










Services on Demand
Journal
Article
 text in
text in  English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkAgrociencia
On-line version ISSN 2521-9766Print version ISSN 1405-3195
Agrociencia vol.50 n.6 Texcoco Aug./Sep. 2016
Fitociencia
Glicerina y potasio en la actividad microbiana del suelo y nutrición y producción de remolacha (Beta vulgaris)
1Universidad de Sancti Spiritus. Avenida de los Mártires 360 esq. Carretera Central, Sancti Spiritus, Cuba.
2Universidade Estadual Paulista, Faculdade de Ciências Agrárias e Veterinária, Via de Acesso Paulo Donato Castellane, s/n, 14884-900, Jaboticabal, São Paulo, Brazil (rmprado@fcav.unesp.br).
3Universidad de Cienfuegos, Carretera a Rodas, km 3 1/2, CP 59430, Cienfuegos, Cuba.
4Universidade Federal do Piauí-UFPI, Câmpus Agrícola Socopo, Teresina, PI, Brazil.
La glicerina es un subproducto de la producción de biocombustibles (biodiésel), tiene posibilidad de uso agrícola, pero la información científica es limitada. El objetivo de este estudio fue evaluar el efecto de proporciones de glicerina en presencia y ausencia de la fertilización con potasio en la actividad microbiana del suelo, nutrición y producción de remolacha (Beta vulgaris). El estudio se realizó en un invernadero de la Universidad Estatal Paulista, Jaboticabal, Brasil, con el cv. Early Wonder de remolacha, cultivado en macetas con muestras de Alfisol. El diseño experimental fue bloques al azar, los tratamientos fueron 0, 50, 100, 200 y 400 m3 ha-1 de glicerina, en ausencia y presencia de K (150 mg dm3), con cuatro repeticiones. En el suelo se evaluó la conductividad eléctrica, actividad respiratoria, biomasa microbiana, y K disponible. En las plantas se evaluó altura, número de hojas, masa seca de la parte aérea y de los tubérculos. Tasas de 16 y 90 m3 de glicerina ha-1, en ausencia y presencia de fertilización con K aumentaron la producción de materia seca de las plantas de remolacha, pero indujeron salinidad a concentraciones altas y promovieron la disminución de la absorción de K y la actividad microbiana en el suelo.
Palabras clave: Cultivo de raíces; subproducto del biodiesel; fertilización con potasio; salinidad del suelo
The glycerin by-product of the production of biofuels (biodiesel) has possibility of agricultural use, but the scientific information is restricted. The objective of this study was to assess the effect of glycerin rates in the absence and presence of potassium fertilization on soil microbial activity, nutrition and production of beet (Beta vulgaris). The experiment was carried out in a greenhouse at Universidade Estadual Paulista, Jaboticabal, Brazil, using beet cv. Early Wonder, grown in pots with samples of Alfisol. The experimental design was randomized block, treatments were 0, 50, 100, 200 and 400 m3 ha-1 of glycerin, in the absence and presence of potassium (150 mg dm-3), with four replicates. Electrical conductivity, respiratory activity, microbial biomass, and available K were evaluated in the soil. Plant height, number of leaves, dry mass of the aerial portion and tubers were evaluated t. Glycerin at 16 and 90 m3 ha-1 rates in absence and presence of K fertilization increased the production of beet plants dry matter, but induced salinity at higher rates and promoted reduction in K absorption and the soil microbial activity.
Key words: Root crops; byproduct of biodiesel; potassium fertilization; soil salinity
Introducción
Los estudios sobre el uso de fuentes de energía renovable avanzan porque el petróleo, una fuente no renovable de energía, se agota y por la preocupación respecto al factor ambiental debido a cambios climáticos globales. La glicerina es un subproducto del biodiésel, es una fuente renovable de energía que se estudia ampliamente (Ma et al., 1998; Muñoz-Lagos et al., 2010).
La glicerina es el subproducto principal de la producción de biodiésel y su composición química depende del tipo de la materia prima usada (Thompson y He, 2006). En el proceso de producción de biodiésel el hidróxido de sodio se usa como un catalizador de transesterificación para aumentar la velocidad de la reacción química y mejorar el rendimiento industrial (Ma et al., 1998). Este procedimiento incluye aproximadamente 1 % de sodio en la glicerina cruda (Lammers et al., 2007), que podría afectar el uso de este residuo en la agricultura.
El Na es beneficioso para la nutrición de las plantas, puede reemplazar la función del K en el control osmótico, y promueve el crecimiento de la planta (Marschner, 1995), principalmente en especies clasificadas como tolerantes a salinidad, como la remolacha (Beta vulgaris) (Aquino et al., 2006). Sin embargo, el exceso de Na puede elevar el contenido de sal (Demiral, 2005) y afectar la vida microbiana del suelo (Chávez y González, 2009), reducir la absorción de nutrientes (Amini et al., 2007), y afectar el metabolismo celular que puede causar desequilibrio osmótico, alteración de las membranas, inhibición de la división y expansión celular, interrupción del equilibrio iónico, disminución de la fotosíntesis (Mahajan y Tuteja, 2005), aumento de la transpiración de la planta (Chen y Jiang, 2010) y limitar su crecimiento y el rendimiento de los cultivos (Cruz et al., 2003). Una posibilidad para aumentar la tolerancia de la planta a la glicerina con Na aumentaría la fertilización con K, ya que las concentraciones altas de ion K en las plantas aumenta la relación K/Na, asegura el metabolismo celular óptimo y el aumento de crecimiento de la planta incluso bajo estrés salino (Bartels y Ramanjulu, 2005).
Los datos del uso agronómico de la glicerina son escasos, lo que es motivo de atención al considerar el aumento en la oferta de este subproducto, debido a la creciente producción mundial de biodiésel. El conocimiento de su desempeño agronómico puede llevar a su uso eficiente en la producción de cultivos.
Las hipótesis serían si el uso de la glicerina con Na en su composición, asociado a la fertilización con K podría minimizar los daños por salinidad y permitir el uso de cantidades mayores del residuo. Sin embargo, el uso de proporciones muy altas de glicerina puede aumentar la salinidad del suelo, disminuir la actividad microbiológica, y afectar la materia seca en las plantas, a pesar de la concentración de K en el suelo.
El objetivo de este estudio fue evaluar el efecto de la glicerina en la ausencia o presencia de fertilización con K en la actividad microbiana, nutrición y producción de remolacha.
Material y métodos
El estudio se realizó en un invernadero de la Universidad Estatal Paulista, Campus Jaboticabal, Brasil (21° 15’ 22” S, 48° 18’ 58” O y altitud de 575 m) con el cv. Early Wonder de remolacha, de septiembre a noviembre de 2014.
El diseño experimental fue bloques al azar y los tratamientos fueron las dosis de glicerina: 0, 50, 100, 200 y 400 m3 h-1 en ausencia y presencia de K (150 mg dm3), con cuatro repeticiones. Glicerina cruda diluida al 14 % en agua y KCl se usó como fuente de K (K2O al 60 %). A cada maceta de 5 dm3 se agregaron 4 dm3 de Agrisol rojo-amarillo, con textura media, de acuerdo con la clasificación de EMBRAPA (1999). El análisis químico del suelo inicial se realizó de acuerdo con lo descrito por Raij et al. (2001), con los siguientes resultados: pH 5.1, 9; g materia orgánica por dm3, 0.8 K, 12 Ca, 8 Mg, 20 HAl, 20.8 SB (suma de bases: KCaMg); CEC (capacidad de intercambio catiónico: SBHAl) 40.8 mmolc dm3 y V (saturación de bases: SB100/ CCA) 51 %.
El encalado se realizó para aumentar la saturación de bases a 70 %, según Raij et al. (1997), con cal calcinada (poder de neutralización relativo total 125 %). Después de encalar, el suelo se mantuvo en bolsas de plástico 30 d para mantener la humedad a 60 % de la capacidad de campo.
La fertilización básica se incorporó al suelo en la siembra y recibió dosis de urea, superfosfato, sulfato de zinc y ácido bórico para nivelar el N (200 mg dm3), P (400 mg dm3), Zn (2 mg dm3) y B (0.5 mg dm3), según Malavolta (1981), y el K se aplicó junto con la fertilización básica antes de plantar.
Las plántulas se obtuvieron en bandejas de semillero con 200 celdas con sustrato y vermiculita y después de 30 d, dos plántulas se transfirieron a cada maceta.
La glicerina se aplicó 30 d después del trasplante y las dosis se subdividieron en dos aplicaciones semanales, según el diseño experimental. El tratamiento testigo recibió sólo agua. En todos los tratamientos la humedad se mantuvo a 70 % de la capacidad de retención del suelo. La glicerina cruda se diluyó a 14 % en agua, con 0.6% de Na,0.02% K2O y pH 6.0.
A los 30 d después del trasplante el efecto inmediato de los tratamientos se evaluó con el registro de la altura de planta, número de hojas y peso seco de los brotes y tubérculo en una planta por maceta. Además se realizó un análisis microbiológico de la capa de suelo de 0 a 10 cm de profundidad. La actividad respiratoria del suelo se determinó con la metodología de Rezende et al. (2004). El C de la biomasa microbiana se determinó según lo propuesto por Ferreira et al. (1999).
Al final del ciclo de cultivo, 70 d después del trasplante se evaluaron la altura de la planta, número de hojas y diámetro de tubérculos. La parte área y raíces del tubérculo se lavaron con una solución de detergente y una solución de ácido, ambas con agua destilada y se secaron a 60-70 °C, en un horno con aire forzado, hasta obtener la masa seca. En este momento se cuantificó la masa seca de la sección aérea y de los tubérculos; Después, el material se trituró y los análisis químicos se realizaron para determinar el contenido de K en estos órganos, según lo descrito por Bataglia et al. (1983). Los análisis químicos de suelo se realizaron para determinar la concentración de K con la metodología descrita por Raij et al. (2001) y para medir la conductividad eléctrica.
Los datos se analizaron con ANDEVA y análisis polinomiales con el programa Sisvar (Ferreira, 2011).
Resultados y discusión
La salinidad del suelo aumentó con ajuste cuadrático como una función de la glicerina con y sin K, a los 30 y 70 d después de la aplicación (Figura 1 A, B). Estos resultados pueden estar relacionados con los niveles de Na (0.6 %) en la glicerina, lo que aumenta la salinidad del suelo. Lammers et al. (2007) también reportan la presencia de Na en la glicerina y el riesgo de salinizar el suelo. La mayor salinidad se obtuvo en los tratamientos conK,comparados con aquellos sin este nutriente. Esto se debe a que las aplicaciones de glicerina con K aumentaron la concentración de este elemento en el suelo (Figura 1 C), porque la fuente usada fue KCl, que es una sal fertilizante.
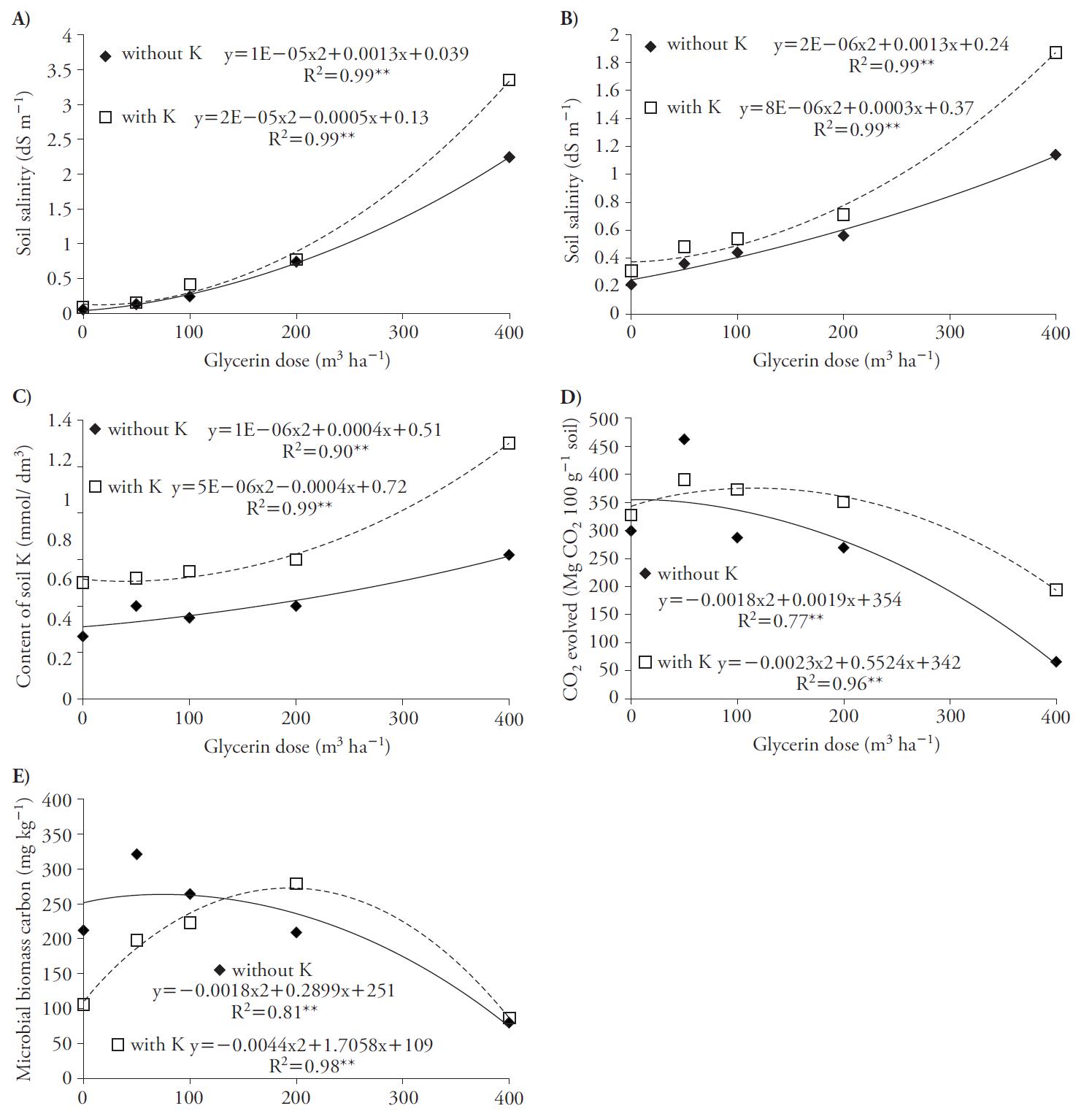
Figura 1 Índice de salinidad del suelo en 30 (A) y 70 d (B) después de aplicar glicerina, contenido de K (C) en el suelo, actividad respiratorio microbiana en el suelo (D) y C de la biomasa microbiana (E) como función de las aplicaciones de glicerina en la ausencia y presencia de K en el cultivo de remolacha (Beta vulgaris).
La aplicaciones de glicerina promovieron aumentos cuadráticos en la actividad respiratoria en el suelo, con tasas máximas de 53 y 120 m3 h-1 con y sin K (Figura1 D), y en el C de la biomasa microbiana, que alcanzó máximos de 80 y 193 m3 h h-1 con y sin K (Figura 1 E). Los suelos con glicerina y tasas de fertilización con K por encima de 53 m3 h h-1 disminuyeron la actividad microbiana, con disminución del contenido de C en la biomasa, en especial con dosis de glicerina superiores a 80 m3 h h-1, lo que podría explicarse por la salinidad mayor del suelo, ya discutida. Este efecto perjudicial en la vida microbiana en el suelo está descrito por Chávez y González (2009).
La glicerina aumentó la acumulación de K en los brotes, y el máximo fue con las dosis de 14 y 300 m3 ha h-1, con y sin K (Figura 2 A) y su acumulación se reflejó en el tubérculo, con un máximo de 59 m3 ha-1, asociado al nutriente. Sin embargo, para los tratamientos sin K acumulación en el tubérculo se redujo (Figura 2 B).
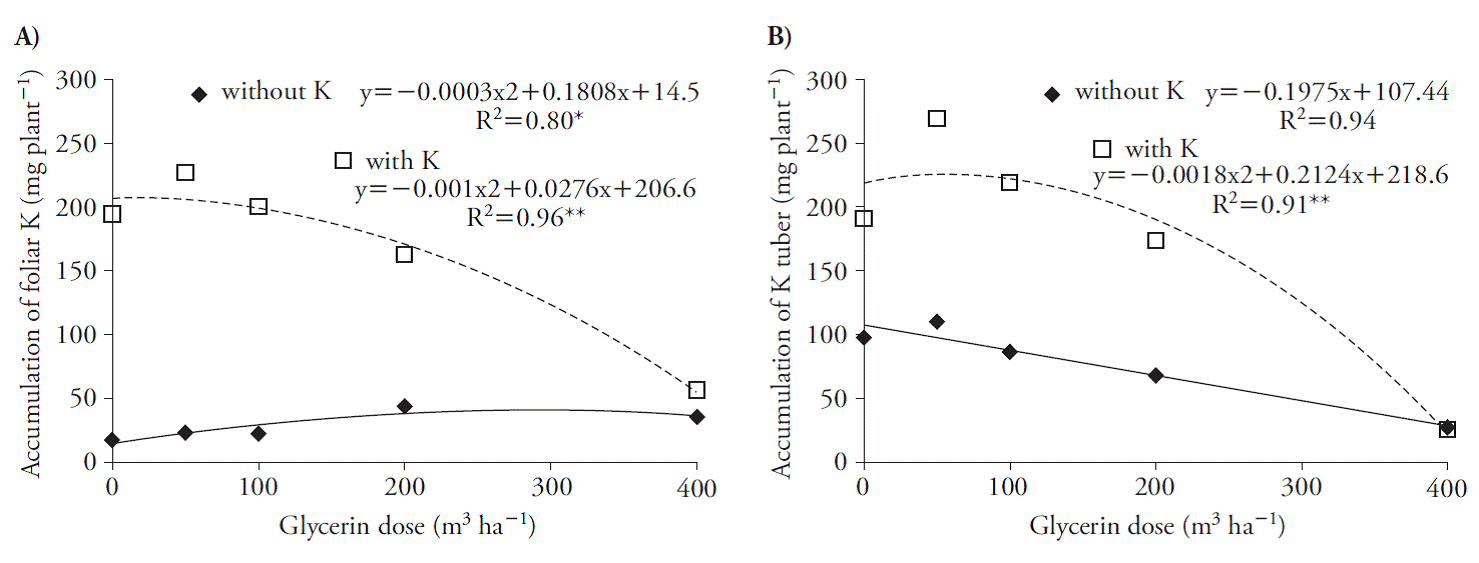
Figura 2 Acumulación de potasio en el brote (A) y tubérculo (B) de plantas de remolacha, 70 d después de aplicar los tratamientos.
Esta reducción en las plantas de remolacha debida al uso de mayores dosis de glicerina probablemente se debió al aumento de la salinidad de este residuo en el suelo (Figura 1 A, B). La concentración de K en las plantas disminuye con el aumento de la salinidad (Cornic, 2004) debido a la competencia de los iones Na y K por los acarreadores, por la similitud química de los cationes (Rodríguez-Navarro, 2000; Rus et al., 2004; Chazen et al., 2005; Wang et al, 2007).
Las concentraciones de glicerina afectaron las variables de crecimiento, independiente de la presencia o ausencia de K, excepto el diámetro del tubérculo y la materia seca (MS), 70 d después del inicio del estudio, cuando hubo interacción (Cuadro 1).
Cuadro 1 Crecimiento y rendimiento de remolacha (Beta vulgaris) a los 30 y 70 d después de aplicar los tratamientos. Jaboticabal SP, Brasil, 2014.
| Treatments | PH | NL | TD | LDM | TDM | ||||
|---|---|---|---|---|---|---|---|---|---|
| K | 30 | 70 | 30 | 70 | 70 | 30 | 70 | 30 | 70 |
| cm | cm | g pot -1 | |||||||
| Without K | 28.7 | 32.85 | 8.90 | 12.50 | 48.9 | 2.47 | 5.78 | 1.04 | 8.82 |
| With K | 28.3 | 35.05 | 8.32 | 14.05 | 52.4 | 2.32 | 6.90 | 1.07 | 10.27 |
| Test F | 0.20 ns | 5.96** | 7.99** | 18.55** | 10.30** | 1.04ns | 9.56** | 0.05ns | 18.91** |
| Glycerin(G) | |||||||||
| 0 | 33.2 | 35.87 | 9.69 | 13.25 | 53.37 | 2.77 | 6.42 | 1.42 | 10.67 |
| 50 | 34.1 | 38.25 | 9.88 | 14.75 | 61.62 | 3.72 | 8.06 | 1.83 | 13.75 |
| 100 | 32.0 | 36.00 | 8.87 | 13.62 | 56.12 | 2.89 | 7.18 | 1.28 | 10.93 |
| 200 | 29.1 | 32.87 | 8.44 | 13.25 | 51.75 | 2.04 | 5.96 | 0.73 | 9.75 |
| 400 | 14.2 | 26.75 | 6.19 | 11.50 | 30.37 | 0.57 | 4.10 | 0.0 | 2.62 |
| Test F | 75.33** | 19.54** | 42.21** | 8.41** | 95.88** | 51.28** | 13.29** | 24.47** | 124.2** |
| KG | 2.33 ns | 1.13 ns | 2.21 ns | 1.62 ns | 4.91** | 2.71 ns | 1.01 ns | 1.24 ns | 4.96** |
| CV (%) | 9.4 | 8.4 | 7.5 | 8.6 | 6.8 | 19.6 | 18.2 | 38.4 | 11.0 |
**: significativo prueba de Tukey (p≤0.01); ns: no significante. PH: altura de la planta; NL: número de hojas; TD: diámetro del tubérculo; LDM: materia seca de la hoja; TDM: materia seca del tubérculo.
En la primera evaluación, 30 d después de aplicar glicerina con y sin K hubo disminución lineal del número de hojas (Figura 3 B), de la MS de la hoja (Figura 3 C) y tubérculos (Figura 3 D) cuando se asoció al K. Sin embargo, dosis mayores a 3.5 y 11 m3 h-1 de glicerina con y sin K redujo la altura de las plantas (Figura 3 A).
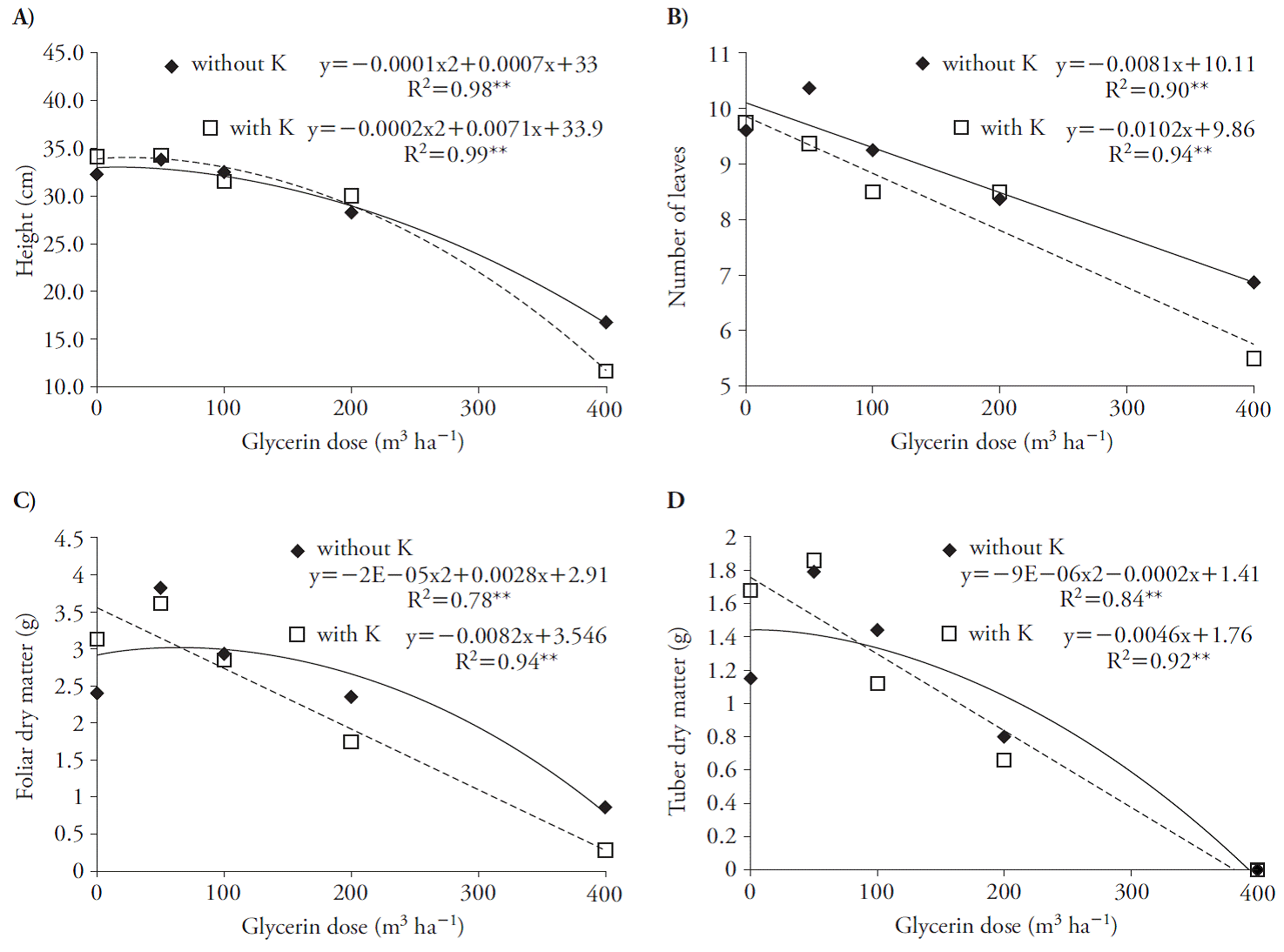
Figura 3 Altura de planta (A), número de hojas (B), materia seca de la hoja (C) y materia seca del tubérculo (D) en plantas de remolacha (Beta vulgaris) de 30 d, después de aplicaciones de los tratamientos.
Estos efectos disminuyeron la producción de MS en la planta; el uso de dosis de glicerina mayores a 70 y 43 m3 h-1, en presencia de K, redujeron las tasas de peso seco en hojas y tubérculos (Figura 3 C, D). Estos resultados indican que las plántulas de remolacha son sensibles a la glicerina, en especial si se asocia con K, debido a la salinidad, como Chinnusamy et al. (2005) reportan en plantas en etapas tempranas de crecimiento. La glicerina aumentó las variables crecimiento en la cosecha con ajuste cuadrático (Figura 4). El uso de la glicerina con y sin K alcanzó el punto máximo con dosis de 28 y 13, 60 y 80, 54 y 82, 45 y 25, 5 y 83, y 16 y 90 m3 ha-1 de altura de planta, número de hojas, MS de hojas, diámetro de la raíz tuberosa, MS de la raíz tuberosa y MS total (Figura 4).
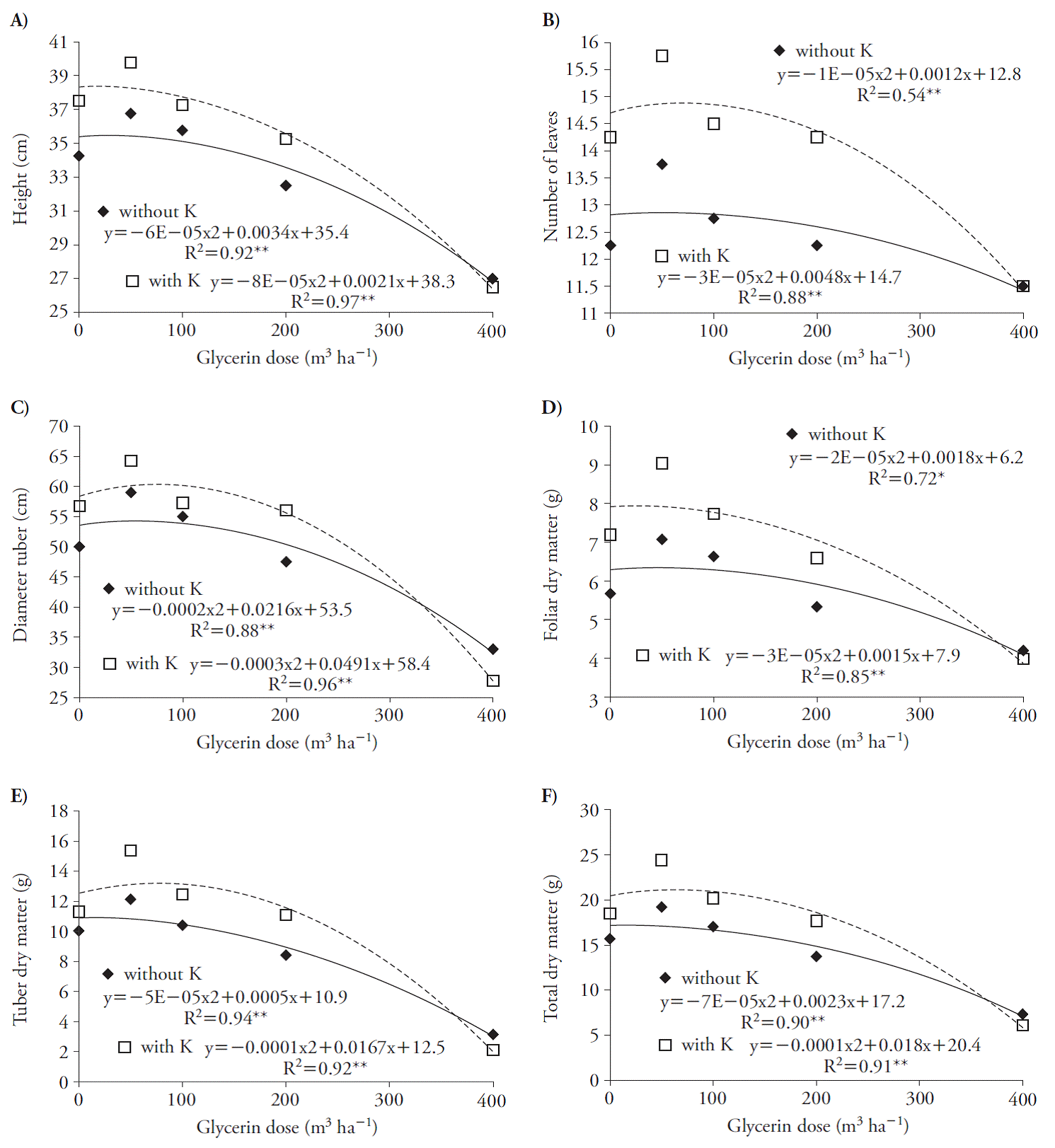
Figura 4 Altura de planta (A), número de hojas (B), diámetro de la raíz tuberosa (C), materia seca de la hoja (D), peso seco de la raíz tuberosa (E) y materia seca total (F) de plantas de remolacha (Beta vulgaris) de 70 d, después de la aplicación de los tratamientos.
El uso de dosis altas de glicerina perjudicó el cultivo, porque limita la acumulación de MS total de la planta, en especial con dosis superiores a 16 y 90 m3 ha-1 en ausencia y presencia de K (Figura 4 F). Este hecho probablemente se debe a la salinidad, porque puede afectar el desarrollo de los cultivos debido a cambios en la tasa fotosintética y en el metabolismo de los carbohidratos (Argente et al., 2009), incluyendo la remolacha, que es una especie tolerante a la salinidad en el suelo (Chávez y González, 2009). Además, la mayor disminución en la MS de la planta entera debido a la glicerina (más de 16 m3 ha-1) se produjeron sin K, probablemente por el aumento de la concentración de Na en las plantas que indujo reducción más severa en K, afectó el equilibrio iónico y la reducción del desarrollo de las plantas (Khoshgoftermanesh y Naeini, 2008).
La tolerancia de las plantas de remolacha a la glicerina fue mayor cuando se combinó con fertilización de K, ya que hubo alta acumulación de este elemento en la planta, lo cual afectó el mantenimiento de la homeostasis iónica, unproceso de importancia vital en el funcionamiento de la célula vegetal. Esto fue reportado por Bartels y Ramanjulu (2005), quienes observaron la necesidad de mantener concentraciones altas de iones K en plantas sometidas a estrés salino, para mantener la relación K+/Na+ apropiada, y asegura el metabolismo celular adecuado y el crecimiento principal de la planta.
Literature cited
Amini, F., A. A. Ehsanpour, Q. T. Hoang, and J. S. Shin. 2007. Protein pattern changes in tomato under in vitro salt stress. Russ. J. Plant Physiol. 54: 464-472. [ Links ]
Aquino, L. A., M. Puiatti, P. R. G. Pereira, F. H. F. Pereira, e I. R. Ladeira, M. R. S. Castro 2006. Produtividade, qualidade e estado nutricional da beterraba de mesa em função de doses de nitrogênio. Hortic. Bras. 24: 199-203. [ Links ]
Argente, L., D. R. López, L. M. González, R. C. López, E. Gómez, R. Girón, e I. Fonseca. 2009. Contenido de clorofila e iones en la variedad de trigo harinero Cuba-C-2004, en condiciones de estrés salino. Cultivos Trop. 30: 32-37. [ Links ]
Bataglia, O. C., A. M. C. Furlani, J. P. F. Teixeira, P. R. Furlani, e J. R. Gallo. 1983. Métodos de análise química de plantas. Instituto Agronômico, Boletim Técnico 78. Campinas, Brasil. 48 p. [ Links ]
Bartels, D., and S. Ramanjulu. 2005. Drought and salt tolerance in plants. Plant Sci. 24: 23-58. [ Links ]
Chávez, L., y L. M. González. 2009. Mecanismos moleculares involucrados en la tolerancia de las plantas a la salinidad. ITE Agraria 15: 231-256. [ Links ]
Chazen, O., W. Hartung, and P. M. Neumann. 2005. The different effects of PEG 6000 and NaCl on leaf development are associated with differential inhibition of root water transport. Plant Cell Environ. 18: 727-735. [ Links ]
Chen, H., and J. Jiang. 2010. Osmotic adjustment and plant adaptation to environmental changes related to drought and salinity. Environ. Rev. 18: 309-319. [ Links ]
Chinnusamy, V., A. Jagendorf, and J. K. Zhu. 2005. Understanding and improving salt tolerance in plants. Crop Sci. 45: 437-448. [ Links ]
Cornic, G. 2004. Drought stress and high light effects on leaf photosynthesis. In: Baker, N. R., and J. R. Bowyer (eds). Photoinhibition of Photosynthesis: From Molecular Mechanisms to the Field. BIOS Scientific Publishers. Oxford, UK. pp: 297-313. [ Links ]
Cruz, J. C., S. Pelacani, and W. Dos Santos. 2003. Production and partitioning of dry matter and stomatal conductance of rangpur lemon under salt stress. Rev. Bras. Frutic. 25: 528-531. [ Links ]
Demiral, A. 2005. Comparative response of twoo live (Olea europea L.) cultivars to salinity. Turk. J. Agric. Forest. 29: 267-274. [ Links ]
EMBRAPA (Empresa Brasileira de Pesquisa Agropecuária). 1999. Centro Nacional de Pesquisa de Solo. Sistema Brasileiro de Classificação de Solos. Embrapa Produção de Informações. Brasilia, Brasil. 412 p. [ Links ]
Ferreira, A. S., F. A. O. Camargo, e C. Vidor. 1999. Utilização de microondas na avaliação da biomassa microbiana do solo. Rev. Bras. Ciência Solo 23: 991-996. [ Links ]
Ferreira, D. F. 2011. Sisvar: a computer statistical analysis system. Ciência Agrotec. 35: 1039-1042. [ Links ]
Khoshgoftermanesh, A. H., and M. R. Naeini. 2008. Salinity effect on concentration, uptake, and relative translocation of mineral nutrients in four olive cultivar. J. Plant Nutr. 31: 1243-1256. [ Links ]
Lammers, P. J., M. S. Honeyman, K. Bregendahl, B. Kerr, and T. Weber. 2007. Energy value of crude glycerol fed to pigs. Iowa State University Animal Industry Report 2007. Disponible: Disponible: http://www.ans.iastate.edu/report/air/2007pdf/R2225.pdf . (Acess: October, 2014). [ Links ]
Ma, F., L. D. Clements, and M.A. Hanna. 1998. The effects of catalyst, free fatty acids, and water on transecterification of beef tallow. Am. Soc. Agric. Eng. 4: 1261-1264. [ Links ]
Mahajan, S., and N. Tuteja. 2005. Cold, salinity and drought stresses: An overview. Arch. Biochem. Biophys. 444: 139-158. [ Links ]
Malavolta, E. 1981. Manual de Química Agrícola. Adubos e Adubações. 3rd ed. Agronômica Ceres. São Paulo, Brasil. 596 p. [ Links ]
Marschner, H. 1995. Mineral Nutrition of Higher Plant. Academic Press., London UK. 889 p. [ Links ]
Muñoz-Lagos, R. E., R. A. Ortega-Blu, L. G. Acosta-Espejo, y R. A. González-Platteau. 2010. Biocombustibles en Chile. II. Evaluación económica de la elaboración de biocombustibles. Agrociencia 44: 849-859. [ Links ]
Raij, B. van., H. Cantarella, J. A. Quaggio, and A. M. C. Furlani. 1997. Recomendações de adubação e calagem para o estado de São Paulo. 2nd ed. Instituto Agronômico. Boletim Técnico 100. Campinas, Brasil. 285 p. [ Links ]
Raij, B. van , J. C. Andrade, H. Cantarella, and J. A. Quaggio. 2001. Análises químicas para avaliação da fertilidade de solos tropicais. Instituto Agronômico. Campinas, Brasil. 285 p. [ Links ]
Rezende, L. A., L. C. Assis, and E. Nahas. 2004. Carbon, nitrogen and phosphorus mineralization in two soils amended with distillery yeast. Biores. Tech. 94: 159-167. [ Links ]
Rodríguez-Navarro, A. 2000. Potassium transport in fungi and plants. Biochi. Biophys. Acta 1469: 1-30. [ Links ]
Rus, A., B. Lee, A. Muñoz., A. Sharkhuu, J. Zhu, J. Bressan, and R. P. Hasegawa. 2004. AtHKT1 facilitates Na+ homeostasis and K+ nutrition in planta. Plant Physiol. 136: 2500-2511. [ Links ]
Thompson, J. C., and B. B. He, 2006. Characterization of crude glycerol from biodiesel production from multiple feed stocks. Appl. Engin. Agric. 22: 261-265. [ Links ]
Wang, S. M., J. L. Zhang, and T. J. Flowers. 2007. Low-affinity Na+ uptake in the halophyte Suaeda maritima. Plant Physiol. 145: 559-571. [ Links ]
Recibido: 01 de Abril de 2015; Aprobado: 01 de Abril de 2016
 This is an open-access article distributed under the terms of the Creative Commons Attribution License
This is an open-access article distributed under the terms of the Creative Commons Attribution License
