










Services on Demand
Journal

Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
 Cited by SciELO
Cited by SciELO Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkAgrociencia
On-line version ISSN 2521-9766Print version ISSN 1405-3195
Agrociencia vol.46 n.7 Texcoco Oct./Nov. 2012
Biotecnología
Total phenolic compounds, flavonoids and antioxidant activity in the flowers of Crataegus spp. from México
Compuestos fenólicos totales, flavonoides y actividad antioxidante en las flores de Crataegus spp. de México
Rosario García–Mateos1,4, Leonor Aguilar–Santelises2, Marcos Soto–Hernández3*, Raúl Nieto–Angel4, Geoffrey Kite5
1 Departamento de Preparatoria Agrícola, Universidad Autónoma Chapingo. 56230. Chapingo, Estado de México, (rosgar08@hotmail.com)
2 Facultad de Estudios Superiores Zaragoza. UNAM. Iztapalapa, México, D.F. 09230. Iztapalapa, México. (lsante02@yahoo.com.mx).
3 Botánica. Campus Montecillo. Colegio de Postgraduados. 56230. Montecillo, Estado de México. *Author for correspondence: (msoto@colpos.mx).
4 Fitotecnia. Universidad Autónoma Chapingo. 56230. Chapingo, Estado de México, (r.nietoangel@gmail.com).
5 Royal Botanic Gardens, Kew. Richmond. Surrey UK. (gkite@kew.org).
Received: febrero, 2012.
Approved: octubre, 2012.
Abstract
There are a number of medicinally active phytochemicals isolated from hawthorn with most of the data generated in studies of those species that are native to Europe and Asia. Comparatively little is known about the North American, specifically the Mexican Crataegus species. The objective of this study was to evaluate phenolic compounds, flavonoids and antioxidant activity of extracts of flowers from six Mexican accessions of Crataegus in order to contribute to the chemistry of the Germplasm Bank accessions at the Universidad Autonóma Chapingo. Antioxidant activity was evaluated using DPPH (2,2–diphenyl–1–picrylhydrazyl) method and the flavonoids were identified by HPLC–MS. Quercetin– 3–O–glucoside, quercetin–3–O–rhamnoside, quercetin 3–O–rhamnosyl–(1 6)–glucoside and quercetin 3–O–rhamnosyl–(12)–[rhamnosyl–(16)]–glucoside were identified. Using the DPPH radical scavenging capacity assay, accessions 52 and 77 showed the greatest antioxidant activity (IC50=431.1±27.2 and 555.4±8.0 µg mL–1). Only flavonoids glycosides were identified in all six species and the phenolics content was higher than flavonoids. There were not found differences in the phenolics total content.
6)–glucoside and quercetin 3–O–rhamnosyl–(12)–[rhamnosyl–(16)]–glucoside were identified. Using the DPPH radical scavenging capacity assay, accessions 52 and 77 showed the greatest antioxidant activity (IC50=431.1±27.2 and 555.4±8.0 µg mL–1). Only flavonoids glycosides were identified in all six species and the phenolics content was higher than flavonoids. There were not found differences in the phenolics total content.
Keywords: Crataegus, flavonoids, flowers, Mexican hawthorn, radical scavenging activity.
Resumen
Hay una serie de fitoquímicos medicinalmente activos aislados del espino (Tejocote), con la mayoría de los datos obtenidos de estudios sobre esas especies nativas de Europa y Asia. Comparativamente, poco se sabe acerca de las especies de América del Norte, específicamente la especie mexicana Crataegus. El objetivo de este estudio fue evaluar los compuestos fenólicos, flavonoides y la actividad antioxidante de extractos de flores de seis recolectas mexicanas de Crataegus para contribuir a la química de las recolectas del Banco de Germoplasma de la Universidad Autónoma Chapingo La actividad antioxidante se midió con el método DPPH (2,2–di–fenil–1–picrilhidracilo) y los flavonoides se identificaron con HPLC–MS. La quercetina–3–O–glucósido, quercetina–3–O–ramnósido, quercetina 3–O–ramnosil–(16)–glucósido y quercetina 3–O–ramnosil–(12)–[ramnosil–(16)]–glucósido fueron identiicadas. Usando la prueba de DPPH con capacidad de barrido de radicales, las recolectas 52 y 77 mostraron la mayor actividad antioxidante (IC50 = 431.1 ±27.2 y 555.4±8.0 µg mL–1). Sólo se identificaron glucósidos de flavonoides en todas las seis especies y el contenido de fenólicos fue mayor que los flavonoides. No se encontraron diferencias en el contenido total de compuestos fenólicos.
Palabras clave: Crataegus, flavonoides, flores, espino mexicano, capacidad de barrido de radicales.
INTRODUCTION
Crataegus is a large genus of shrubs or trees from Eurasia and America. Species of Crataegus native to North America make up approximately half of all known species of the genus (Phipps et al., 2003). In México exists 13 species known as "tejocote" (Phipps, 1997; 1998). The plants are also known by various local names such as "manzanita", "manzanilla" and "carasu" in Tarasco (Michoacan state), "belohui" in Zapoteco (Oaxaca state) and "pedyi" in Mazahua (Estado de México), and these often do not correspond to species (Martínez, 1979).
In México, there is an increasing interest in under–utilized species, also known as minor, secondary or alternatives species, and it is the case of tejocote (Nieto–Angel, 2007). Tejocote has several traditional medicinal uses: flowers, leaves and fruits being used to treat ailments such as heart or cough problems. However, in México the medicinal properties of tejocote have not been fully investigated.
The wide diversity and genetic variability of tejocote in México is represented by 166 accessions maintained by the Germplasm Bank of the Universidad Autónoma Chapingo, México (Pérez–Ortega et al., 2004; Nuñez–Colín et al., 2008). Using 37 morphological variables 92 accessions were grouped in five (Nieto–Angel, 2007; Nieto–Angel et al., 2009); but no detailed taxonomic, phytochemical or molecular studies have been performed. According to Phipps (1997), there are difficult taxonomic problems in Mexican Crataegus.
Phytochemical investigations of the Crataegus genus have centered on the identification of phenolic compounds and flavonoids (Liu et al., 2011; Edwards et al., 2012). They are used as taxonomic markers because the patterns tend to be species specific; they are also used to recognize new varieties (Van Sumere et al., 1993; Iwashina, 2000). For example, ecotypes of Crataegus azarolus collected in different regions of Italy showed variation in the sugar phenolic content of their fruits (Bignami et al., 2003). The presence of phenolic compounds justifies in part the pharmacological activity reported for Crataegus (Craig, 1999; Chang et al., 2002; Cui et al., 2006). Beneficial effects are attributed to the main active constituents, its flavonoid and proanthocyanidin components (Chang et al., 2002; Cui et al., 2006; Liu et al., 2011). Various species of Crataegus are used in Europe and Asia for their medicinal properties (Ody, 1993; Craig, 1999; Chang et al., 2002). Chinese hawthorn (Crataegus pinnatifida) extracts have beneficial effects, such as antioxidants, anti–inflammatory and hypo lipidemic (Cui et al., 2006), especially on the cardiovascular system (Chang et al., 2002), as well as positive effects on the brain and vascular endothelium (Zhang et al., 2002). Crataegus spp. are used for treatment of heart failure, hypertension, atherosclerosis, angina pectoris, indigestion, and abdominal distension (Chang et al., 2002). The leaves, fruit, and flowers are used for improving the pumping capacity of heart and for treating angina, causing dilation of the smooth muscles of the coronary vessels, thereby increasing blood flow and reducing the tendency for angina (Craig, 1999). These beneficial effects are attributed to its flavonoid and proanthocyanidin components (Chang et al., 2002).
The aim of this study was to determine the total phenolic compounds, flavonoids and antioxidant activity of crude flower extracts from six Mexican accessions of Crataegus spp. in order to contribute to the chemistry of the Germplasm Bank accessions at the Universidad Autonóma Chapingo. The hypothesis was that the composition profiles of phenolic compounds of flowers are different in the Mexican species comparing with those of Europe and Asia.
MATERIALS AND METHODS
Plant material
Flowers of tejocote were obtained from the Germplasm Bank of the Universidad Autonoma Chapingo, México (19° 29' N and 98° 53' W, and 2240 m altitude; García, 1988). The climate is C(Wo) (w)b (I')g, with moderate rain, temperate and the driest of the sub humid climates, with rains in summer; the mean annual temperature is 17.8 °C and the annual rainfall is 644.8 mm (García, 1988).
Preparation of extracts
Flower samples were taken randomly from six accessions (probable specie) representative of the five groups in the Germplasm Bank (Nieto–Angel, 2007; Nieto–Angel et al., 2009). Then 1 g of ground and dried sample was weighed, and each one was suspended in 20 mL 95 % ethanol. After 24 h, the volume was adjusted to 25 mL with 80 % ethanol, and the mixture was centrifuged at 1409 g.
Quantification of total phenols
The total phenols content was quantified according to the method proposed by Waterman and Mole (1994). The concentration was obtained from a standard curve based on phenol (y = 0.00l4x; R2=0.997). Results were expressed as mg of phenol equivalent per 1 g of dry tissue.
Quantification of total flavonoids
An aliquot of the ethanolic extract (0.5 mL) was mixed with 1.5 mL of 95 % ethanol, 0.1 mL of 10 % AlCl3, 0.1 mL of 1M potassium acetate and 2.8 mL of distilled water. After 30 min at room temperature the absorbance was obtained, at 415 nm, and a standard curve (y=0.0122x – 0.0067; R2=0.965) was obtained using quercetin (Sigma–Aldrich®, St Louis Missouri, USA) as standard. Results were expressed as quercetin equivalents (mg of quercetin equivalent per 1 g of dry weight). The analyses were carried out in triplicate (Chang et al., 2002).
Identification of flavonoids by HPLC–MS
Analyses were performed using a Termo Scientific HPLC–MS (Termo Scientific, Walthman, MA, USA) system consisting of autosampler, pumps and PDA and a LCQ classic ion trap mass spectrometer (Termo Scientific, Walthman, MA, USA), fitted with an API source. Chromatography was performed on a 150 × 4.6 mm i.d., 5 µm Phenomenex Luna C18 column (Torrance CA, USA), using a 1 mL min–1 linear mobile phase gradient of 20 to 50 % aqueous methanol (containing 1 % acetic acid) in 30 min. The compounds were identified by interpretation of their mass spectra, through the MS/MS Library ViewTM Software (AB Sciex, Foster City CA, USA) and using an in–house library of MS/MS spectra of flavonoid aglycones, compiled from authentic standards (Cuychens and Claeys, 2004).
Antioxidant activity
The analysis was made following the method proposed by Amico et al. (2008), which is based on the transformation of the DPPH*(2,2–diphenyl–1–picrylhydrazyl, Sigma–Adrich®; St. Louis, Missouri, USA) as stable radical to reduced DPPH, reaction was monitored at 515 nm by the gradual color change (purple to yellow), which increases gradually according to the number of electrons accepted (Cotelle et al., 1996).
Ten g of dry flowers were suspended in 100 mL of 80 % methanol, left for 48 h at 20 °C, filtered and the extract was evaporated to dryness in a Büchi evaporator R–210® (Flawil, Switzerland) and lyophilized in a freeze dryer Labconco, Mod 75034 (Labconco, Co. Missouri, USA). Prior to the analysis of the antioxidant activity, both the solution of DPPH* and of the sample were maintained in an ice bath. To calculate the 50 inhibitory concentration (IC50) three concentrations (1.0, 0.5 and 0.1 mg mL–1 in methanol) of each extract were prepared; 0.1 mL of each solutions was mixed with 3.9 mL of 0.1mM DPPH*. Immediately, zero time and after 30 min reading were made, at 515 nm, in a Spectronic 20® spectrophotometer (Rochester, N Y, USA). Quercetin solution (1.0 mg mL–1) was used as positive control. The low absorbance of the reaction mixture showed high antioxidant activity.
The initial concentration of DPPH* radical in methanol was controlled for every experiment from a calibration curve (y=0.016x + 0.033; R2=0.994) made by measuring the absorbance at 515 nm of standard DPPH* solutions at different concentrations (Amico et al., 2008). The percentage of DPPH* was calculated with the formula: % DPPH* = (Acontrol – Asample)*100/Acontrol, where Acontrol is the control absorbance (DPPH* 0.1mM), Asample is the absorbance obtained after 30 min of each sample with 0.1 mM DPPH*. To calculate the IC50, a linear regression was performed of the calibration curve (% of DPPH* vs. concentration of the added extract). IC50 values are concentration of the sample required to scavenge 50 % free radical DPPH.
Statistical analysis
This study consisted of six accessions with three replicates each one. A replication consisted of three samples of flowers from the same tree (experimental unit) An ANOVA was performed and means were compared using Tukey test (p<0.05) by means of SAS (1996).
RESULTS AND DISCUSSION
Quantification of phenols and flavonoids
Content of phenols was higher that flavonoids (Table 1). Bahorum et al. (1994) observed the same behavior in extracts of C. monogyna. Content of phenolic compounds was not statistically different within accessions (p>0.05) (Table 1). Results (7.7714.87 mg g–1 dry weight) were similar to that in flowers of seven species from Europe and Asia (0.45 – 10.14 mg g–1 dry weight) (Edwards et al., 2012).
The flavonoid content (2.85–7.23 mg g–1 dry weight) was lower than in 12 European species (3.18 – 19.4 mg g–1 dry weight) (Edwards et al., 2012). A tendency of the flavonoids content was not observed within origin and type of accession. Edwards et al. (2012) pointed out the wide variability of flavonoid content on hawthorn within and between species. Flavonoid content was highest in accessions 48 and 77 and lowest in accession 11 (Table 1). The difference in flavonoid content is possibly due to the presence of procyanidins or other phenolic acids as described by Cui et al. (2006) and Liu et al. (2011) for fruits of C. pinnatifida containing procyanidins (19.7 %), phenols (1.27 % chlorogenic acid) and flavonoids (0.48 %). Bignami et al. (2003) report similar results of total flavonoids and phenols content in five accessions of C. azarolus L. flowers. The environmental factors do not seem to influence the phenolic compounds content among species and accessions analyzed, and the variations of the metabolites may be of genetic origin (Bignami et al., 2003). There is no relationship between the total phenolic content and the antioxidant activity of berry, fruit wines and liquors (Robards et al., 1999), a fact which should be taken into account when interpreting the results of any study, because some others metabolites can contribute to this activity. Therefore, it is useful to associate individual phenolic compounds with antioxidant activity because their particular structural characteristics can easily neutralize free radicals (Bors et al., 1990; Cotelle et al., 1996; Robards et al., 1999).
In our study the highest percentage of flavonoids was 0.72 %, close to the values described by Chang et al. (2002) and by Melikoglu et al. (2004) in leaves and flowers of Crataegus spp. and C. microphylla (0.7 %). According to Chang et al. (2002), flavonoid content varies from 0.1 to 1.0 % in Crataegus sp. fruits and from 1 to 2 % in leaves and flowers, which could be due to the method of extraction and analysis. However, the variation in the content of these metabolites therein some Crataegus species may be related to environmental conditions. The phenolic composition of fruits is determined by genetic and environmental factors but may be modified by oxidative reactions during processing and postharvest (Robards et al., 1999). Kirakosyan et al. (2003; 2004) found high levels of phenolic compounds in leaves of C. laevigata and C. monogyna grown under drought and cold stress conditions.
Identification of flavonoids by HPLC–MS
The analysis by HPLC–MS allowed to tentatively identify four glycosides of quercetin (Figure 1; Table 2). Using soft ionization techniques such as electrospray with serial mass spectrometry, the molecular mass and product ion spectrum of the aglycone moiety of a flavonoid O–glycoside can be obtained relatively easily, particularly in positive ion mode, and the aglycone spectrum can be compared with those of standards in a mass spectral library. All accessions showed the typical UV spectra of quercetin 3–O–glycosides, and serial MS analysis confirmed quercetin as the aglycone. Glycosides 1, 3 and 4 have not been reported in Crataegus from America. Flavonoid 2 was confirmed as rutin (quercetin 3–O–rhamnosyl–(16)–glucoside) against a standard, and it was assumed that glucose was also primary glycosidic sugar in the mono hexoside 3 and the branched tri glycoside 1, quercetin–triglycoside have not been reported in Crataegus genus (Liu et al., 2011). The serial MS spectra of 1 matched as quercetin 3–O–rhamnosyl–(1®2)–[rhamnosyl(16)]–glucoside in an in–house spectral library. Besides the analysis of the MS/MS spectra through the Library View TM Software (from AB Sciex, Foster City CA, USA) allowed to confirm these observations. The flavonoid 4 showed the loss of 146 following MS/MS and so was assigned as quercetin 3–O– rhamnoside (quercitrin).
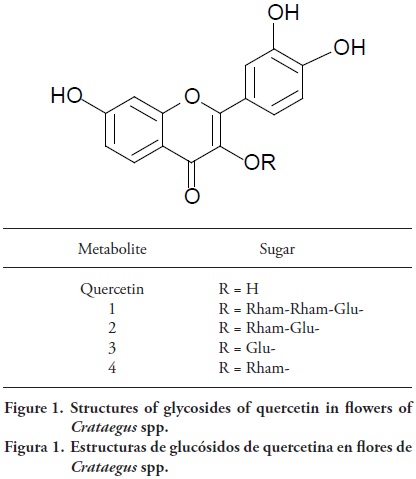
Quercetin is present in the European species C. microphyila, C. oxyacantha, C. sinaica and C. monogyna (Rehwald et al., 1994; Shahat et al., 2002; Melikoglu et al., 2004). Furthermore, Urbonaviciute et al. (2006) and Zhang et al. (2001) suggested that rutin is commonly detected in C. monogyna. Rohr et al. (1999) identified by HPLC (–)–epicatechin and several procyanidins in Crataegus leaves and flowers, whereas Dauguet et al. (1993) identified some derivatives of kaempferol and other flavonoids from bee pollen of C. monogyna. Vitexin–2"–O–rhamnoside, acetylvitexin–2"–O–rhamnoside, rutin, hyperoside, vitexin, and chlorogenic acid are found in leaves and flowers of C. monogyna (Rehwald et al., 1994). Fong and Bauman (2002) point out that the composition profile of phenolics in European hawthorn species also suggest that flavonoids were the markers for the differentiation of C. monogyna, C. laevigata, and C. pentagyna from one another.
The flavonoid glycosides 1–4 differed qualitatively or quantitatively between the six accessions. Accessions 2 and 6, as well as 52, had the same flavonoids but their relative abundance was different (Table 3). Also these accessions showed four glycosides and they had greatest relative abundance of quercetin 3–O–rhamnoside and accessions 11 (58 %), 48 (54 %) and 77 (66 %) showed greatest relative abundance of rutin. These accessions showed only three glycosides (1, 2, and 3), and its flavonoids profile was different to those of Europe and Asia (Rehwald et al, 1994; Kirakosyan et al, 2003; Cui et al, 2006).
Glycoside 4 was the most abundant in the accessions 2 (58 %) and 6 (88 %) probably due to the fact that they belong to the same specie (C. stipulosa) and have the same origin (Chiapas). Glycoside 2 was found in higher proportion in the accessions 11 (58 %) and 48 (54 %) of C. mexicana, from Puebla. In the accessions 52 and 77 of C. nelsoni the glycosides 4 and 2 were the most abundant (62 and 66 %) and they come from Chiapas. These differences could be due to a genetic factors (similar within species) and not to their origin (Bignami et al, 2003). Edwards et al. (2012) point out that this discrepancy indicate the need for a long–term phytochemical study performed using a validated analytical method, and involves looking at the chemistry of a single species collected from multiple locations over a number of years. Such study may answer the question of whether previously reported concentration values vary as a result of species, location, seasonality, or research methodology. It would also allow for quantitative determination of chemotaxonomic differences between the species.
Antioxidant activity
The antioxidant activity of extracts is due, in part, to the quercetin, but the presence of flavones and procyanidins, not studied in the present work, also might contribute to this activity (Robards et al., 1999; Harborne and Williams 2000; Chang et al., 2002). To this respect, the antioxidant activity of flavonols and the flavones is associated with their chemical structure. Those molecules with hydroxyl groups in 3' and 4' positions of the B ring and OH in C–3 permit stable and efficient structures to capture free radicals, a requirement for the maximum antioxidant capacity (Bors et al., 1990; Cotelle et al., 1996; Robards et al., 1999,). In this case, quercetin having a 3,5,7,3',4'–pentahydroxy group showed stronger DPPH radical scavenging activity than kaempferol; this has 3,5,7,4'–tetrahydroxy group, in the B–ring of flavonol skeleton, which might enhance DPPH radical reducing activity (Rice–Evans et al, 1996; Okawa et al., 2001). It is important to mention that some of the glycoside derivatives present lower antioxidant activity than the aglycones, since the hydroxy group at C–3 in 3–O–glycoside may not contribute to scavenge the DPPH radical (Okawa et al., 2001). This information explains the moderate antioxidant activity found in relation to the glycosides structure in the present study.
Pharmacokinetic studies indicate that the sugar fraction in the quercetin is important because retards its absorption due to the fact that the flavonol is absorbed well in the small intestine, whereas rutin is absorbed in the colon, after intestinal bacteria hydrolysis (Harborne and Williams, 2000).
Other studies indicate properties of the glycosides of quercetin; for instance higher spasmolytic activity than of quercetin has been demonstrated, due to its effect on the relaxation of the skeletal muscle. Additionally, numerous physiological effects beneficial to human health are attributed to the flavonoids as anticancer agents. Their action in vitro with low density lipoproteins inhibits the development of vascular diseases when they are consumed in a diet of fruits and vegetables (Rice–Evans et al., 1996; Okawa et al., 2001; Schramm and German, 1998).
In this study were analyzed the type of flavonoids present in species of Crataegus found in Mexico. Furthermore, the profile and the abundance of the flavonoids detected in six accessions of the Germplasm Bank of the Universidad Autónoma Chapingo, Mexico, will contribute to the chemistry of the 92 accessions morphologically characterized. Analysis of more accessions of the different plant species would be necessary to determine whether these differences were species specific or due to inter–individual variation (Edwards et al., 2012).
CONCLUSIONS
There are not differences in the content of total phenolics within accessions; it was not observed a tendency of the content of flavonoids within the origin and the type of accession neither. The content of total phenolics was similar to those of the flowers on seven species from Europe and Asia. Four flavonol glycosides were detected in flowers of Mexican species of Crataegus and provide evidence of the antioxidant activity of the extracts of flowers. The flowers of one accession presented the higher antioxidant activity and contrasts to the other accessions. This is the first report of composition and content of phenolic compounds and profile of flavonoids in the flowers of Crataegus spp. from Mexico.
LITERATURE CITED
Amico, V., R. Chillemi, S. Mangiafico, C. Spatafora, and C. Tringali. 2008. Polyphenol–enriched fractions from Sicilian grape pomace: HPLC–DAD analysis and antioxidant activity. Bioresour. Technol. 99: 5960–5966. [ Links ]
Bahorum T., F. Trotin, J. Pommery, J. Vasseur, and M. Pinkas. 1994. Antioxidant activities of Crataegus monogyna extracts. Planta Med. 60: 323–328. [ Links ]
Bignami, C., M. Paolocci, A. Scossa, and G. Bertazza. 2003. Preliminary evaluation of nutritional and medicinal components of Crataegus azarolus fruits. Acta Hort. 597:95–100. [ Links ]
Bors, W., W. Heller, C. Michel, and S. Manfred. 1990. Flavonoids as antioxidants: determination of radical–scavenging efficiencies. Methods Enzymol. 186: 343–355. [ Links ]
Chang, C., M. Yang, H. Wen, and J. Chern. 2002. Estimation of total flavonoids content in propolis by two complementary colorimetric methods. J. Food Drug Anal. 10: 176–182. [ Links ]
Chang, Q., Z. Zuo, F. Harrison, and M. S. Chow. 2002. Hawthorn. J. Clin. Pharmacol. 42: 605–612. [ Links ]
Cotelle, N., J. Bernier, J. Catteau, J. Pommery, J. Wallet, and E. M. Gaydou. 1996. Antioxidant properties of hydroxy–flavones. Free Radical Bio. Med. 20: 35–43. [ Links ]
Craig, W. J. 1999. Health–promoting properties of common herbs. Am. J. Clin. Nutr. 70 (Suppl.): 491S–499S. [ Links ]
Cui N., K. Nakamura, S. Tian, H. Kayahara, and Y. Tian. 2006. Polyphenolic content and physiological activities of Chinese Hawthorn extracts. Biosci. Biotech. Biochem. 70: 2948–2956. [ Links ]
Cuychens, F., and M. Claeys. 2004. Mass spectra in the structure analysis of flavonoids. J. Mass Spectrom. 39: 1–15. [ Links ]
Dauguet, J. C., M. Bert, J. Dolley, A. Bekaert, and G. Lewin. 1993. 8–methoxykaempferol, 3–neohesperidoside and other flavonoids from pollen of Crataegus monogyna. Phytochemistry 33: 1503–1505. [ Links ]
Edwards, J. E., P. N. Brown, N. Talent, T. A. Dickinson and P. R. Shipley. 2012. A review of the chemistry of the genus Crataegus. Phytochemistry 79: 5–26. [ Links ]
Fong, H. H. S., and J. L Bauman. 2002. Hawthorn. J. Cardiovasc. Nurs.16: 1–8. [ Links ]
García, E. 1988. Modificaciones al Sistema de Clasificación Climática de Kópen. Editorial UNAM. México, D. F., México. 246 p. [ Links ]
Harborne, J. B., and C. A. Williams. 2000. Review advances in flavonoid research since 1992. Phytochemistry 55: 481–504. [ Links ]
Iwashina, T. 2000. The structure and distribution of the flavonoids in plants. J. Plant Res. 113: 287–299. [ Links ]
Kirakosyan, A., P. Kaufman, S. Warbe, S. Zick, K. Aaronson, S. Bolling, and S. Ch. Chang. 2004. Applied environmental stresses to enhance the levels of polyphenolics in leaves of hawthorn plants. Physiol. Plantarum 121: 182–186. [ Links ]
Kirakosyan, A., E. Seymour, P. B. Kaufman, S. Warber, S. Bolling, and S. Ch. Chang. 2003. Antioxidant capacity of polyphenolic extracts from leaves of Crataegus laevigata and Crataegus monogyna (Hawthorn) subjected to drought and cold stress. J. Agr. Food Chem. 51: 3973– 3976. [ Links ]
Liu, P., H. Kallio, and B. Yang. 2011. Phenolic compounds in hawthorn (Crataegus grayana) fruits and leaves and changes during fruits ripening. J. Agric. Food Chem. 59: 11141–11149. [ Links ]
Martínez, M. 1979. Catálogo de Nombres Vulgares y Científicos de Plantas Mexicanas. Fondo de Cultura Económica. México, D. F., México. 1247 p. [ Links ]
Melikoglu, G., L. Bitis, and A. H. Mericli. 2004. Flavonoids of Crataegus microphylla. Nat. Prod. Res. 18: 211–213. [ Links ]
Nieto–Ángel, R. 2007. Colección, conservación y caracterización del tejocote (Crataegus spp.). In: Nieto–Angel, R. (ed). Frutales Nativos, un Recurso Fitogenético de México. Universidad Autónoma Chapingo. Estado de México, México. pp: 25–107. [ Links ]
Nieto–Ángel, R., S. A. Pérez–Ortega, C. A. Nuñez–Colín, J. Martínez–Solís, and F. González–Andrés. 2009. Seed and endocarp traits as markers of the biodiversity of regional sources of germoplasm of tejocote (Crataegus spp.) from Central and Southern México. Scientia Horticulturae 121 (2): 166–170. [ Links ]
Núñez–Colín, C. A., R. Nieto–Ángel, A. F. Barientos–Priego, S. Segura, J. Sahagún–Castellanos, y C. González–Andrés. 2008. Distribución y caracterización eco–climática del género Crataegus L. (Rosaceae, Subfam. Meloideae) en México. Rev. Chapingo Serie Hort. 14: 177–184. [ Links ]
Ody, P. 1993. The Herb Society's Complete Medicinal Herb. Dorling Kindersley Publishing. New York, USA. 192 p. [ Links ]
Okawa, M., J. Kinjo, T. Nohara, and M. Ono. 2001. DPPH (1,1–diphenyl–2–picrylhydrazyl) radical scavenging activity of flavonoids obtained from some medicinal plants. Biol. Pharm. Bull. 24: 1202–1205. [ Links ]
Pérez–Ortega, S. A., C. A. Núñez–Colín, R. Nieto–Ángel, A. F. Barrientos–Priego, y S. Segura. 2004. Los recursos genéticos de tejocote (Crataegus spp.) en México. Variación eco–climática. Proc. Interamer. Soc. Trop. Hort. 48: 149–151. [ Links ]
Phipps, J. B. 1997. Monograph of Northern Mexican Crataegus (Rosaceae, Subfam. Maloideae). Sida Botanical Miscelany No. 15. Botanical Research Institute of Texas, Inc. USA. 94 p. [ Links ]
Phipps, J. B. 1998. Introduction to the red fruited hawthorns (Crataegus, Rosaceae) of western North America. Can. J. Bot. 76: 1863–1899. [ Links ]
Phipps, J. B., R. J O'Kennon and R.W. Lance 2003. Hawthorns and Medlars. Timber Press, Inc. Portland, OR. 139 p. [ Links ]
Rehwald, A., B. Meier, and O. Sticher. 1994. Qualitative and quantitative reversed–phase high–perfomance liquid chromatography of flavonoids in Crataegus leaves and flowers. J. Chromatogr. A 677: 25–33. [ Links ]
Rice–Evans, C. A., N. J. Miller, and G. Paganga. 1996. Structure–antioxidant activity relationships of flavonoids and phenolic acids. Free Radical Bio. Med. 20: 933–956. [ Links ]
Robards, K., P. D. Prenzler, G. Tucker, P. Swatsitang, and W. Glover. 1999. Phenolic compounds and their role in oxidative processes in fruits. Food Chem. 66: 401–436. [ Links ]
Rohr, G., B. Meier, and O. Sticher. 1999. Quantitative reversed–phase high–performance liquid chromatography of procyanidins in Crataegus leaves and flowers. J. Chromatogr. A 835: 59–65. [ Links ]
SAS (Statistical Analysis System). 1996. SAS/STAT User's Guide (Release 6.12.). SAS Institute Inc., Cary, N. C. 1028 p. [ Links ]
Schramm, D. D., and J. B. German. 1998. Potential effects of flavonoids on the etiology of vascular disease. J. Nutr. Biochem. 9: 560–566. [ Links ]
Shahat, A., P. Cos, T. De Bruyne, S. Apers, F. M. Haammouda, S. I. Isamail, S. Azzam, M. Claeys, E. Goovaerts, L. Pieters, D. V. Berghe, and A. J. Lietinck. 2002. Antiviral and antioxidant of flavonoids and proanthocyanidins from Crataegus sinaica. Planta Med. 68: 539–541. [ Links ]
Urbonaviciute, A., V. Jakstas, O. Kornysova, V. Janulis, and A. Maruska. 2006. Capillary electrophoretic analysis of flavonoids in single–styled hawthorn (Crataegus monogyna Jacq.) ethanolic extracts. J. Chromatogr. A 1112: 339–344. [ Links ]
Van Sumere, C. F., L. De Colman L, E. Everaert, D. De Keukeleire, and K. Vande Casteele. 1993. Phenolic markers in chemotaxonomy and plant cultivar recognition. In: Scarlbert, A. (ed). Polyphenolic Phenomena INRA Editions. Paris, France. pp: 257–266. [ Links ] .
Waterman, P. G., and S. Mole. 1994. Analysis of Phenolic Plant Metabolites. Methods in Ecology. Blackwell Scientific Publications. Oxford, UK. 238 p. [ Links ]
Zhang, Z., Qi Chang, M. Zhu, Y. Huang, W. K. K. Ho, and Z. Y. Chen. 2001. Characterization of antioxidants present in hawthorn fruits. J. Nutr. Biochem. 12: 144–152. [ Links ]
Zhang, Z., W. K. K. Ho, Y. Huang, A. E. Jame, L. W. Lam, and Z. Y. Chen. 2002. Hawthorn fruit is hypolipidemic in rabbits fed a high cholesterol diet. J. Nutr. 132: 5–10. [ Links ]

