










Services on Demand
Journal

Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
 Cited by SciELO
Cited by SciELO Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkAgrociencia
On-line version ISSN 2521-9766Print version ISSN 1405-3195
Agrociencia vol.44 n.7 Texcoco Oct./Nov. 2010
Recursos naturales renovables
Inducción de estructuras reproductivas en Pseudotsuga menziesii
Induction of reproductives structures in Pseudotsuga menziesii
Liliana Muñoz–Gutiérrez1, 3, J. Jesús Vargas–Hernández1*, Javier López–Upton1, Nicolás Gutiérrez–Rangel2
1 Forestal. Campus Montecillo. Colegio de Postgraduados. 56230. Montecillo, Estado de México. * Autor responsable: (vargashj@colpos.mx).
2 Fruticultura. Campus Puebla, Colegio de Postgraduados. 72130. Carretera México–Puebla La Libertad Km 125.5 Puebla.
3 INIFAP–CIRNE 78430, km 14.5. Carretera, San Luis–Matehuala, San Luis Potosí.
Recibido: Febrero, 2010.
Aprobado: Agosto, 2010.
Resumen
Pseudotsuga menziesii es una conífera muy utilizada en plantaciones de árboles de navidad en México, por lo que es necesario un abasto eficiente de germoplasma de calidad. Con el propósito de reducir el tiempo requerido para la producción de semilla en esta especie, se evaluó el efecto del ácido giberélico (AG4/7) y del anillado en el tronco en la formación de estructuras reproductivas en dos ensayos de campo con árboles juveniles. Se utilizaron cuatro dosis de AG4/7 (0, 1.27, 2.55 y 5.10 mg cm–2) aplicadas por inyección en el tronco y un año después se evaluó el porcentaje de árboles con respuesta y el número y posición de las estructuras reproductivas. Los tratamientos fueron más efectivos en formar estróbilos femeninos, con una respuesta que varió de 37.5 % en el sitio 1 a 62.5 % en el sitio 2; menos del 20 % de los árboles formaron estróbilos masculinos en los dos sitios. El desarrollo fenológico del árbol al momento de aplicar los tratamientos influyó (p≤0.01) sobre el número de estróbilos femeninos y masculinos formados. Además, el efecto del AG4/7 sobre el número de estróbilos varió entre ensayos; en el sitio 1 la dosis baja (1.27 mg cm–2) fue la más efectiva en inducir estróbilos femeninos, mientras que en el sitio 2 fue la dosis alta (5.10 mg cm 2). El anillado aumentó (p≤0.01) el número de estróbilos cuando se combinó con dosis bajas de AG4/7, pero no tuvo efecto positivo cuando se utilizaron dosis altas. El análisis de los resultados indica que la aplicación de AG4/7 en combinación con el anillado del tronco es efectiva en la inducción de floración en individuos sexualmente inmaduros de P. menziesii, lo que permite acelerar la producción de semilla en esta especie.
Palabras clave: ácido giberélico (AG4/7), anillado, estado fenológico, floración, producción de semilla.
Abstract
Pseudotsuga menziesii is a conifer widely used in Christmas tree plantations in México, which increases the need for an efficient supply of quality germoplasm. In order to reduce the time required for seed production in this species, the effect of gibberelic acid (GA4/7 ) and trunk girdling on the formation of reproductive structures was evaluated in two field trials with juvenile trees. Four dosages of GA (0,1.27, 2.55 and 5.10 mg cm–2) injected into the tree trunk were used, and one year later, the percentage of trees with a response and the number and position of reproductive structures were evaluated. The treatments were more effective on formation of female cones, with a response that varied from 37.5 % in site 1, to 62.5 % in site 2, whereas less than 20 % of the trees formed male cones in both sites. The phenological stage of the tree at the time of applying the treatments influenced (p≤0.01) the number of male and female cones formed. Also, the effect of GA on the number of cones varied with each trial; in site 1 the lowest dose (1.27 mg cm–2) was the most effective for the induction of female cones, whereas in site 2, the high dose (5.10 mg cm–2) was the most effective. Trunk girdling increased (p≤0.01) the number of cones when combined with low doses of GA4/7, but had no positive effect when high doses were used. The analysis of the results indicates that the application of GA4/7combined with trunk girdling is effective in inducing flowering in sexually immature P. menziesii trees, which helps accelerate the production of seed in this species.
Key words: gibberellic acid (GA4/7), girdling, phenological stage, flowering, seed production.
INTRODUCCIÓN
Pseudotsuga menziesii es una conífera valiosa en México para la producción de árboles de navidad, aunque suele importarse de EE.UU. y Canadá (Zavala–Chávez y Méndez–Montiel, 1996). Aunque escasa, se localiza de manera natural en México, en las Sierras Madre Occidental, Oriental y del Sur (Martínez, 1963; Debreczy y Racz, 1995), por lo que existe el potencial de establecer plantaciones comerciales. Para ello se requiere un abasto continuo de semillas de buena calidad; sin embargo, la producción de semillas en rodales naturales de esta especie es influenciada por las fluctuaciones climáticas bruscas, las deficiencias de nutrimentos, la polinización, la depredación de conos y semillas y los efectos de endogamia (Zavala–Chávez y Méndez–Montiel, 1996; Mápula–Larreta et al., 2007).
Para producir semillas abundantes, de buena calidad y en el menor tiempo posible, en especies de coníferas se han usado con éxito diversas prácticas silvícolas que inducen la diferenciación de estructuras reproductivas (Pharis et al., 1987). Estas prácticas incluyen el anillado del tronco y la aplicación de reguladores del crecimiento (Ross y Bower, 1989, 1991; Philippe et al.,, 2004; Cherry et al., 2007). El anillado consiste en un bloqueo parcial del floema para modificar el transporte de carbohidratos y de otras sustancias dentro del árbol. Ross y Bower (1989) señalan que el anillado aumenta la proporción de injertos con estróbilos femeninos (69 vs. 20 %) y masculinos (83 vs. 43 %) en P. menziesii, pero en otro estudio con la misma especie el anillado fue efectivo únicamente cuando se combinó con la aplicación de giberelinas (Ross y Pharis, 1976).
El ácido giberélico (AG) es el regulador de crecimiento más utilizado para inducir diferenciación floral en árboles (Kong y Aderkas, 2004), en especial la combinación AG4/7, en los géneros Pinus, Picea, Abies y Pseudotsuga (Pharis et al., 1980; Owens et al.,, 2001). Sin embargo, diferentes factores pueden influir en el éxito del AG4/7 como la edad de los árboles (Cherry et al., 2007), la época, la dosis y el método de aplicación (Aderkas et al., 2004), además de la combinación con anillado, poda, manejo del riego y fertilización (Ross, 1991, 1992; Philipson, 1992, 1996).
Se han desarrollado técnicas de inducción reproductiva en individuos jóvenes de origen asexual (estacas o injertos) de P. menziesii (Ross y Pharis, 1976; Ross et al., 1980; Ross, 1983), y en clones injertados de mayor edad (Ross y Bower, 1989; Cherry et al., 2007), con diferente grado de éxito. Sin embargo, no existe información del efecto de estos tratamientos en individuos juveniles de origen sexual y en materiales de México. En este estudio se evaluó el efecto de la dosis de AG4/7en combinación con el anillado del tronco sobre la diferenciación de estructuras reproductivas en árboles jóvenes de P. menziesii de origen sexual; también se analizó la efectividad de los tratamientos en ensayos separados y la influencia del estado fenológico del árbol sobre la respuesta, con la finalidad de establecer una técnica eficiente para acelerar la producción de semilla en esta especie.
MATERIALES Y MÉTODOS
Localización de los ensayos de campo
Se realizaron dos ensayos en el Conjunto Predial Forestal, en el municipio de Aquixtla, estado de Puebla (19° 43' 13" N y 97° 59' 20" O) con árboles jóvenes de P. menziesii establecidos en el bosque. El ensayo 1 se efectuó en el paraje El Manantial a 2900 m, en una plantación de árboles de navidad de 10 años de edad (plantados en este paraje en el 2000, a la edad de 2 años; la semilla para producir estos árboles se recolectó en Mineral del Chico, estado de Hidalgo), con luz solar plena y podas recurrentes de la copa. El ensayo 2 se estableció en el paraje Área semillera a 2800 m, en una plantación con árboles de 11–13 años de edad (plantados en 1996 y 1997 a la edad de un año, originados con semilla recolectada en Terrenates, estado de Tlaxcala, y Huayacocotla, estado de Veracruz), bajo el dosel de árboles adultos de Pinus fatula, sin poda de ramas durante cuatro años previos al estudio (2004–2007). En cada paraje se instaló una micro–estación meteorológica automatizada para registrar la temperatura y precipitación. En cada sitio se seleccionaron 32 árboles lo más homogéneo posible en altura (ensayo 1: 1.77±0.10 m; ensayo 2: 2.80±0.18 m), diámetro (ensayo 1: 5.4±0.3 cm; ensayo 2: 6.1 ±0.5 cm) y condiciones de la copa. En cada árbol se midió el diámetro del tronco a 50 cm del suelo, para calcular el área transversal del mismo.
Definición y aplicación de los tratamientos
En cada sitio se evaluó el anillado y la dosis de AG4/7 en un arreglo factorial y diseño experimental completamente al azar con cuatro repeticiones, con dos niveles de anillado (con y sin anillado) y cuatro dosis de AG4/7: 0, 1.27, 2.55 y 5.10 mg cm–2de área transversal del tronco, equivalente a 0, 100, 200 y 400 mg de AG4/7 en un árbol de 10 cm de diámetro del tronco. Los tratamientos se aplicaron al inicio del crecimiento del brote terminal y se determinó en cada árbol el estado fenológico de la copa (porcentaje de brotación de las yemas vegetativas en la copa del árbol). El anillado consistió en realizar con un serrote dos cortes transversales al tronco, en forma de media luna, que abarcaron cada uno de ellos aproximadamente el 60 % de la circunferencia del tronco; el primer corte se hizo a 50 cm del suelo y el segundo a una distancia del primero igual a 1.5 veces el diámetro del tronco.
El AG se aplicó en una solución de alcohol etílico al 95 %, con una concentración base de 30 mg mL–1, en un solo evento en los dos sitios. En el tronco se hicieron orificios inclinados de 2–3 cm de profundidad, a 10 cm por arriba del anillado superior y a una altura similar en los árboles no anillados. La cantidad requerida (mL) de solución, en función de la dosis y el área transversal del tronco, se aplicó con micro–pipeta; en el testigo se aplicó un volumen equivalente de alcohol etílico al 95 %; los orificios se taparon con cera para evitar pérdida de la solución.
Variables evaluadas y análisis de los datos
Al año siguiente de aplicar los tratamientos se determinó en cada ensayo el porcentaje de árboles con estróbilos femeninos y masculinos, así como su número y posición a lo largo de la copa de cada árbol, dividiéndola en cuatro secciones, desde el ápice del árbol (primer cuartil o cuartil superior) y hasta la base (cuarto cuartil o cuartil inferior). Con estos datos se calculó el número de estróbilos por árbol, el número de ramas con estróbilos y la distribución relativa de estróbilos por cuartil en la copa.
Dado que las variables en estudio no se ajustan a una distribución normal (Kuehl, 2001), se utilizaron Modelos Lineales Generalizados (MLG) que permiten trabajar con variables que se ajustan a una distribución de la familia exponencial (Poisson, Binomial, etcétera). El número de estróbilos por árbol y el número de ramas con estróbilos son variables discretas que no poseen valores negativos y tienen una distribución tipo Poisson. Para introducirlas en el MLG se realizó la transformación logarítmica de la respuesta [loge(x+0.1)] que permite contraer la distribución, normalizarla y homogeneizar las varianzas (Zar, 1996). Después del análisis, las variables fueron retransformadas a su escala original. El procedimiento GENMOD (SAS, 1999) toma directamente la variable con distribución Poisson suponiendo, entre otras cosas, que se incrementa la varianza al aumentar la media; para este modelo se utilizó la función de enlace log (SAS, 1999). Con los datos se hizo un análisis de varianza conjunto para los dos sitios, de acuerdo con el diseño experimental; aunque esto conduce a un efecto de pseudorepetición en la comparación del efecto de los sitios, ya que las repeticiones de los sitios están anidadas dentro de éstos, el modelo estadístico considera dicho anidamiento. Dado que se encontró una correlación significativa (p≤0.05) entre el estado fenológico de la copa y la diferenciación de estructuras reproductivas, para remover el efecto de este factor se incluyó el estado fenológico de los árboles como covariable en el modelo. En las variables donde se encontró un efecto significativo asociado a un factor, se realizó una comparación de medias con la prueba de Tukey (p≤0.05).
Para comparar el porcentaje de árboles con respuesta y la distribución relativa de los estróbilos en los cuartiles de la copa entre los niveles de cada factor por separado, se utilizó la prueba de ji–cuadrada (χ2), mediante tablas de contingencia (Infante y Zárate, 2000).
RESULTADOS Y DISCUSIÓN
Proporción de árboles con estructuras reproductivas
En los dos ensayos de campo se observó la formación de estructuras reproductivas, pero mientras en el sitio 1 el 37.5 % de los árboles formaron estróbilos femeninos, en el sitio 2 la proporción fue 62.5 % (Cuadro 1). La frecuencia de árboles con estróbilos masculinos fue notoriamente menor en ambos sitios, variando de 6 % en el sitio 2 a 12.5 % en el sitio 1. El anillado al tronco no afectó significativamente (p>0.05) la proporción de árboles que formaron estructuras reproductivas en ninguno de los dos sitios. En cambio, la dosis de AG4/7 sí tuvo un efecto importante y significativo (p≤0.05); en el sitio 1 la frecuencia de árboles con estróbilos femeninos varió de 0 % en el testigo hasta 75 % al aplicar 1.27 mg cm–2 de AG4/7 y la de estróbilos masculinos de 0 a 38 % en los mismos tratamientos (Cuadro 1).
Número de estructuras reproductivas por árbol
El número de estróbilos por árbol fue afectado significativamente (p≤0.05) por dos de los factores evaluados (Cuadro 2), aunque los efectos fueron más notorios en los femeninos. Por ejemplo, el estado fenológico del árbol influyó en el número de estróbilos femeninos por árbol y el número de ramas con estróbilos (p≤0.001), pero no sobre los estróbilos masculinos (Cuadro 2). Los factores sitio y anillado por sí solos no tuvieron efectos significativos (p>0.05), pero cada uno de ellos en interacción con la dosis de AG4/7 sí tuvo un efecto importante sobre la inducción del desarrollo de estructuras reproductivas. Además, la dosis de AG4/7 influyó (p≤ 0.001) en el número de estróbilos femeninos por árbol y el número de ramas con estróbilos femeninos, pero no sobre el número de estróbilos masculinos (Cuadro 2).
El número de estróbilos tendió a aumentar en los árboles que se encontraban más avanzados en el desarrollo de los brotes (Figura 1a); el efecto más notorio fue en árboles del sitio 2, donde el mayor número de estróbilos femeninos se presentó en árboles con más del 90 % de brotación de las yemas vegetativas al momento de aplicar los tratamientos. La inducción de estróbilos masculinos en el sitio 1 ocurrió en árboles que tenían entre 30 y 70 % de brotación de yemas (Figura 1b), mientras que en el sitio 2 no se observó efecto del estado fenológico sobre esta variable.
El número de estróbilos masculinos formados por árbol fue relativamente bajo (alrededor de 0.5) y sin diferencias significativas entre dosis de AG4/7 ; sin embargo, el número de estróbilos femeninos por árbol y el número de ramas con estróbilos aumentó con la dosis de AG4/7 (Cuadro 3). Aunque no hubo diferencias estadísticas entre la dosis baja y alta de AG4/7(1.27 vs. 5.10 mg cm–2), con la última se formó el triple de estróbilos femeninos (4.23 vs. 14.54). Además, el efecto de la dosis de AG4/7 sobre el número de estructuras reproductivas fue diferente de un ensayo a otro (Cuadro 3). En el sitio 1, la dosis baja de AG4/7(1.27 mg cm–2) fue la más efectiva para promover la formación de estróbilos femeninos y masculinos (9.20 y 1.57), mientras que en el sitio 2 se requirió una dosis alta (5.10 mg cm–2) para obtener el mayor número de estróbilos (25.25 y 1.61).
El anillado del tronco con dosis bajas de AG4/7 aumentó la formación de estróbilos femeninos por árbol, pero combinado con dosis altas tuvo un efecto negativo, en comparación con árboles sin anillar (Figura 2). Así, árboles anillados y con 1.27–2.55 mg cm–2 de AG4/7 aumentaron de 2.1 a 8.4 el número de estróbilos, mientras que el anillado redujo el número de estróbilos de 16.9 a 12.5 con la dosis de 5.10 mg cm–2.
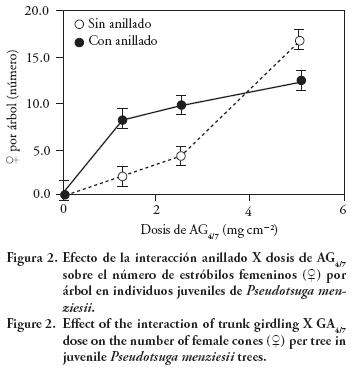
Posición de las estructuras reproductivas en la copa del árbol
En ambos ensayos, la mayoría de los estróbilos femeninos se encontró en los dos primeros cuartiles (parte superior de la copa del árbol), y ninguno en el cuarto cuartil (parte inferior de la copa, Cuadro 4). En ambos casos el anillado modificó la distribución de los estróbilos, aumentando su proporción hacia las ramas intermedias de la copa, en el segundo (sitio 1) y tercer (sitio 2) cuartil, con respecto a los árboles no anillados (Cuadro 4). La dosis de AG4/7 también afectó la distribución de los estróbilos femeninos en la copa del árbol, pero el efecto fue distinto entre sitios; en el sitio 1, la dosis elevada propició una mayor formación de estróbilos en el primer cuartil de la copa, mientras que en el sitio 2 el efecto fue en el segundo (Cuadro 4).
Efectividad de la inducción de estructuras reproductivas
Se logró inducir la formación de estróbilos masculinos y femeninos en árboles jóvenes de origen sexual de P. menziesii de manera consistente en dos ensayos realizados bajo diferentes condiciones ambientales y silvícolas. En la mayoría de los estudios publicados acerca de esta especie se han usado material de origen asexual, generalmente injertos obtenidos de individuos sexualmente maduros (Philippe et al., 2004) y con un historial de producción conocido. Aunque la proporción de árboles que formó estróbilos femeninos y masculinos fue baja (Cuadro 1), el estudio muestra que con la aplicación oportuna de una dosis apropiada de AG4/7 con o sin anillado del tronco, es posible acelerar la producción de semilla en individuos sexualmente inmaduros de P. menziesii.
La efectividad de la inducción varió entre los ensayos; la proporción de árboles que formaron estróbilos femeninos fue mayor en el sitio 2 (62.5 y 37.5 %) y la proporción con estróbilos masculinos fue mayor en el sitio 1(12.5 vs. 6.0 %); el número de estróbilos femeninos fue mayor en el sitio 2 y el de estróbilos masculinos similar en los dos sitios (Cuadro 3). Estas diferencias se pueden deber a diversos factores, incluyendo la edad (los árboles del sitio 2 eran mayores que los del sitio 1), el microambiente [en el sitio 2 la temperatura media de febrero a mayo de 2008 fue ligeramente mayor (11.4 vs. 10.2 °C)], el manejo previo de los árboles (los árboles del sitio 1 recibieron podas recurrentes, mientras que los del sitio 2 no fueron podados en los últimos 4 años), o el estado fenológico de los árboles (los árboles del sitio 2 se encontraban más avanzados fenológicamente que los del sitio 1, como resultado del microambiente y la procedencia del germoplasma).
El estado fenológico de los brotes influyó sobre la efectividad de los tratamientos para inducir estructuras reproductivas; la formación de estróbilos femeninos fue más común en los árboles con mayor avance en la etapa fenológica estudiada, mientras que los masculinos ocurrieron en árboles menos avanzados (Figura 1). Al parecer, la diferenciación de las estructuras femeninas y masculinas en esta especie ocurre en diferentes etapas de desarrollo de los brotes, como se ha encontrado en otras especies; en Picea engelmannii, el estrés por sequía y calor aplicado en etapas tempranas del desarrollo de los brotes favorece la formación de estróbilos masculinos, mientras que en etapas más avanzadas favorece a los femeninos (Ross, 1985).
Aunque Ross (1983) señala que las aplicaciones de AG4/7 en P. menziesii al inicio del crecimiento del brote son más efectivas para inducir los dos tipos de estróbilos, otros autores han mostrado que el estado fenológico del brote influye sobre la proporción de sexos en las estructuras diferenciadas (Harrison y Owens, 1994). Ho y Eng (1995) encontraron que en Pinus strobus la producción de estróbilos masculinos aumenta al inicio de la etapa de crecimiento rápido del brote, mientras que en la etapa final de crecimiento no hay efecto de los tratamientos.
Efecto de la dosis de AG4/7
Los resultados corroboraron la efectividad del AG4/7 para inducir la diferenciación de estructuras reproductivas; sin embargo, la dosis más adecuada varió de un sitio a otro y con respecto al sexo de los estróbilos (Cuadro 3). Greenwood (1987) menciona que el AG4/7 es más efectivo en promover la producción de estróbilos femeninos, aunque las diferencias entre ambos sexos también podrían deberse al momento de aplicación de la hormona en relación con la fenología del brote (Harrison y Owens, 1994) o a las dosis utilizadas. Cherry et al. (2007) lograron promover mayor número de estróbilos masculinos que femeninos con dosis bajas de AG4/7 (0.338 y 1.35 mg cm2). En la familia Cupressaceae también se ha encontrado que las dosis bajas de AG4/7 promueven los estróbilos masculinos (Pharis y Kuo, 1977).
La formación de estróbilos femeninos fue mayor en el sitio 2 al utilizar dosis altas de AG4/7 mientras que en el sitio 1 la dosis baja tuvo mayor efecto. La edad y el manejo previo de los árboles en cada sitio pudieron influir en estos resultados, al modificar su estado juvenil o fisiológico; los árboles del sitio 2 representan una mayor edad fisiológica y se ha encontrado que la respuesta a la estimulación floral incrementa con la edad de los árboles (Cherry et al., 2007); estos mismos autores sugieren aplicar dosis bajas en árboles jóvenes en combinación con el anillado del tronco, lo cual coincide con Ross (1983), quien con dosis bajas (100 mg L–1) obtuvo una mayor formación de estróbilos femeninos en Pseudotsuga menziesii que con las dosis altas (800 mg L–1) de AG4/7.
Dosis elevadas de AG4/7 pueden tener efectos adversos sobre los árboles tratados, especialmente en los jóvenes; en algunos árboles se observó una ligera declinación en las ramas de la parte baja del árbol, posiblemente por efectos de fito–toxicidad, como lo señalan Ross y Bower (1991). Además, las dosis elevadas de AG4/7 pueden tener efectos indirectos que afectan la salud y el vigor de los árboles, derivados de la energía utilizada en la producción de estróbilos (Bonnet–Masimbert, 1987; Cherry et al., 2007).
En condiciones naturales las especies de coníferas forman los estróbilos femeninos en la parte superior de la copa y los masculinos en la parte media y baja (Dick et al., 1985). Este mismo patrón se ha observado en árboles jóvenes o injertos fisiológicamente maduros al aplicar tratamientos para estimular la floración (Philipson, 1985; Ross, 1990). Sin embargo, en el presente estudio se encontró que la dosis de AG4/7 afectó la distribución de los estróbilos femeninos en ambos sitios (Cuadro 4). En el sitio 1 la dosis baja aumentó la proporción de estróbilos en el segundo cuartil de la copa y la dosis más alta la aumentó en el primer cuartil; en el sitio 2, aunque se mantiene una mayor proporción de estróbilos en la parte alta de la copa, también aumenta la formación de estróbilos en el tercer cuartil. Dado que los árboles del sitio 2 tienen mayor altura que los del sitio 1, las diferencias en la posición de los estróbilos pueden deberse a la distancia que hay del punto de aplicación de la hormona al ápice de la copa.
Efecto del anillado del tronco
El anillado en el tronco por si solo no tuvo el efecto esperado, ya que en otros estudios se ha mostrado que el anillado es una práctica efectiva en la formación de estróbilos en Larix y Pseudotsuga (Ross y Bower, 1989; Philipson, 1996; Cherry et al., 2007). Sin embargo, cuando se utilizaron dosis bajas de AG4/7, el anillado aumentó la producción de estróbilos femeninos, indicando que influyó sobre la efectividad del tratamiento hormonal. En Picea sitchensis el AG4/7 combinado con calor y sequía estimuló la formación de ambos tipos de estróbilos y el anillado fue más efectivo cuando se combinó con calor (Philipson, 1992).
El anillado también modificó la posición de los estróbilos femeninos en la copa del árbol, aumentando su proporción en la parte intermedia (segundo y tercer cuartil) de la copa (Cuadro 4). Este efecto coincide con la acción del AG4/7, indicando que el bloqueo parcial en el floema ocasionó una mayor disponibilidad de carbohidratos y de otras substancias promotoras del crecimiento en las ramas bajas de la copa más cercanas al punto de bloqueo, según lo indicado por Ross y Bower (1989).
Implicaciones para la producción de semillas
Los tratamientos aplicados son efectivos y prácticos para inducir estróbilos femeninos y masculinos en árboles juveniles de P. menziesii; sin embargo, se debe promover un mayor equilibrio en la formación de estructuras de ambos sexos. La época de aplicación fue apropiada para la formación de estróbilos femeninos, pero es necesario explorar la aplicación en etapas fenológicas más tempranas para aumentar la formación de estróbilos masculinos y la proporción de árboles con respuesta favorable. Es evidente que las diferencias en la respuesta de los árboles se deben a múltiples factores relacionados con edad, vigor y estado fisiológico de cada uno de ellos, por lo que es importante asegurar una mayor homogeneidad en la plantación para aumentar la capacidad de producción de semilla.
La producción de polen es a menudo un factor limitante en árboles juveniles; sin embargo, en la producción operativa de semilla el problema se puede resolver en forma práctica mediante la recolecta de polen de árboles maduros y la polinización artificial de los estróbilos femeninos inducidos. Aunque la mayoría de los estróbilos femeninos formados se desarrollaron de manera normal, posiblemente por el efecto de las giberelinas, no formaron semillas viables por la falta de polen.
CONCLUSIONES
En los dos ensayos se logró promover la formación de estructuras reproductivas femeninas y masculinas en árboles juveniles de origen sexual con la aplicación de AG4/7 inyectado al tronco; el nivel de éxito varió en cada caso debido a las condiciones del sitio, la edad de los árboles, el manejo previo y el desarrollo fenológico. El anillado fue efectivo cuando se combinó con dosis bajas de AG4/7 (≤2.55 mg cm–2), pero combinado con dosis altas tuvo un efecto negativo. Estos tratamientos pueden ser útiles en programas operativos de producción de semilla de P. menziesii.
AGRADECIMIENTOS
Los autores expresan su agradecimiento al Ing. León Jorge Castaños Martínez y al Ing. Salvador Castro Zavala, propietarios del Conjunto Predial Forestal, por todas las facilidades y el apoyo logístico proporcionado durante la realización del estudio.
LITERATURA CITADA
Aderkas, P., L. Kong, S. Abrams, I. Zaharia, S. Owens, and B. Porten 2004. Flower induction methods for lodgepole pine and Douglas–fir. Centre for Forest Biology, University of Victoria. 59 p. [ Links ]
Bonnet–Masimbert, M. 1987. Preliminary results on gibberellins induction of flowering of seedlings and cuttings of Norway spruce indicate some carry–over effects. For. Ecol. Manage. 19:163–171. [ Links ]
Cherry, M. L., T. S. Anekonda, M. J. Albrecht, and G. T. Howe. 2007. Flower stimulation in young miniaturized seed orchards of Douglas–fir (Pseudotsuga menziesii). Can. J. For. Res. 37:1–10. [ Links ]
Debreczy, Z., and I. Racz. 1995. New species and varieties of conifers from Mexico. Phytologia 78:217–243. [ Links ]
Dick, J. M., R. I. Smith, and K. A. Longman. 1985. Effect of bark–ringing and gibberellins on the number and distribution of cones in Sitka spruce (Picea sitchensis). Acta Univ. Agrie. Fac. Agron. (Brno). 3:659–663. [ Links ]
Greenwood, M. S. 1987. Rejuvenation of forest trees. Plant Growth Regulation 6:1–12. [ Links ]
Harrison, D. L. S., and J. N. Owens. 1994. Gibberellins A4/7 enhanced flowering in Tsuga heterophylla. II. Apical development and shoot formation. Int. J. Plant Sci. 155:302–312. [ Links ]
Ho, R. H., and K. Eng. 1995. Promotion of cone production on field–grown eastern white pine grafts by gibberellin A4/7 application. For. Ecol. Manage. 75:11–16. [ Links ]
Infante G., S., y G. P. Zárate L. 2000. Métodos Estadísticos; un Enfoque Interdisciplinario. Ed. Trillas, México. 643 p. [ Links ]
Kong, L., and P. Aderkas. 2004. Plant growth regulators and cone induction in Pinaceae. University of Victoria, Victoria, BC. Canada. 27 p. [ Links ]
Kuehl, R. 2001. Diseño de Experimentos. Principios Estadísticos para el Diseño y Análisis de Investigaciones. Thomson Learning. México. 666 p. [ Links ]
Mápula–Larreta, M., J. López–Upton, J. J. Vargas–Hernández and A. Hernández–Livera. 2007. Reproductive indicators in natural populations of Douglas–fir in Mexico. Biodiversity and Conservation 16:727–742. [ Links ]
Martínez, M. 1963. Las Pináceas Mexicanas. Universidad Nacional Autónoma de México. 3ª Ed. México, pp: 27–74. [ Links ]
Owens, J. N., L. M. Chandler, J. S. Bennett, and T. J. Crowder. 2001. Cone enhancement in Abies amabilis using GA4/7 , fertilizer, girdling and tenting. For. Ecol. Manage. 154: 227–236. [ Links ]
Pharis, R. P., and C. G. Kuo. 1977. Physiology of gibberellins in conifers. Can. J. For. Res. 7:299–325. [ Links ]
Pharis, R. P., S. D. Ross, and E. E. McMullan. 1980. Promotion of flowering in the Pinaceae by gibberellins. III. Seedlings of Douglas–fir. Physiol. Plant. 50:119–126. [ Links ]
Pharis, R. P., J. E. Weber, and S. D. Ross. 1987. The promotion of flowering in forest trees by gibberellin A4+7 and cultural treatments: a review of the possible mechanisms. For. Ecol. Manage. 19:65–84. [ Links ]
Philippe, G., S. J. Lee, G. Schute, and B. Heois. 2004. Flower stimulation is cost–effective in Douglas–fir seed orchards. Forestry 77:279–286. [ Links ]
Philipson, J. J. 1985. The effect of top pruning, girdling, and gibberellins A4/7 application on the production and distribution of pollen and seed cones in Sitka spruce. Can. J. For. Res. 15:1125–1128. [ Links ]
Philipson, J. J. 1992. Optimal conditions for inducing coning of container–grown Picea sitchensis grafts: effects of applying different quantities of GA4/7 , timing and duration of heat and drought treatment, and girdling. For. Ecol. Manage. 53:39–52. [ Links ]
Philipson, J. J. 1996. Effects of girdling and gibberellins A4/7 on flowering of European and Japanese larch grafts in an outdoor clone bank. Can. J. For. Res. 26:355–359. [ Links ]
Ross, S. D. 1983. Enhancement of shoot elongation in Douglas–fir by gibberellins A4/7 and its relation to the hormonal promotion of flowering. Can. J. For. Res. 13:986–994. [ Links ]
Ross, S. D. 1985. Promotion of flowering in potted Picea engelmannii (Perry) grafts: effects of heat, drought, giberellin A4/7 and their timing. Can. J. For. Res. 15:618–624. [ Links ]
Ross, S. D. 1990. Control sex expression in potted Picea engelmannii grafts by gibberellins A4/7 and the auxin, naphthaleneacetic acid. Can. J. For. Res. 20:875–879. [ Links ]
Ross, S. D. 1991. Promotion of flowering in western larch by girdling and gibberellins A4/7 and recommendations for selection and treatment of seed trees. Research Note No. 105, B.C. Ministry of Forests, Research Laboratory. 13 p. [ Links ]
Ross, S. D. 1992. Promotion of flowering in Engelmann–white spruce seed orchards by GA4/7 stem injection: effects of site, clonal fecundity, girdling and NAA, and treatment carryover effects. For. Ecol. Manage. 50:43–55. [ Links ]
Ross, S. D., and R. C. Bower. 1989. Cost–effective promotion of flowering in a Douglas–fir seed orchard by girdling and pulsed stem injection of gibberellin A4/7. Silvae Genet. 38:189–195. [ Links ]
Ross, S. D., and R. C. Bower. 1991. Promotion of seed production in Douglas–fir grafts by girdling + gibberellin A4/7 stem injection, and effect of retreatment. New Forests 5:23–34. [ Links ]
Ross, S. D., and R. P. Pharis. 1976. Promotion of flowering in the Pinaceae by gibberellins. I. Sexually mature, non–flowering grafts of Douglas–fir. Physiol. Plant. 36:182–186. [ Links ]
Ross, S. D., R. P. Pharis, and J. C. Heaman. 1980. Promotion of cone and seed production in grafted and seedling Douglas–fir seed orchards by applications of gibberellins A4/7 mixture. Can. J. For. Res. 10:464–469. [ Links ]
SAS Institute Inc. 1999. SAS/STAT User's Guide Version 8, Cary, NC: SAS Institute Inc. Chapter 29. The GENMOD Procedure, pp: 1363–1464. [ Links ]
Zar, J. 1996. Biostatistical Analysis. Prentice–Hall. New Jersey. EE.UU. 662 p. [ Links ]
Zavala–Chávez, F. y J. T.Méndez–Montiel. 1996. Factores que afectan la producción de semillas en Pseudotsuga macrolepis Flous. en el Estado de Hidalgo, México. Acta Bot. Mex. 36:1–13. [ Links ]

