










Services on Demand
Journal

Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
 Cited by SciELO
Cited by SciELO Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkAgrociencia
On-line version ISSN 2521-9766Print version ISSN 1405-3195
Agrociencia vol.42 n.5 Texcoco Jul./Aug. 2008
Recursos naturales renovables
Efecto del tamaño y color de la semilla en la germinación de Cecropia obtusifolia Bertol (Cecropiaceae)
Effect of seed size and color on germination of Cecropia obtusifolia Bertol (Cecropiaceae)
Gabriela Tenorio–Galindo, Dante A. Rodríguez–Trejo* y Georgina López–Ríos
División de Ciencias Forestales. Universidad Autónoma Chapingo. 56230. Chapingo, Estado de México *Autor responsable: (dantearturo@yahoo.com)
Recibido: Mayo, 2007.
Aprobado: Abril, 2008.
Resumen
Se estudió el efecto de tamaño y color de la semilla de Cecropia obtusifolia Bertol en su germinación. Hubo dos tamaños: semillas chicas (1.3 mm longitud) y grandes (1.7 mm longitud); tres colores de la cubierta seminal: negro, café claro y café oscuro; dos temperaturas en cámara de ambiente controlado: 30 °C en el día y 25 °C en la noche, 25 °C en el día y 20 °C en la noche. El diseño experimental fue de bloques completos al azar, con los bloques anidados dentro del factor temperatura. El color y el tamaño influyeron significativamente (p<0.05) en la capacidad germinativa, al igual que las interacciones temperatura x color, tamaño x color, y la interacción triple: temperatura x tamaño x color. Las semillas grandes germinaron casi el doble que las pequeñas; las de color café oscuro germinaron bien en ambos regímenes de temperatura. Con temperatura alta la mayor germinación fue en semillas grandes color café claro; con temperatura baja, las semillas grandes color café oscuro germinaron más. Así, las variantes en tamaño y color de las semillas les permiten germinar en distintas condiciones ambientales.
Palabras clave: Chancarro, ecofisiología de semillas, guarumbo.
Abstract
The effect of seed size and color on germination was studied in Cecropia obusifolia Bertol. Tested were two seed sizes: small (1.3 mm long) and large (1.7 mm long); three seed coat colors: black, light brown and dark brown; two day and nighttime temperatures in a controlled environment chamber: 30 °C during the day and 25 °C at night and 25 °C during the day and 20 °C at night. The experimental design was randomized complete blocks, with blocks nested within the factor temperature. Color and size had a significant (p<0.05) effect on germinative capacity, as did the interactions temperature x color and size x color, and the triple interaction temperature x size x color. The number of large seeds that germinated was almost double that of small seeds. Dark brown seeds germinated well under both temperature regimes. At the higher temperatures, the highest rate of germination was that of large light brown seeds; at the lower temperatures, more large dark brown seeds germinated. Thus, the varying seed sizes and colors allow them to germinate in different environmental conditions.
Key words: Chancarro, seed ecophysiology, guarumbo.
INTRODUCCIÓN
El guarumbo, Cecropia obtusifolia Bertol, se distribuye a lo largo de las costas del Pacífico (desde el sur de Sinaloa y Durango hasta Chiapas) y el Atlántico (del sur de Tamaulipas y San Luis Potosí hasta Chiapas) en México, y en las selvas de Centroamérica; se encuentra de 0 a 800 m (Pennington y Sarukhán, 2005). Típicamente coloniza áreas perturbadas en los trópicos (Álvarez y Martínez, 1992; Álvarez, 1997), por lo que es una especie pionera en la sucesión ecológica.
Un árbol de guarumbo puede producir semillas casi todo el año; la tasa de fecundidad anual varía de 1.4x104 a 1.4x107 semillas por árbol, y aumenta con el diámetro y el número de ramas (Álvarez y Martínez, 1992). Las semillas son fotoblásticas, sensibles a cambios en la calidad de la luz (proporción de rojo y rojo lejano) y a su intensidad; la mayor germinación es con luz plena, semejante a la de claros grandes en la selva (Vázquez y Smith, 1982).
El tamaño de las semillas en una especie varía hasta en 50% (Leishman et al., 2000), lo cual puede tener repercusiones importantes. En condiciones naturales y simuladas, las semillas grandes germinan más y con mayor vigor que las pequeñas (van Molken et al., 2005). Además, el color de la simiente puede influenciar la capacidad germinativa, como en Atriplex inflata F. Muell., donde las semillas claras no requieren tratamientos pregerminativos, a diferencia de las oscuras (Baskin y Baskin, 2001).
Por tanto, el objetivo para el presente trabajo fue evaluar el efecto de temperaturas en la germinación y su relación con el tamaño y color de las semillas. La hipótesis fue que la germinación de las semillas con distinto tamaño y color varía según la temperatura. Además se hizo un análisis físico para describir las características principales de la semilla en esta especie.
METODOLOGÍA
Las semillas se recolectaron en noviembre de 2004 en una zona de transición entre selva alta perennifolia y bosque mesófilo de montaña en la localidad de San Antonio, Mpio. de Huehuetla, Hidalgo. Los frutos de 10 árboles fueron procesados en laboratorio, obteniéndose 0.6 kg de semilla.
Se hizo el análisis de semillas con las normas de AOSA (Bonner et al., 1994), excepto por el número de semillas en las pruebas de germinación, debido a limitaciones en la cantidad y espacio requerido para todos los tratamientos. El peso de las semillas se calculó mediante el conteo y obtención de peso de seis muestras de 100 semillas cada una. La viabilidad se determinó con la prueba de tetrazolio, colocando 100 semillas en una solución de cloruro de 2,3,5, trifenil tetrazolio al 1%, a 30 °C por 24 h.
Las pruebas de germinación se hicieron en una cámara de ambiente controlado marca Conviron®. Se consideró germinada una semilla cuando la radícula duplicaba su longitud. Se estableció un experimento en bloques completos al azar, con los bloques anidados dentro del factor temperatura, para analizar su efecto y de el tamaño de semilla y color. El factor temperatura tuvo dos regímenes basados en fotoperiodo de 12 h: 30/25 °C: 30 °C (luz) y 25 °C (oscuridad); 25/20 °C: 25 °C (luz) y 20 °C (oscuridad). Se usó luz incandescente y fluorescente (RFA = 124.7 µmol m-2 s-1), medida con un sistema portátil LCi (Bio Scientific Ltd.). Se empleó luz directa en todos los tratamientos con base en el carácter fotoblástico para la semilla de la especie estudiada con mayor germinación bajo luz plena (Vázquez y Smith, 1982). Para el factor tamaño hubo semillas chicas (< 1.5 mm; media 1.3 mm) y semillas grandes (>1.5 mm; media 1.7 mm). Para el factor color de las semillas hubo café claro, café oscuro y negro. Se generaron 12 tratamientos, con cinco repeticiones (bloques) y 20 semillas por unidad experimental. El ensayo de germinación se realizó en cajas de Petri con papel filtro. Se regó con una solución de Captán (3 g L–1) con la frecuencia necesaria para mantener saturado el sustrato. La germinación se registró cada día hasta un periodo final de 7 d sin germinación alguna, 30 d después de la siembra.
Para el análisis estadístico se usó un modelo mixto:
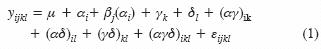
donde: µ = media general; αi = efecto del i–ésimo nivel del factor temperatura; βj(αi) = efecto del j–ésimo bloque anidado dentro del i–ésimo nivel del factor temperatura; γk = efecto del k–ésimo nivel del factor tamaño de semilla; δl = efecto del l–ésimo nivel del factor color de la semilla; εijkl = error experimental. Las combinaciones de letras representan interacciones. Los tratamientos y sus interacciones se consideraron efectos fijos; los bloques efectos aleatorios.
Para el análisis estadístico la variable germinación fue transformada mediante la función arco seno. Se usó el procedimiento Mixed (SAS, v. 8.00 para microcomputadoras) y se aplicó la prueba de diferencia mínima significativa (p<0.05).
RESULTADOS
Análisis de semillas
Con base en el peso de las muestras se calculó un peso de mil semillas de 0.71 g, así como 1 408 451 semillas kg–1. Su contenido de humedad, con base en peso seco, fue 12%. La pureza fue 99%; las impurezas fueron semillas rotas, restos del fruto y ramillas principalmente. La viabilidad de la semilla fue 53.5%. La semilla de C. obtusifolia es oblonga–ovoide y su longitud de 1.1 a 2 mm. En el presente trabajo la variación en la longitud de la semilla fue el doble que la reportada por Leishman et al. (2000).
Germinación
Hubo un efecto significativo del tamaño de la semilla y del color, así como de las interacciones temperatura x color, tamaño x color, y temperatura x tamaño x color (p<0.05), sobre la capacidad germinativa de las semillas (Cuadro 1).
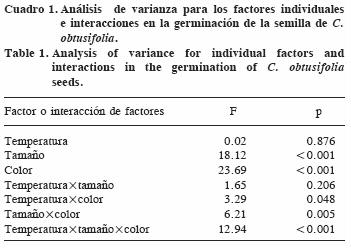
No hubo efecto significativo de la temperatura sola, y las semillas germinaron con valores semejantes en ambos regímenes de temperatura: 33.5% en promedio para 30/25 °C y 31.3% para 25/20 °C. Las semillas grandes germinaron aproximadamente el doble que las semillas pequeñas, mientras que las semillas café oscuro tuvieron mayor germinación (Figura 1). En el régimen de temperaturas más alto no hubo diferencia entre las semillas café oscuro y café claro, pero ambas superaron a las negras. En el régimen de temperatura más bajo, las semillas café oscuro superan a las café claro y negras, estas dos últimas sin diferencias entre sí (Figura 2).
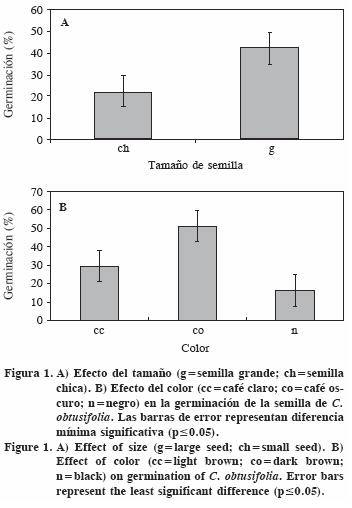
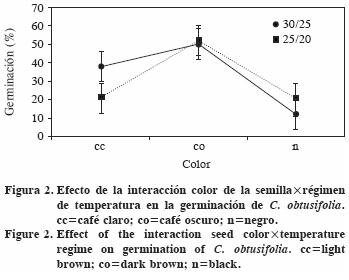
En la interacción entre tamaño y color de la semilla (Figura 3) destaca que las semillas grandes y cafés (en cualquier tonalidad) y las semillas chicas café oscuro, superan a las semillas negras de cualquier tamaño y a las chicas café claro.
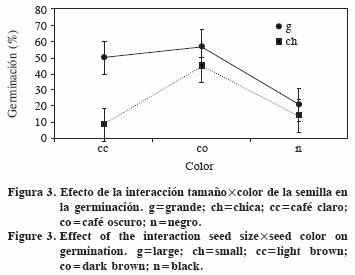
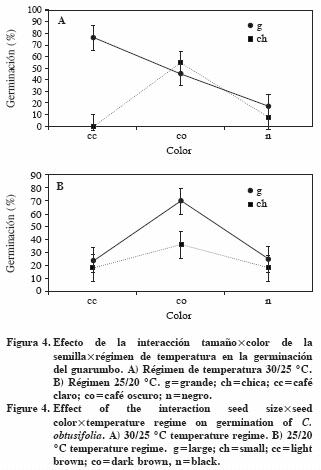
La interacción entre los tres factores muestra que en el ambiente más cálido las semillas grandes café claro germinan más, en tanto que para el ambiente más fresco las semillas grandes café oscuro tuvieron mayor germinación (Figura 4). En el ambiente cálido, después de las semillas grandes café claro, las café oscuro de cualquier tamaño superaron al resto. En la temperatura alta, las semillas chicas café claro o negro germinaron menos que las del mismo tamaño y color café oscuro.
DISCUSIÓN
La variación en tamaños, colores y formas de las simientes es controlada por el ambiente, la genética y su interacción, interviniendo en el primer caso factores como nutrientes, luz, sombra, época del año, defoliación, temperatura y humedad, así como la posición en el fruto. Tal variación se relaciona con diferencias en los requerimientos para germinar y en el rompimiento de la latencia (Baskin y Baskin, 2001).
Tamaño de la semilla
En Raphanus raphanistrum L. hay variación en el tamaño de las semillas en la silicua. Las semillas distales son más pequeñas que aquéllas en otras posiciones porque los óvulos distales son fertilizados después de los de porciones centrales; así, las semillas distales no tienen mucho tiempo para desarrollarse como las otras (Mazer et al., 1986). Garrido et al. (2005) señalan que la variación en el tamaño de semillas de Helloborus foetidus L. obedece a diferencias en la adjudicación de recursos. La variación en el tamaño de la semilla de C. obtusifolia se puede deber a la posición de la semilla en la infrutescencia. Durante su formación los aquenios pueden competir por recursos entre sí y tener diferentes tamaños de fruto y semilla, como señala Méndez (1997) para Arum italicum L., cuyos frutos son bayas agrupadas en racimos.
La masa de la semilla es un factor biológico importante que afecta la germinación, el alargamiento y crecimiento de las plántulas (Cordazzo, 2002). El tamaño también está positivamente relacionado con el tamaño inicial de la planta (Moegenburg, 1996). Así, una semilla grande resultará en una plántula más grande y con mayores probabilidades de sobrevivir que una pequeña. Según Leishmann et al. (2000), la supervivencia de plántulas está directamente relacionada con el tamaño de la semilla. En girasol (Helianthus annuus L.), las semillas grandes (4 mm) tuvieron mayor capacidad germinativa que las semillas de menores tamaños (Krishnaveni y Sivasubramanian, 2001). Una tendencia similar fue encontrada por van Mölken et al. (2005) para Tragopogon pratensis sbsp. pratensis. Hendrix (1984) indica que las semillas grandes frecuentemente tienen mayor porcentaje de germinación o emergencia que las semillas pequeñas, lo cual ocurrió en nuestro trabajo. Cordazzo (2002) encontró resultados similares al probar el efecto de la masa de la semilla en la germinación y el crecimiento de plántulas de Blutaparon portulacoides (St. Hill) Mears, Panicum racemosum (P. Beauv.) Sprengel y Spartina ciliata Brongn., con efectos significativos en todas las especies. La capacidad de alargamiento de las plántulas fue mayor en las provenientes de semillas grandes. Esta ventaja de tamaño inicial de la plántula puede persistir hasta la madurez (Weis, 1982) o perderse con el tiempo (Zimmerman y Weis, 1983).
Un tamaño grande en semillas tiene otras ventajas, como la mayor capacidad para emerger desde mayores profundidades del suelo, en comparación con las simientes pequeñas (Radford, 1977) o una mayor concentración de nitrógeno, como en la semilla de Pericopsis elata Harms van Meeuwen, árbol del bosque tropical subcaducifolio de Ghana (Burslem y Miller, 2001). Con deficiencia de nutrientes, una plántula de una semilla grande tiene más probabilidad de sobrevivir que una plántula de una semilla pequeña porque tiene más reservas (Jurado y Westoby, 1992). Sin embargo, cuando las plantas crecen en ambientes limitativos tienden a mostrar mayor variación en el tamaño de las simientes que producen, como Halpern (2005) señala para Lupinus perennis L. Otro factor importante es la herbivoría, pues las plántulas pueden sufrir daños por remoción de hojas en sus primeras etapas de vida. En C. obtusifolia la colonización por hormigas que defienden contra depredadores se da hasta que las plantas tienen una altura de unos 80 cm. La capacidad de una plántula para sobrevivir cuando se ha removido parte de su follaje, está positivamente asociada con el tamaño de la semilla (Armstrong y Westoby, 1993; Bonfil, 1998).
Color de la semilla
El color de la semilla está correlacionado positivamente con restricción a la germinación, debido a los componentes fenólicos en la cubierta seminal (Debeajun et al., 2002). En algunas especies como Sinapis arvensis L., se reduce la latencia en semillas rojas, comparadas con semillas negras (Duran y Retamal, 1989). Las semillas blancas de leguminosas se embeben más rápidamente que las de otro color y germinan primero. En el trigo, la latencia está asociada con una cubierta seminal roja, mientras que en las semillas blancas no hay latencia o es débil (Mares, 1994). Las semillas oscuras del mijo (Panicum miliaceum L.) tienen la cubierta de la semilla más pesada, se embeben y germinan más lentamente; por tanto, persisten más tiempo en el suelo que las semillas claras (Khan et al., 1996).
La variación en el color de la semilla se puede deber al contenido y distribución de pigmentos, por ejemplo antocianinas, como en la soja (Glycine max (L.) Merr.) (Todd y Vadkin, 1996). En semillas de frijol el color de la cubierta está determinado por la presencia y cantidad de glicósidos, flavonoides, antocianinas y taninos condensados. Estos últimos están en la cubierta de la semilla de C. obtusa Trécul, C. palmata (Willd.) Kuntze y C. sciadophylla Mart. (Lobova et al., 2003). La cantidad y distribución de flavonoides puede afectar el color de la cubierta de C. obtusifolia como sucede con otras especies. Así, las semillas negras pueden tener mayores concentraciones de estos agentes, menguando la germinación, independientemente de su tamaño. La variación en color de la cubierta de la semilla se interpreta como una estrategia adaptativa para producir semillas que pueden germinar en un intervalo más amplio de condiciones ambientales.
Temperatura x color de la semilla
Los distintos colores de la semilla aquí estudiados pueden implicar mejores aptitudes para germinar a diferentes temperaturas; es decir, puede ser una estrategia adaptativa ante las variaciones en temperatura, por las siguientes razones. Primero, la especie coloniza áreas perturbadas, requiere de radiación solar directa. Segundo, los colores oscuros absorben más radiación infrarroja, y se calientan más. Entonces se supone que en presencia de luz y a temperaturas menores (dentro del intervalo de germinación de las temperaturas adecuadas para las semillas de la especie), las semillas oscuras tenderán a presentar una mayor germinación, y las claras una menor. Con temperaturas altas las semillas negras germinan poco debido a los pigmentos; mientras que las semillas café claro, además de tener menos pigmentos, absorben menos calor, se pueden mantener a una temperatura más cercana a la óptima y germinar mejor.
La tendencia anterior se cumple en parte en la presente interacción: la germinación de semillas café claro cambió por la temperatura, pues germinaron casi al doble en el régimen cálido que en el régimen más fresco. A su vez, a bajas temperaturas las semillas café oscuro germinaron más que las café claro. Las semillas café oscuro resultaron más estables y con mayor germinación ante los regímenes de temperatura porque están más cerca de uno u otro de los extremos de color (café claro y negro), ya que las primeras germinan mejor a alta temperatura y las segundas germinan poco a cualquier temperatura.
Tamaño x color de la semilla
Sólo en las semillas café claro hubo diferencias en la capacidad germinativa entre semillas grandes y pequeñas. Las semillas café claro pequeñas no germinaron bien por la influencia del tamaño de la semilla y posiblemente por una temperatura menor a la óptima.
Temperatura x tamaño x color de la semilla
En la interacción temperatura x tamaño x color la mayor germinación en las semillas café claro grandes en la temperatura alta, pudo deberse a la menor absorción de radiación y la menor concentración de sustancias inhibidoras de la germinación, al mayor vigor de las semillas y a una temperatura óptima o cercana a ésta. Las semillas café claro y pequeñas no germinaron en altas temperaturas por la influencia del tamaño de la semilla.
Con temperatura alta las semillas grandes y chicas café oscuro no tuvieron diferencias entre sí, pero con temperatura baja las semillas café oscuro pequeñas tuvieron menor germinación que las semillas café oscuro grandes, debido a la influencia del tamaño de la semilla. Las semillas café oscuro grandes germinaron más con temperaturas bajas que con temperaturas altas.
Con temperaturas altas las semillas café claro grandes responderán mejor, pero con menores temperaturas las semillas café oscuro y grandes tendrán mayor capacidad germinativa. La escasa germinación de las semillas negras en todos los tamaños y temperaturas, puede relacionarse con la presencia de sustancias inhibidoras en la cubierta seminal.
CONCLUSIONES
C. obtusifolia es una especie que produce semillas de diferentes tamaños y tonalidades. Como en otras especies, las semillas grandes exhibieron una mayor germinación que las pequeñas. En cuanto al color, las café oscuro germinaron bien en ambos regímenes de temperatura, mientras que las café claro redujeron su germinación a la mitad en el régimen de temperatura más fresco. Dado que las semillas café oscuro son las más abundantes, la mayor parte de las semillas que producen los árboles son estables ante variaciones de temperatura, y se espera que dominen el banco de semillas de la especie.
En el ambiente más cálido las semillas grandes café claro mostraron mayor germinación que las demás categorías. Sin embargo, en el ambiente más fresco, las semillas grandes café oscuro fueron superiores, evidenciando aptitudes diferenciales entre las combinaciones de tamaño y color de semilla para germinar a distintas temperaturas.
LITERATURA CITADA
Álvarez B., E. 1997. Cecropia obtusifolia (Chancarro). In: González S., E., R. Dirzo, y R. C. Vogt (eds). Historia Natural de los Tuxtlas. CONABIO. Instituto de Biología. Instituto de Ecología. México, D. F. pp: 109–114. [ Links ]
Álvarez B., E., and M. Martínez R. 1992. Demography and allometry of Cecropia obtusifoliaa Neotropical pioneer tree: an evaluation of the climax–pioneer paradigm for tropical forest. J. Ecol. 80: 275–290. [ Links ]
Armstrong D. P., and M. Westoby. 1993. Seedling from large seed tolerate defoliation better: a test using phylogenically independent contrast. Ecology. 74: 1092–1100. [ Links ]
Baskin, C. C., and J. M. Baskin. 2001. Seeds. Ecology, Biogeography and Evolution of Dormancy and Germination. Academic Press. San Diego. 666 p. [ Links ]
Bonfil, C. 1998. The effects of seed size, cotyledon reserves, and herbivory on seedlings survival and growth in Quercus rugosa and Quercus laurina (Fagaceae). Am. J. Bot. 85: 79–87. [ Links ]
Bonner, F. T., J. A. Vozzo, W. W. Elam, and S. B. Land Jr. 1994. Tree Seed Technology Training Course. Instructor's Manual. USDA, FS. General Technical Report SO–106. New Orleans, Louisiana, USA. 160 p. [ Links ]
Burslem, D. F. R. P., and J. Miller. 2001. Seed size, germination and seedling growth rates in three tropical tree species. J. Trop. For. Sci. 13: 148–161. [ Links ]
Cordazzo, C. V. 2002. Effect of seed mass on germination and growth in three dominant species in southern Brazilian coastal dunes. Braz. J. Bot. 62: 427–435. [ Links ]
Debeajun, I., L. Kloosterziel M., and M. Koorneef. 2002. Influence of the testa on seed dormancy, germination, and longevity in Arabidopsis. Plant Physiol. 122: 304–414. [ Links ]
Duran, J. M., and N. Retamal. 1989. Coat structure and regulation of dormancy in Sinapis arvensis L. seeds. J. Plant Physiol. 135: 218–222. [ Links ]
Garrido, J. L., P. J. Rey, y C. M. Herrera. 2005. Fuentes de variación en el tamaño de la semilla de la herbácea perenne Helleborus foetidus L. (Ranunculaceae). Ann. Jardín Bot. Madrid. 62: 115–125. [ Links ]
Halpern, S. L. 2005. Sources and consequences of seed size variation in Lupinus perennis (Fabaceae): adaptive and non–adaptive hypothesis. Am. J. Bot. 92: 205–213. [ Links ]
Hendrix, S. D. 1984. Variation in seed weight and its effects on germination in Pastinaca sativa L. (Umbelliferae). Am. J. Bot. 71: 795–802. [ Links ]
Jurado, E., and M. Westoby. 1992. Seedling growth in relation to seed size among species of arid Aust. J. Ecology. 80: 407–416. [ Links ]
Khan M., P. B. Cavers, M. Kane, and K. Thompson. 1996. Role of the pigmented seed coat of proso millet (Panicum miliaceum L.) in imbibition, germination and seed persistence. Seed Sci. Res. 7: 21–25. [ Links ]
Krishnaveni, K., and K. Sivasubramanian. 2001. Effect of seed size on seed quality in sunflower cv. Morden. Madras Agr. J. 88: 133–134. [ Links ]
Leishman, M. R., I. J. Wright, A. T. Moles, and M. Westoby. 2000. The evolutionary ecology of seed size. In: Fenner, M. (ed). The Ecology of Regeneration in Plant Communities. CABI Pub. London. pp: 31–57. [ Links ]
Lobova, A., L. Mori A., S. F. Blanchard, H. Peckham, and C. Dominique P. 2003. Cercopia as food resource for bats in French Guiana and the significance of fruits structure in seed dispersal and longevity. Am. J. Bot. 90: 388–403. [ Links ]
Mares D., J. 1994. Mechanism and genetic control of dormancy in wheat. In: Proceedings 1st International Symposium on Plant Dormancy. August 4–6, 1994. Corvallis, OR. pp: 39. [ Links ]
Mazer, S. J., A. A. Snow, and M. L. Stanton. 1986. Fertilization dynamics and parental effects upon fruit development in Raphanus raphanistrum: Consequences for seed size variation. Am. J. Bot. 73: 500–511. [ Links ]
Méndez, M. 1997. Sources of variation in seed mass in Arum italicum. Int. J. Plant Sci. 158: 298–305. [ Links ]
Moegenburg, S. M. 1996. Sabal palmetto seed size–causes of variation, choices of predators, and consequences for seedlings. Oecologia. 106: 539–543. [ Links ]
Pennington, T. D., y J. Sarukhán K. 2005. Árboles Tropicales de México. Manual para la Identificación de las Principales Especies. 3ª ed. UNAM–FCE. México, D. F. 523 p. [ Links ]
Radford, B. J. 1977. Influence of size of achenes sown and depth of mowing on growth and yield of dryland oilseed sunflowers (Helianthus annus) on the Darling Downs. Aust. J. Exp. Agric. Anim. Husb. 17: 489–494. [ Links ]
Todd, J., and L. O. Vadkin. 1996. Duplications that suppress and deletions that restore expression from a Chalcone syntrase multigene family. Plant Cel. 8: 687–699. [ Links ]
van Mölken, T., L. D. Jorritsma–Wienk, P. H. W. van Hoek, and H. de Kroon. 2005. Only seed size matters for germination in different populations of the dimorphic Tragopogon pratensis subsp. pratensis (Asteraceae). Am. J. Bot. 92: 432–437. [ Links ]
Vázquez Y., C., and H. Smith. 1982. Phytochrome control of seed germination in the tropical rain forest pioneer trees Cecropia obtusifolia and Piper auritum and its ecological significance. New Phytol. 92: 477–485. [ Links ]
Weis, M. I. 1982. The effects of propagule size on germination and seedling growth in Mirabilis hirsuta. Can. J. Bot. 60: 1868– 1874. [ Links ]
Zimmerman, J. K., and M. I. Weis. 1983. Fruit size variation and its effects on germination and seedling growth in Xanthium strumarium. Can. J. Bot. 61: 2309–2315. [ Links ]

