | 724 | |
AB,abcdAverages with different upper case letters in a column and different lower case letter in a row are statistically different (p≤0.05). †EEM = standard error of the mean. §Fixed cutting= Fall: 35 d, winter: 42 d, and spring-summer: 28 d between cuttings.
In both years, the treatment with 95 % intercepted light produced the largest amount of leaf, 19,240 kg MS ha-1 on average, and the fixed cutting had the lowest amount of leaf and also of stem, 14,634 and 7,090 kg MS ha-1, respectively (p≤0.05), relative to the other treatments (Table 2). These results are similar to those of Giacomini et al. (2009), who evaluated B. brizantha cv. Marandu at intercepted light of 95 % and 100 % and two cutting intensities (10 and 15 cm); they found a larger quantity of leaf at 95 % intercepted light at a height of residual forage of 10 cm. In our study, dead forage was not different (p>0.05) among treatments during the first year. In the second year, the highest values were recorded with 100 % intercepted light and the lowest at 95 % intercepted light and fixed cutting. This indicates that the optimal harvest time is when intercepted radiation is at 95 %, which produced the best quality forage and, based on the lowest quantity of dead matter, the highest yield (Table 1, year 2) and the largest amount of leaf (Table 2), similar to the results reported by Montagner et al. (2012). Moreover, the minimal presence of dead or senescent matter in the fixed cutting could be because the plant did not reach its optimal physiological moment for harvesting and more light penetrated into the pasture (Matthew et al., 2000), causing higher production of green matter, leaves and stems but of lower weight (Villegas et al., 2004).
Depending on the season of the year, leaves were the component that most contributed to forage yield (Figure 2). The highest and the lowest seasonal yields were found in spring and fall-winter, respectively, with 5813 and 3378 kg MS ha-1. The treatment with 95 % intercepted light and fixed cutting had yields of 6480 and 5177 kg MS ha-1. These results indicate effects of season and of the treatments on production of leaves, influenced by the environmental conditions during the study (Figure 1). Moreover, the highest and the lowest quantities of stems were produced during spring and winter, with seasonal averages of 3814 and 1612 kg MS ha-1. In this respect, Sbrissia et al. (2010) mention that the production of stems changes in function of the seasonal variations in quantity and quality of light that reaches the pasture, precipitation and temperature. Regarding dead matter and weeds, there were no seasonal differences between the two years.

Plant height
There were differences among treatments applied and seasons of the year (p≤0.05), except in summer of the second year (p>0.05). The highest average annual heights were recorded in 100 and 95 % intercepted light with 23 cm in the first year and 24 cm in the second, while the lowest heights were in the fixed cutting with an average of 19 cm in both years. These results are related to plant age because the higher the percentage of intercepted light the more the days of plant regrowth (51 d, average of the two years1), relative to fixed cutting (33 d, both years4), which was always at a younger age with more frequent cuttings, affecting forage yield (Villegas et al., 2006). In this respect, Santos et al. (2004) pointed out that taller height means higher forage yield, but Scheffer et al. (2011) related height and yield of L. corniculatus to growth habit (erect and prostrated).
Table 3 L. corniculatus, genotype 202700, mean plant height in function of percentage of intercepted light (LI) and one fixed cutting defined seasonally.
| LI (%) | Otoño | Invierno | Primavera | Verano | Promedio anual | EEM† | |
| Año 1 (2014 - 2015) | |||||||
| 90 | 22 Ab | 17 Cc | 23 BCab | 25 Aa | 22 B | 0.7 | |
| 95 | 21 Ab | 19 Bb | 25 Ba | 25 Aa | 22 AB | 0.8 | |
| 100 | 16 Bc | 21 Ab | 28 Aa | 26 Aa | 23 A | 0.6 | |
| Corte fijo§ | 22 Aa | 15 Db | 22 Ca | 22 Ba | 20 C | 1.0 | |
| Promedio estacional | 20 b | 18 c | 24 a | 24 a | 22 | 22 | |
| EEM† | 0.6 | 0.3 | 0.7 | 0.6 | 0.2 | ||
| Año 2 (2015 - 2016) | |||||||
| 90 | 21 Aab | 16 Bb | 24 Ba | 23 Aa | 21 B | 1.5 | |
| 95 | 19 ABc | 23 Ab | 30 Aa | 23 Ab | 24 A | 0.7 | |
| 100 | 18 BCc | 23 Ab | 28 ABa | 22 Ab | 23 AB | 0.8 | |
| Corte fijo§ | 17 Cb | 13 Bc | 19 Cb | 23 Aa | 18 C | 1.1 | |
| Promedio estacional | 19 c | 19 c | 25 a | 23 b | 21 | 21 | |
| EEM† | 0.7 | 1.6 | 1.4 | 0.8 | 0.7 | ||
AB,abcdAverages with different upper case letters in a column and different lower case letter in a row are statistically different (p≤0.05). EEM† = standard error of the mean. §Fixed cutting= Fall: 35 d, winter: 42 d, and spring-summer: 28 d between cuttings.
In the first year, the tallest heights were found in spring and summer with 24 cm and the shortest in winter with 18 cm, while in the second year, the tallest heights were found in spring with 25 cm and in fall and winter the shortest, with 19 cm. The tallest plants were found in the seasons with the conditions of humidity and temperature that were optimum for growth and yield of the species (Figure 1). These results are similar to those of García et al. (2014), who evaluated 12 genotypes of L. curniculatus. Also, according to Marcelino et al. (2006), with adequate conditions of photoperiod, temperature and humidity, forage plants have accelerated growth and change in height in different seasons (Difante et al., 2009), correlating positively with forage yield (Da Silva and Nacimiento Jr., 2007), as occurred in our study.
Plant height can be associated with forage production (Hakl et al., 2010). Moreover, production of L. corniculatus forage is related to height, but also to the growth habit of the genotype, with the lowest yields and heights in prostrated genotypes (García et al., 2014). Height of the pasture gives an idea of the forage produced. Therefore, when calculating the equivalence of the highest average seasonal yields and of the treatments, each cm of height corresponds to 178 kg MS ha-1 for spring and 184 kg MS h-1 for 95 % LI, averages of both years.
Leaf: stem ratio
The highest annual average leaf: stem ratio occurred with fixed cutting (Table 4), followed by 90, 95 and 100 % intercepted light in both years. Fixed cutting surpassed the average of the treatments with 95 and 100 % LI by 26 % (2.3 vs 1.7: 1) in the first year, and in the second year by 39 % at 100 % LI (2.8 vs 1.7:1). In the fixed cutting treatment, the highest leaf: stem ratio was the result of more frequent cuttings (33 d yearly average5), which did not permit harvesting the species at its optimal moment, but rather in the phase of accelerated growth when the largest percentage of young leaves and less stem is found. Consequently, there was more light penetrating into the interior of the pasture (Barbosa et al., 2011). Also, Montagner et al. (2012) mention that, after cutting, the quality and quantity of incidental light inside the pasture is altered by the cutting interval, which causes variations in the production of leaves and stems and, therefore, in the leaf: stem ratio.
Table 4 Leaf:stem ratio of L. corniculatus, genotipo 202700, in function of the percentage of intercepted light and one fixed cutting defined seasonally.
| LI (%) | Otoño | Invierno | Primavera | Verano | Promedio anual | EEM† | |
| Año 1 (2014 - 2015) | |||||||
| 90 | 2.0 Aa | 2.2 Aa | 1.8 Aa | 2.0 Aa | 2.0 AB | 0.30 | |
| 95 | 1.9 Aab | 2.0 Aa | 1.6 Abc | 1.4 Ac | 1.7 B | 0.14 | |
| 100 | 1.9 Aa | 1.9 Aa | 1.5 Ab | 1.5 Ab | 1.7 B | 0.32 | |
| Corte fijo§ | 2.7 Aa | 2.4 Aa | 2.0 Aa | 2.1 Aa | 2.3 A | 0.47 | |
| Promedio estacional | 2.1 a | 2.1 a | 1.7 b | 1.8 b | 1.9 | 0.09 | |
| EEM† | 0.28 | 0.52 | 0.26 | 0.37 | 0.20 | ||
| Año 2 (2015 - 2016) | |||||||
| 90 | 2.6 Aba | 2.9 Aa | 1.5 Bb | 1.8 Bb | 2.2 B | 0.28 | |
| 95 | 2.4 Ba | 2.3 ABa | 1.3 Bb | 1.7 Bb | 2.0 BC | 0.23 | |
| 100 | 2.1 Ba | 1.7 Bab | 1.2 Bb | 1.6 Bab | 1.7 C | 0.24 | |
| Corte fijo§ | 3.3 Aa | 2.7 Abc | 2.2 Ac | 2.9 Aab | 2.8 A | 0.20 | |
| Promedio estacional | 2.6 a | 2.4 a | 1.6 c | 2.0 b | 2.1 | 0.12 | |
| EEM† | 0.27 | 0.25 | 0.15 | 0.20 | 0.11 | ||
AB,abcdAverages with different upper case letters in a column and different lower case letter in a row are statistically different (p≤0.05). †EEM = standard error of the mean. §Fixed cutting= Fall: 35 d, winter: 42 d, and spring-summer: 28 d between cuttings.
The leaf: stem ratio in both years was different (p≤0.05; Table 4) between seasons. It was higher in fall and winter, with 2.1:1 and 2.5:1 for the first and second years. The lowest values of the leaf: stem ratio were recorded in spring, in both years, and were different to summer only in the second year (p≤0.05). These results are similar to those reported by Barbosa et al. (2011), who found a higher leaf: stem ratio in seasons when plant growth is slower (fall and winter), due to a higher density, but lower weight, of stems. Moreover, a lower leaf: stem ratio in spring and summer results from heavier individual stems (Villegas et al., 2004). In this respect, Giacomini et al. (2009) mention that translocation of assimilates, from leaves to stems, is greater in spring and summer, resulting in heavier stems. The proportion of leaf relative to stem was higher in all seasons and treatments, with an average of 1.9:1 (first year) and 2.1:1 (second year).
Conclusions
The best productive performance of the Lotus corniculatus genotype 202700 was obtained with the percentages of intercepted light, marking differences with fixed cutting. Nevertheless, with 95 % LI, the largest quantity of leaves, mainly in spring, was produced, and thus we can define this as the best management strategy. However, with fixed cutting, the species yielded less. Forage production in our study is different from that observed in other parts of the world and it should be investigated under the different types of environments in Mexico.
REFERENCES
Barbosa, R. A., D. Nascimento Jr., H. H. Vilela, S. C. Da Silva, V. P. Batista-Euclides, A. F. Sbrissia, and B. S. Da Lana. 2011. Morphogenic and structural characteristics of guinea grass pastures submitted to three frequencies and two defoliation severities. Rev. Bras. Zootec. 40: 947-954. [ Links ]
Beuselinck, P. R., and F. W. Grant. 1995. Birsdfoot trefoil. In: Barnes R. F., H. Brown A., and J. Crowford D. Forages, an Introduction to Grassland Agriculture. Iowa State University. pp: 237-248. [ Links ]
Castro B., M. R. 2006. Elaboración de Material Curricular para la Práctica Básica de Estudiantes de Medicina Veterinaria de la Universidad Austral de Chile. Facultad de Ciencias Veterinarias. Valdivia-Chile. 144 p. [ Links ]
Da Silva S., C., and D. Nascimento Jr. 2007. Avanços na pesquisa com plantas forrageiras tropicais em pastagens: características morfofisiológicas e manejo do pastejo. Rev. Bras. Zootec. 36: 121-138. [ Links ]
Da Silva S., C. , y A. Hernández G. 2010. Manejo de Pastoreo en Praderas Tropicales. Forrajes y su Impacto en el Trópico. 1ra Edición. México. Universidad Autónoma de Chiapas (UNACH). pp: 43-62. [ Links ]
Díaz L., J. E., J. García A., y M. Rebuffo. 1996. Crecimiento de leguminosas en la Estanzuela. I.N.I.A (ed). Serie Técnica 71: 12. [ Links ]
Difante G., S., D. Nascimento Jr., C. Da Silva S., V. P. Bautista E., A. de Moura Z., and B. Adese. 2008. Dinâmica do perfilhamento do capim-marandu cultivado em duas alturas e três intervalos de corte. Rev. Bras. Zootec. 37: 89-196. [ Links ]
Difante G., S. , D. Nascimento Jr., V. P. Batista E., S. C. Da Silva., R. Amorin B., and W. Velásquez G. 2009. Sward structure and nutritive value of tanzania guineagrass subjected to rotational stocking managements. Rev. Bras. Zootec. 38: 9-19. [ Links ]
García B., DV., J de D. Guerrero R., G. García de los S., y S. A. Lagunes R. 2014. Rendimiento y calidad de forraje de genotipos de Lotus corniculatus L., en el Estado de México. Nova Scientia. 7: 170-189. [ Links ]
García E. 2004. Modificación al Sistema de Clasificación Climática de Köppen. 4ª. Edición. Instituto de Geografía. Universidad Nacional Autónoma de México. México. 217 p. [ Links ]
Giacomini, A. A., S. C. Da Silva, D. O. de L. Sarmento, C. V. Zeferino, S. J. Souza Jr., J. K. Da Trindade, V. Del A. Guarda, and D. Nacimiento Jr. 2009. Growth of marandu palisadegrass subjected to strategies of intermittent stocking. Sci. Agric. (Piracicaba, Braz.). 66: 733-741. [ Links ]
Grant, F. W. 2009. Lotus corniculatus. Sci. Topics. Canada. http://www.scitopics.com/Lotus_corniculatus.html (Consulta: septiembre 2016). [ Links ]
Gunn, C. R., H. J. Wiersema, A. C. Ritchie, and H. J. Kirkbride. 1992. Families and general of spermatophytes recognized by the Agricultural Research Service. Tech. Bull. 1796: 476 p. [ Links ]
Lagler J., C. 2003. Lotus: Un género que no acaba en dos especies. Rev. Forrajes y Granos. Buenos Aires, Argentina. pp: 72-76. [ Links ]
Lemaire, G., and M. Agnusdei. 2000. Leaf tissue turn-over and efficiency of herbage utilization. In: Lemaire G., J. Hodgson, A. de Morales, C. Nabinger, and C. de F Carvalho P. Grassland Ecophysiology and Grazing Ecology. Wallingford: CAB International. pp: 265-288. [ Links ]
Marcelino K., R. A., D. Nascimento Jr., S. Da Silva C., V. P. Euclides B., and D. Da Fonseca M. 2006. Características morfogênicas e estruturais e produção de forragem do capim-marandu submetido a intensidades e frequências de desfolhação. Rev. Bras. Zootec. 35: 2243-2252. [ Links ]
Marley, C., L., R. Fychan, and R. Jones. 2006. Yield, persistency and chemical composition of Lotus species and varieties (birdsfoot trefoil and greater birdsfoot trefoil) when harvested for silage in the UK. Grass Forage Sci. 61: 134-145. [ Links ]
Mattera, J., L. A. Romero, A. L. Cuatrín, P. S. Cornaglia, and A. A. Grimoldi. 2013. Yield components, light interception and radiation use efficiency of lucerne (Medicago sativa L.) in response to row spacing. Europ. J. Agron. 45: 87-95. [ Links ]
Matthew, C., G. S. Assuero, K. C. Black, and H. N. R. Sackville. 2000. Tiller dynamics of grazed swards. In: Lemaire G. , J. Hodgson , A. de Morales C., Nabinger C., de F Carvalho P. Grassland Ecophysiology and Grazing Ecology. Wallingford: CABI Publishing. pp: 127-150. [ Links ]
Montagner, D., B., D. Nascimento Jr., H. H. Vilela, B. M. Sousa, de L. V. P. B. Euclides, S. C. Da Silva, and M. N. Carloto. 2012. Tillering dynamics in pastures of guinea grass subjected to grazing severities under intermittent stocking. Rev. Bras. Zootec. 41: 544-549. [ Links ]
Nabinger C., and S. Pontes L. 2001. Morfogênese de plantas forrageiras e estrutura do pasto. In: Reunião Anual da Socidade Brasileira de Zootecnia, Piracicaba. Anais. Piracicaba: SBZ. 38: 755-771. [ Links ]
Rojas G., A. R., A. Hernández G., S. Joaquín C., M. de los A. Maldonado P., S. I. Mendoza P., P. Álvarez V., y B. M. Joaquín T. 2016. Comportamiento productivo de cinco variedades de alfalfa. Rev. Mex. Cienc. Agríc. 7: 1855-1866. [ Links ]
Rojas G., A. R., A. Hernández G., S. Joaquín, C., S. I. Mendoza, P., J. D. Guerrero R., y J. L. Zaragoza, R. 2012. Comportamiento productivo y rendimiento de forraje de cinco variedades de alfalfa. 2da Reunión Internacional conjunta de manejo de pastizales y producción animal. Zacatecas, México. pp: 336-340. [ Links ]
Santiñaque F., H., y P. De Battista J. 2003. Niveles de agua y frecuencias de defoliación en el comportamiento de Lotus corniculatus L. Agrociencia. 7: 41-51. [ Links ]
Santos P., M., A. Balsalobre M., and M. Corsi. 2004. Características morfogenéticas e taxa de acúmulo de forragem do capim Mombaça submetido a três intervalos de corte. Rev. Bras. Zootec. 33: 843-851. [ Links ]
SAS, Institute. 2009. SAS/STAT® 9.2. User´s Guide. Second Edition. Cary, NC: SAS Institute Inc. USA. [ Links ]
Sbrissia, A., F., S. C. Da Silva, O. L. D. Sarmento, L. K. E. F. Molan, M. Andrade, A. C. Goncalves, and A. V. Lupinacci. 2010. Tillering dynamics in palisadegrass swards continuously stocked by cattle. Plant Eco. 206: 349-359. [ Links ]
Scheffer, B. M. S., R. Brustolin, and M. A. Dall. 2011. Performance of Lotus corniculatus L. genotypes submitted to cutting interval: subsidies to a breeding program. Rev. Bras. Zootec. 40: 1645-1650. [ Links ]
Teixeira, E. I., M. J. Derrick, B. E. Hamish, and F. L. Andrew. 2007. The dynamics of lucerne (Medicago sativa L.) yield components in response to defoliation frequency. Eur. J. Agron. 26: 394-400. [ Links ]
Villegas A., Y., A. Hernández-Garay, J. Pérez P., C. López C., J. G. Herrera H., J. F. Enríquez Q., y A. Gómez V. 2004. Patrones estacionales de crecimiento de dos variedades de alfalfa (Medicago sativa L.). Téc. Pecu. Méx. 42: 145-158. [ Links ]
Villegas A., Y. , A. Hernández-Garay, PA Martínez H., J. Pérez P., JG Herrera H., C. López C. 2006. Rendimiento de forraje de variedades de alfalfa en dos calendarios de corte. Rev. Fitotec. Méx. 29: 369-372. [ Links ]
Received: July 2017; Accepted: January 2018
 Este es un artículo publicado en acceso abierto bajo una licencia Creative Commons
Este es un artículo publicado en acceso abierto bajo una licencia Creative Commons
