











 text in
text in  English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO 
 Permalink
PermalinkIntroducción
A través del año los árboles frutales presentan diversas fases del desarrollo o fenológicas, tales como flujos de crecimiento vegetativo o de raíces, y de desarrollo floral y del fruto (Wolstenholme y Whiley, 1989). Los ritmos de crecimiento anual varían con el tipo de clima y las condiciones de cultivo, por lo cual se han elaborado diagramas fenológicos para el aguacate (Persea americana Miller) ‘Hass’ con diferente grado de complejidad en regiones productoras como Australia (Whiley et al., 1988), Sudáfrica (Whiley y Wolstenholme, 1990), Chile (Gardiazabal-Irazábal y Rosenberg, 1991), EUA (Arpaia et al., 1994-95), y Nueva Zelanda (Thorp et al., 1995). En México, la fenología de ‘Hass’ está descrita para las regiones productoras de Nayarit (Cossio-Vargas et al., 2008) y Michoacán (Rocha-Arroyo et al., 2011a). El aguacate ‘Méndez No. 1’ (Plant Patent 11,173 USA 2002), conocido en México como ‘Méndez’, tiene importancia económica en el sur del estado de Jalisco, donde hay más de 6000 ha plantadas. Su relevancia se debe a que una proporción importante de su cosecha anual madura más temprano (verano) que la de ‘Hass’ (otoño-invierno) y esa cosecha alcanza los precios más altos del año.
Por desconocimiento de los aspectos hortícolas de ‘Méndez’, el manejo de los huertos se hace similar a ‘Hass’. Pero, dada la importancia creciente de ‘Méndez’ es necesario desarrollar tecnología de producción apropiada para este cultivar de aguacate. Por lo tanto, el objetivo de esta investigación fue documentar el comportamiento fenológico del aguacate ‘Méndez’ en el sur del estado de Jalisco.
Materiales y Métodos
La investigación se realizó durante los ciclos de producción 2014-15 y 2015-16 en dos huertos comerciales de aguacate ‘Méndez’ de la empresa Agro González, S.P.R. de R.L., con fertirriego, suelo Feozem háplico y clima Semicálido subhúmedo [AC(w)] (García-Amaro, 1998) del sur del estado de Jalisco. El huerto Colorín está en Atequizayan, Municipio de Zapotlán el Grande y el huerto Ocote Cuate 2 está en Zapotiltic, Municipio de Zapotiltic. Para estos huertos, la altitud es 1556 m y 1428 m, el distanciamiento entre árboles 7×3.5 m y 5×5 m y la edad al inicio del estudio de 4 y 6 años, respectivamente.
Características del suelo
En cada huerto se seleccionaron al azar cinco árboles y se obtuvo una muestra de suelo integrada por cuatro submuestras de la zona de goteo del árbol, a 0-30 cm de profundidad, que es la zona con mayor cantidad de raíces jóvenes (Salazar-García y Cortés-Flores, 1986). Los análisis se realizaron en un laboratorio acreditado por The North American Proficiency Testing (NAPT) Program (programa de Soil Science Society of America). Las variables fueron textura, pH (1:2 agua) (McLean, 1982), materia orgánica (MO) por el método de Walkley y Black (Nelson y Sommers, 1982), N-inorgánico (Dahnke, 1990), P-Bray (Bray y Kurtz, 1945), K, Ca, Mg y Na con acetato de amonio (Doll y Lucas, 1973), Fe, Zn, Cu, Mn por el método DTPA (Lindsay y Norvell, 1978) y B se determinó por el método de agua caliente y Azometina-H (Bingham, 1982). Las variables P y B se determinaron en un espectrofotómetro Genesys 20 (Thermo Scientific, Madison, USA); para los otros nutrientes se usó un espectrofotómetro de absorción atómica (Thermo Series S, Shangai, China).
Variables fenológicas
En 10 árboles por huerto se marcaron 20 brotes por árbol, al inicio de los flujos vegetativos de invierno y verano 2014 y 2015. De cada árbol y brote marcado se recolectó una yema apical o brote floral a intervalos quincenales o semanales según se aproximaba la antesis. Las yemas y brotes florales se fijaron en FAA (formaldehído:ácido acético:etanol, 5:5:90, v:v:v) y después se introdujeron a una campana de vacío (Nalgene 8040317, Nalgen Company) a 30 KPa por 5 h. Luego se clasificaron con un microscopio estereoscópico (Zeiss Stereomikroskop Mod. Stemi 2000-C, Carl Zeiss, Göttingen, Germany), con la escala visual de Salazar-García et al. (1998) que comprende desde E-1 (yema vegetativa) a E-11 (antesis).
En cada huerto se seleccionó otro grupo de 10 árboles, con historial de producción anual de al menos 50 kg por árbol y fueron usados en todas las evaluaciones, excepto las del desarrollo floral. En febrero y agosto de 2014 y 2015 en cada árbol se etiquetaron 30 brotes al inicio de cada flujo vegetativo (invierno y verano, respectivamente). El tipo de crecimiento producido (brotes florales, vegetativos o inactivos) por los brotes marcados se cuantificó al término de cada floración, en octubre y febrero 2014, así como en noviembre 2015 y marzo 2016.
Los muestreos de raíces se hicieron cada mes de febrero 2014 a febrero 2016 alternando árboles nones y pares, así como las orientaciones norte y sur de los árboles. Las raíces jóvenes, de color café claro, en un volumen de suelo de 40 ( 40 ( 40 cm en la zona de goteo del árbol, se extrajeron, lavaron y deshidrataron en un horno con aire forzado (Binder Mod. ED 240, Binder Corp., Tutllingen, Alemania) a 70 °C por 72 h para obtener su peso seco.
La caída de fruto se cuantificó cada año para frutos de las floraciones de verano e invierno. En antesis se marcaron cinco brotes florales en cada punto cardinal de la parte media de la copa de cada 10 árboles seleccionado. Para frutos de la floración de verano se contabilizó el número de frutos por brote floral dos (noviembre) y seis (marzo) meses después de antesis. En frutos de la floración de invierno esto se hizo dos (abril) y cinco (julio) meses después de antesis.
En cada árbol marcado se etiquetaron cinco frutos de cada floración cuando tenían 2 cm de longitud. Cada mes se midió su longitud y diámetro con un vernier digital (Mitutoyo, Mod. CD-6’’ CSX, Mitutoyo Corp., Kawasaki, Japón) desde octubre (frutos de la floración de verano) y abril (frutos de la floración de invierno).
Variables climáticas
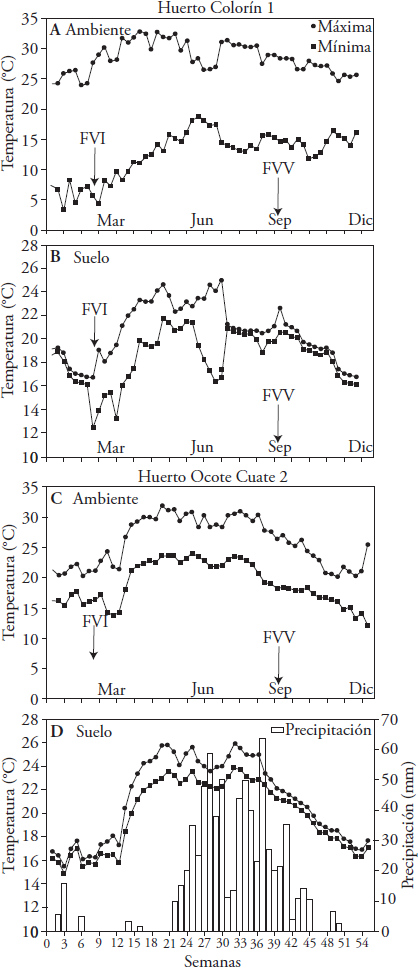
Los datos de precipitación pluvial fueron obtenidos del Servicio Meteorológico Nacional (SMN, 2016). Las temperaturas ambiental y del suelo (a 30 cm de profundidad) fueron registradas en cada huerto con termómetros automatizados (HOBO H8, Onset Computer, Witzprod, Englewood Cliffs, NJ, USA) operados a batería.
Análisis de la información
El diseño experimental fue de bloques al azar con un arreglo factorial 2 ( 2, huertos
(Colorín 1 y Ocote Cuate 2), flujos vegetativos (verano e invierno), con 10
repeticiones (árboles) y la unidad experimental fue 30 brotes por árbol. Los
ANDEVA se realizaron con medidas repetidas para cada huerto y flujo vegetativo.
Previo a su análisis, los valores expresados como porcentaje fueron
transformados mediante el arcoseno
Resultados y Discusión
Fertilidad del suelo
Las características de fertilidad del suelo fueron similares entre huertos. La MO, Ca y Mg fueron de moderadamente bajos a bajos y en el huerto Colorín 1 fueron notorios los niveles bajos de Mn, B y Zn (Cuadro 1). El contenido bajo de MO además de los niveles insuficientes de Mn, B y Zn son característicos de los suelos de origen volcánico (Salazar-García et al., 2016), lo que sugiere la necesidad de incorporar MO y los nutrientes en concentraciones moderadamente bajas a bajas para mejorar la fertilidad del suelo. En ambos huertos el pH fue neutro (Cuadro 1). En otras regiones productoras de aguacate los suelos presentan pH similares o más ácidos (Aguilera-Montañez y Salazar-García, 1991; Salazar-García y Lazcano-Ferrat, 1999). Sin embargo, el aguacate puede prosperar en pH de 4.8 o superiores a 6.5 (Salazar-García, 2002).
Cuadro 1 Características del suelo de los huertos a 0-30 cm de profundidad al inicio del estudio (febrero 2014).
| Características | Huerto Colorín 1 | Huerto Ocote Cuate 2 |
| pH (1:2 H2O) | 7.1 (Neutro) | 6.6 (Neutro) |
| Materia orgánica (%) | 0.82 (B) | 1.7 (MoB) |
| Conductividad eléctrica (dS/m) | 0.26 | 0.33 |
| mg kg-1 | ||
| N-NO3 | 5.6 (MB) | 6.4 (MB) |
| P-Bray | 22.8 (M) | 66.4 (MA) |
| K | 180 MoB) | 491 (MoA) |
| Ca | 1273 (MoB) | 859 (MoB) |
| Mg | 96.3 (B) | 183 (MoB) |
| Na | 74 (M) | 49.1 (B) |
| Fe | 17.6 (MoA) | 79.4 (MoA) |
| Cu | 2.3 (A) | 4.0 (A) |
| Mn | 1.6 (MB) | 3.9 (B) |
| Zn | 0.54 (B) | 1.2 (MoA) |
| B | 0.22 (MB) | 0.32 (MB) |
Interpretación de laboratorio: A: alto; MoA: moderadamente alto; MA: muy alto; M: medio; B: bajo; MoB: moderadamente bajo; MB: muy bajo; N: normal.
Tipo de crecimiento
La producción de brotes florales tuvo un comportamiento diferente entre huertos. En el Colorín 1 se registró 33.1 % y en el Ocote Cuate 2 19.1 % (Cuadros 2 y 3). El descenso en la temperatura ambiental es el factor que promueve el desarrollo floral en aguacate (Salazar-García et al., 1999, 2006a, 2013). La diferencia en la producción de brotes florales entre los huertos se explica por las temperaturas más bajas en el Colorín 1 (promedio anual 20.5 °C) respecto al Ocote Cuate 2 (22.5 °C) (Figuras 1a y 1b) lo que favoreció mayor exposición del huerto El Colorín 1 a temperaturas más frescas. El flujo vegetativo de origen (invierno o verano) no afectó el tipo de crecimiento producido (vegetativo, floral o inactivo) (Cuadros 2 y 3).
Cuadro 2 Análisis de varianza e interacciones huerto x flujo del tipo de crecimiento producido por brotes de los flujos vegetativos de invierno y verano del aguacate ‘Méndez’. Datos de dos años de los huertos Colorín 1 y Ocote Cuate 2.
| Tipo de crecimiento | Fuente de variación† | GL | SC | CM | F | P > F |
| Brotes vegetativos | Huerto | 1 | 0.01413000 | 0.01413000 | 0.00 | 0.9525 |
| Flujo | 1 | 3.27166727 | 3.27166727 | 0.83 | 0.3658 | |
| Huerto*Flujo | 1 | 0.42702000 | 0.42702000 | 0.11 | 0.7434 | |
| Brotes florales | Huerto | 1 | 47.17805953 | 47.17805953 | 6.58 | 0.0119 |
| Flujo | 1 | 3.01030617 | 3.01030617 | 0.42 | 0.5185 | |
| Huerto*Flujo | 1 | 14.57155268 | 14.57155268 | 2.03 | 0.1572 | |
| Inactivos | Huerto | 1 | 76.71389754 | 76.71389754 | 14.12 | 0.0003 |
| Flujo | 1 | 3.53008144 | 3.53008144 | 0.65 | 0.4223 | |
| Huerto*Flujo | 1 | 23.07924822 | 23.07924822 | 4.25 | 0.0421 |
†Análisis de varianza realizado con datos transformados mediante el arcoseno
Cuadro 3 Efecto del huerto y flujo vegetativo de origen sobre el tipo de crecimiento producido en aguacate ‘Méndez’. Evaluaciones realizadas en: octubre 2014 y noviembre 2015 (brotes de invierno) y febrero 2015 y marzo 2016 (brotes de verano).
| Fuente de variación | N† | Tipo de crecimiento del total de brotes marcados (%) | ||
| Vegetativo | Floral | Inactivo | ||
| Huerto Colorín 1 | 774 | 53.8 | 33.1 a¶ | 13.0 b |
| Huerto Ocote Cuate 2 | 775 | 53.8 | 19.3 b | 26.7 a |
| Pr > F | 0.9525 | 0.0119 | 0.0003 | |
| Brotes de invierno | 917 | 52.2 | 26.3 | 21.1 |
| Brotes de verano | 634 | 55.8 | 26.3 | 17.8 |
| Pr > F | 0.3658 | 0.5185 | 0.4223 | |
†Número de brotes evaluados.
¶ Medias con distinta letra en una columna para cada huerto o tipo de brote son estadísticamente diferentes (Tukey; p≤0.05).
Crecimiento vegetativo
El registro fenológico comenzó con la brotación (febrero 2014) de brotes del flujo vegetativo de invierno. Del 100 % de brotes de invierno marcados, 25 % produjeron floración en el verano (septiembre 2014) y 6 % florecieron en el invierno siguiente (febrero 2015). El 55 % de los brotes de invierno produjo brotes de verano en septiembre 2014 y de éstos, 27 % florecieron en invierno (febrero 2015). Los brotes de invierno y verano produjeron una cantidad importante de brotes vegetativos (entre 50 y 60 %) (Figura 2). A diferencia de ‘Méndez’, en ‘Hass’ el flujo de invierno no produce floración en el verano; sin embargo, puede ocurrir si las condiciones ambientales son favorables (Rocha-Arroyo et al., 2011a).
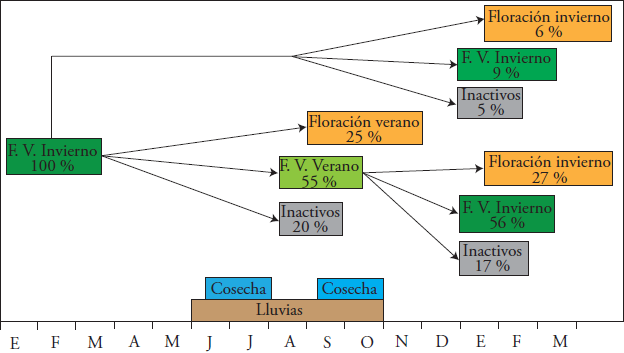
En ‘Hass’ el flujo vegetativo de invierno (conocido como flujo de primavera en otras regiones) es el más importante para la floración de invierno-primavera del siguiente año, y está documentado en México (Cossio-Vargas et al., 2008; Rocha-Arroyo et al., 2011b) y otros países (Mena-Volker, 2004; Robinson et al., 2002; Salazar-García y Lovatt, 1998; Thorp et al., 1995; Whiley et al., 1990).
Desarrollo floral
La duración del desarrollo floral (de yema vegetativa a antesis) mostró ligera variación entre años. Para el 2014 transcurrieron 218 y 180 d para los brotes de invierno y verano, respectivamente. La 1ª floración, o de verano, presentó antesis en septiembre y la 2ª floración, o de invierno, en febrero (Figura 3). En el 2015, los brotes de invierno completaron el desarrollo floral en 225 d produciendo la 1ª floración entre la última semana de septiembre y la primera de octubre. En el caso de brotes de verano el desarrollo floral requirió 193 d, con antesis en la primera semana de marzo 2016 (2ª floración).
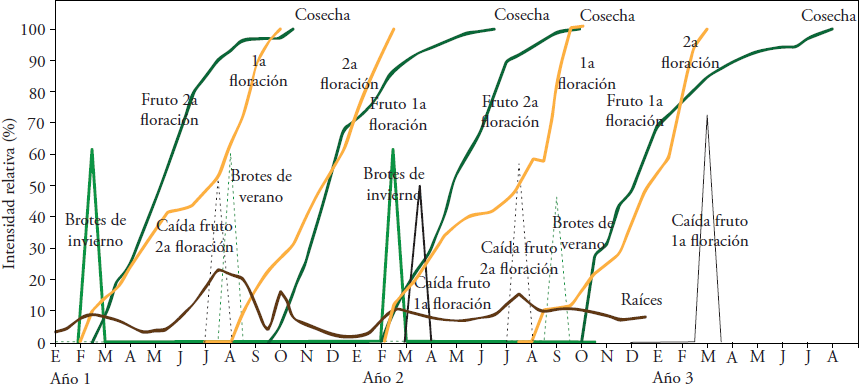
Figura 3 Principales etapas fenológicas del aguacate ‘Méndez’ en el sur del estado de Jalisco. 1ª floración: floración de verano; 2ª floración; floración de invierno.
En los dos años de estudio, los brotes de verano requirieron, en promedio, 35 d menos que los de invierno para completar el desarrollo floral. Esta diferencia fue menor que los 120 y 180 d mencionados para ambos flujos de ‘Hass’ en Nayarit y Michoacán, respectivamente (Cossio-Vargas et al., 2008; Rocha-Arroyo et al., 2011a). El menor tiempo requerido para que los brotes de verano de ‘Méndez’ completaran su desarrollo floral pudo deberse a que cuando emergieron (agosto-septiembre) la temperatura ambiental ya había iniciado su descenso (21 °C, en promedio), estimulando el desarrollo floral. El efecto del descenso de la temperatura en brotes de ‘Hass’ en desarrollo fue mencionado por Rocha-Arroyo et al. (2010) y Salazar-García et al. (2013).
Caída de fruto
Los frutos caídos en la denominada “segunda caída”, “caída fisiológica” o “caída de junio” (hemisferio norte) presentaron diámetros entre 5 y 6 cm. Para frutos provenientes de la primera floración (verano) la caída fue 61.8 % y ocurrió en marzo. La caída de fruto de la segunda floración (invierno) fue en julio (55.4 %). La magnitud de la caída fisiológica registrada en nuestro estudio es similar a la descrita para ‘Hass’ en otras regiones productoras: 45 % en Nayarit, México (Cossio-Vargas et al., 2008) y 54 % en Sudáfrica (Toerien, 1979). La edad de los frutos caídos procedentes de las floraciones de verano e invierno fue de 22 y 26 semanas después de antesis, respectivamente. La caída de fruto de la segunda floración fue poco antes del flujo vegetativo de verano y para los de la primera floración fue después del flujo vegetativo de invierno (Figura 3).
Cossio-Vargas et al. (2008) y Lovatt (1990) mencionan la coincidencia y Wolstenholme y Whiley (1989) la no coincidencia entre la caída de fruto y la presencia de crecimiento vegetativo, pero no hay información concluyente sobre las causas que originan la segunda caída de fruto (Dixon et al., 2006). Una hipótesis menciona el desbalance hormonal como responsable de esta caída de fruto, y destaca el ácido abscísico (ABA) como principal causante (Adato y Gazit, 1977). Para disminuir la intensidad de esta caída en aguacate ‘Hass’ se evaluaron aspersiones foliares con 2,4-D o aminoethoxyvinylglycina desde dos meses antes del inicio de la caída de fruto, y en el segundo año de estudio disminuyó 18 y 17.7 %, respectivamente; además aumentó 32 y 23.6 % la producción de fruto de calibres grandes (170 a ≥ 266 g fruto-1), respectivamente, respecto al testigo (Salazar-García et al., 2006b). Garner y Lovatt (2016) confirmaron que el ABA está involucrado en la segunda caída de fruto en ‘Hass’ en California.
Crecimiento de fruto
Los frutos de ‘Méndez’ resultantes de las floraciones de verano e invierno requirieron 10 y ocho meses, respectivamente, para alcanzar la madurez fisiológica (22.7 % m.s.) (Figura 3). Para frutos de la floración de invierno hay resultados similares para ‘Hass’ (21.5 % m.s.) en Nayarit y Michoacán (Cossio-Vargas et al., 2008; Rocha-Arroyo et al., 2011a). Los frutos de la floración de verano alcanzaron la madurez fisiológica en julio, con un diámetro promedio de 6.5 cm y los de la floración de invierno entre septiembre y octubre con un diámetro promedio de 6.4 cm. Los frutos derivados de la floración de verano requirieran dos meses más para alcanzar la madurez fisiológica, lo cual se puede atribuir a que cuando éstos iniciaron su crecimiento (septiembre-octubre) las temperaturas del aire (21.8 °C) y suelo (20.8 °C) habían iniciado su descenso, lo cual disminuye el ritmo del metabolismo del árbol.
Producción de raíces
La producción de raíces no cesó durante el año y hubo dos periodos de mayor intensidad (Figura 3). El flujo más importante fue en los meses lluviosos del verano (junio-agosto), previo al flujo vegetativo de verano. El segundo flujo de crecimiento de raíces tuvo menor intensidad, fue de febrero a marzo y coincidió con la floración de invierno y su correspondiente flujo vegetativo. La mayor producción de raíces en el verano coincidió con lo descrito para ‘Hass’ cultivado sin riego en Nayarit (Cossio-Vargas et al., 2008) y con riego o sin él para el mismo cultivar en Michoacán (Bárcenas-Ortega et al., 2007). En Israel, Kalmar y Lahav (1976) encontraron en ‘Ettinger’ y ‘Fuerte’ una asociación positiva entre la cantidad de agua aplicada y la producción de raíces, pero en Michoacán (Bárcenas-Ortega et al. (2007) documentaron lo contrario en ‘Hass’.
Según Bernstein et al. (2004) en aguacate el crecimiento de las raíces puede ser más restringido por la salinidad que por la presencia de brotes vegetativos. En nuestro estudio, en ambos huertos la CE del suelo varió de 0.26 a 0.33 dS m-1 por lo que no fue un factor limitante para el desarrollo de raíces y no afectó la caída de fruto. Además, la menor producción de raíces ocurrió cuando el fruto estaba en su máximo crecimiento (Figura 3).
Precipitación pluvial y temperaturas del aire y suelo
En ambos huertos la lluvia anual fue 728 mm, concentrada entre junio y septiembre (Figura 1). En el huerto Colorín 1 las temperaturas ambientales máximas y mínimas promedio de los meses más calientes (abril y mayo) y fríos (enero y febrero), fueron 32, 31 y 6.4 y 6.0 °C, respectivamente (Figura 1A). En el huerto Ocote Cuate 2 las temperaturas de los meses más calientes (mayo y junio) y frío (marzo) fueron 30.8 y 30.6 y 15.0, respectivamente (Figura 1C).
La temperatura del suelo más alta (23.8 °C) fue en mayo y la más baja (14.5 °C) en marzo en el Colorín 1 (Figura 1B). Para el Ocote Cuate 2 la temperatura del suelo máxima fue en mayo (25.2 °C) y las mínimas en enero (15.9 °C) y febrero (16.0 °C) (Figura 1D). En ambos huertos, las temperaturas máximas del suelo se mantuvieron entre 21 a 27 °C, mencionadas por Yusof et al. (1969) como adecuadas para el crecimiento de las raíces de aguacate. De igual manera, la temperatura mínima fue superior a 13 °C, señalada por Whiley et al. (1990) como el límite inferior a partir del cual el crecimiento de las raíces de aguacate se detiene.
Conclusiones
El aguacate ‘Méndez’ presentó dos flujos de crecimiento vegetativo: verano e invierno. Los brotes de verano produjeron la floración de invierno y los de invierno la de verano. El desarrollo floral se completó en 186 y 221 d para brotes de verano e invierno, respectivamente. La máxima producción de raíces fue en la época de lluvias y la menor durante el máximo crecimiento del fruto. La caída de fruto fue ligeramente superior para la 1ª floración (verano). Los frutos de la floración de verano requirieron dos meses más que los de invierno para lograr su madurez fisiológica. La presencia permanente de brotes vegetativos desocupados (sin fruto) junto con el descenso de la temperatura en la primavera es clave para la floración de verano cuyo fruto es de alto valor comercial.

