











 text new page (beta)
text new page (beta) Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO 
 Permalink
PermalinkIntroducción
La familia Campanulaceae incluye 14 géneros y 117 especies para México (Rzedowski, 2019). Esta familia comprende el género Lobelia que se caracteriza por ser casi cosmopolita con 423 especies en 18 secciones. La sección Stenotium presenta amplia distribución y es la de mayor diversidad de especies con 153 (Lammers, 2011); debido a la presencia de flores espolonadas esta sección se divide en especies espolonadas y no espolonadas. Las especies espolonadas de Lobelia crecen en áreas húmedas en bosques templados desde Estados Unidos hasta Costa Rica. En particular, 10 especies crecen en la vertiente del Pacífico de la Sierra Madre Occidental en México (Gutiérrez-Sánchez et al., 2018). Dentro de las especies espolonadas de la sección Stenotium se encuentra Lobelia villaregalis Ayers, una especie endémica del Área de Protección de Flora y Fauna del Bosque La Primavera (APFFLP), Jalisco. La especie se caracteriza por ser una hierba perenne de 15-20 cm de alto, con tallos decumbentes, hojas simples ovadas de 15-30 mm de largo, margen dentado; inflorescencia racemosa, pedúnculo de 5-7 cm; flores de color rosa, zigomorfas, simpétalas de 1-1.2 cm, con espolón cónico de 8.5-10 mm y apertura dorsal; estambres 5-5.5 mm, filamentos blancos, connados y anteras azules, cuyo fruto es una capsula con semillas numerosas. Su distribución es particularmente restringida, de hecho cuando la especie fue descrita sólo se conocía una localidad con una población de 5 individuos (Ayers, 1987). Favorablemente, se han encontrado poblaciones en otras localidades dentro del APFFLP (Hernández-López, en proceso), sin embargo, se hallan cerca de senderos recreativos y asentamientos irregulares lo que amenaza su existencia. Desafortunadamente, no es posible establecer estrategias adecuadas para su conservación dado que se desconocen las características del hábitat donde se desarrolla.
La distribución y abundancia de las comunidades vegetales está determinada por diversos factores (bióticos y abióticos), entre los que destacan: los mecanismos de dispersión, capacidad de adaptación a nuevas condiciones del hábitat e interacciones con otras especies (Wiens y Donoghue, 2004). No obstante, se ha venido señalando que un factor importante para la distribución de las especies vegetales, así como para su posterior propagación, es el conocimiento de las características del suelo donde se desarrollan (Huerta-Martínez et al., 2014; 2012). Las propiedades físicas del suelo como temperatura, textura, humedad y cantidad de materia orgánica, ayudan a la formación de microhábitats específicos para las plantas, particularmente las especialistas (Brady y Weil, 2002). Además de las características fisicoquímicas del suelo, los aspectos biológicos del mismo determinan el microhábitat de las especies vegetales, donde dentro de las relaciones más significativas que establecen es la denominada simbiosis micorrícica, que es un fenómeno ecológico-evolutivo producto de la interacción entre hongos y plantas con el suelo y el ambiente atmosférico que les ha brindado entre otras cosas: estabilización del suelo, mejor acceso a agua y nutrientes, así como bioprotección (estrés hídrico, patógenos, contaminantes del suelo) y bioregulación (Neri-Luna y Villarreal-Ruiz, 2012; Smith y Read, 2008). Asimismo, la simbiosis micorrícica es importante para aumentar las probabilidades de supervivencia y establecimiento de las plántulas, lo cual es determinante para los programas de re-población de especies (Davidson et al., 2016).
En el APFFLP se han realizado numerosas investigaciones (Gallegos, 2011). En lo que a la flora se refiere, existen diversos estudios, algunos con énfasis en grupos taxonómicos particulares, de localidades específicas y otros sobre diversos grupos biológicos en toda el área. Entre ellos destacan los trabajos de Curiel (1988), Reyna Bustos (1989) y CONANP (2000), en este último se da cuenta de 961 especies, 419 géneros y 107 familias de plantas vasculares, donde Campanulaceae incluye cinco taxa, pero desafortunadamente no se lista a L. villaregalis.
Con el objetivo de contribuir al conocimiento biológico que ayude a la conservación de L. villaregalis, en este estudio se describen algunas de las condiciones ambientales de las dos localidades donde la especie se ha encontrado, en particular, se dan a conocer las características del suelo, su potencial relación con hongos micorrícicos y la composición florística asociada.
Métodos
Área de estudio
El Bosque la Primavera fue declarado en 1980 como Área de Protección de Flora y Fauna (APFFLP) con una superficie de 30,500 ha. Se ubica en la parte central del estado de Jalisco, al oeste de la Zona Metropolitana de Guadalajara, entre las coordenadas extremas 103º28' a 103º42' de longitud oeste y 20º32' a 20º44' de latitud norte (CONANP, 2000). Abarca la superficie de cuatro municipios que en orden descendente por su cobertura en el área son: Zapopan, Tala, Tlajomulco y El arenal (Fig. 1).
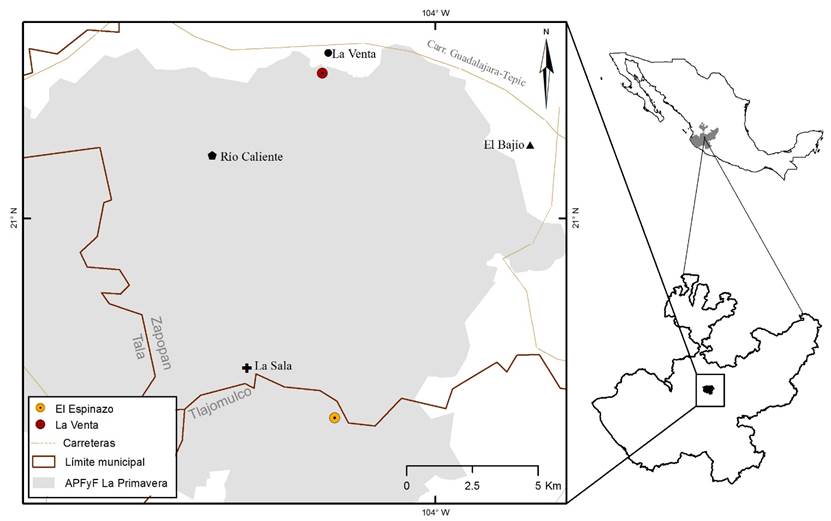
El APFFLP presenta un rango de elevación de los 1400 a 2200 m s.n.m y forma parte de la Sierra La Primavera, la cual fisiográficamente pertenece a una zona donde se traslapan el Eje Neovolcánico Transversal y la Sierra Madre Occidental. Desde el punto de vista geológico, la Sierra de La Primavera es, en términos técnicos, una caldera volcánica formada por erupciones de tobas y piroclastos (Valdivia Ornelas, 2018). Los procesos geológicos y geomorfológicos recientes en combinación con el enfriamiento de tobas y su erosión posterior, le confieren al relieve de La Primavera rasgos únicos que deberían considerarse como Geopatrimonio (González Torreros et al., 2018). Los tipos de suelos mejor representados son regosol derivado del intemperismo de la toba, pómez y riolita, y litosol, resultante de procesos erosivos, los cuales cubren el 92% y 8% del área, respectivamente (CONANP, 2000). Las comunidades vegetales registradas, con base en la clasificación de Rzedowski (2006), son: bosque de encino (Quercus), bosque de encino-pino (Quercus-Pinus), bosque de pino (Pinus), bosque tropical caducifolio, vegetación riparia y vegetación secundaria o ruderal (Reyna Bustos, 2004).
Hasta el año 2009, sólo se había encontrado a L. villaregalis en dos localidades, una de ellas en un paredón ubicado al sur (“El Espinazo”) donde las poblaciones crecen en un solo punto y la otra donde se observó en manchones de unos cuantos individuos dispersos a lo largo de una cañada localizada al norte, cerca del pueblo La Venta del Astillero (“La Venta”), ambas en el APFFLP (Hernández-López et al., 2009) (Fig. 2, C y D). La vegetación circundante a las localidades donde se ubica L. villaregalis es bosque de pino-encino, siendo el clima predominante templado subhúmedo y semicálido subhúmedo del tipo estacional, conformado por dos periodos: estiaje (seco) que ocurre desde octubre hasta junio y el de lluvias (húmedo) en los meses restantes (CONANP, 2000).

Muestreo de suelo
Características del suelo
Durante la época de estiaje del año 2010, en cada localidad donde se registró la presencia de L. villaregalis, se midió la profundidad del suelo alrededor de las raíces de 50 individuos. Por otra parte, tanto en “La Venta” como en “El Espinazo”, se colectaron muestras de suelo ya sea cerca a las raíces de las plantas (n = 6) como de sitios donde la especie no estaba presente. Sin embargo, debido a que el suelo es poco profundo, las muestras de los diferentes sitios se tuvieron que mezclar conformando una sola (aproximadamente 500 g), es decir, en total se obtuvieron 2 muestras por cada localidad (una del sitio donde crece la especie y otra donde no se observa). El suelo fue analizado en el Laboratorio de Agrología de la Universidad de Guadalajara, donde se realizaron las siguientes determinaciones: Textura (método de Bouyoucos), materia orgánica (método de Walkey-Black), pH mediante la relación 1:2 suelo:agua destilada, nitrógeno nítrico y nitrógeno amoniacal, fósforo, potasio, calcio, magnesio y manganeso (método de Morgan). Los resultados del análisis fueron interpretados con base en los criterios de calidad de suelo de SAGARPA y FAO (2012) y Castellanos et al. (2000).
Asociación Micorrícica
Con la finalidad de evaluar si las raíces de L. villaregalis establecen relación con hongos micorrícicos del suelo y en caso de haberla, si hay una potencial variación estacional de la colonización micorrícica, en cada una de las localidades se colectaron 5 plantas (con una altura aproximada de 15-20 cm y con hojas jóvenes) en dos épocas del año: la primera de ellas durante la fase de floración de la especie que ocurre en el estiaje, y la otra en la estación húmeda. Sin embargo, debido al bajo número de individuos en la localidad “La Venta”, la comparación entre estaciones sólo se llevó a cabo en la localidad “El Espinazo”.
Las plantas de L. villaregalis se llevaron al laboratorio, donde las raíces se limpiaron y almacenaron en alcohol (50%); posteriormente, se procesaron siguiendo la técnica de tinción de Kormanik y McGraw (1982). Para cuantificar el porcentaje de colonización de las raíces por el hongo micorrícico (%RC) se utilizó el método de intersección magnificada propuesto por McGonigle et al. (1990).
Análisis estadísticos
El análisis estadístico se realizó empleando el programa estadístico R versión 3.4.4 (R Core Team, 2018). Para evaluar si existen diferencias en la fertilidad del suelo entre sitios (donde la especie crece y donde está ausente) dentro de la misma localidad, se realizó una prueba de Mann-Withney pareada. Para evaluar las diferencias en %RC entre las localidades se llevó a cabo una prueba de comparación de dos muestras (T-student). Asimismo, se determinó si existían diferencias entre el estiaje y la estación de lluvias utilizando la prueba T-student pareada en la localidad “El Espinazo”.
Muestreo de vegetación
Para documentar la composición florística de las dos localidades donde crece L. villaregalis se realizaron salidas de campo entre diciembre del 2007 y marzo del 2010. El material recolectado se procesó con base en Lot y Chiang (1986) y se depositó en el Herbario Luz María Villareal de Puga de la Universidad de Guadalajara (IBUG). Ahí mismo, se llevó a cabo la determinación taxonómica de las muestras utilizando bibliografía especializada para posteriormente integrar un listado de las especies encontradas, ordenado alfabéticamente por nombre científico de los taxa y por presencia. Aparecen primero los taxa compartidos entre las dos localidades (Apéndice). En algunos casos, no fue posible determinar el material hasta el nivel de especie ya que al momento de la colecta no presentaba estructuras fértiles, por lo que en el listado sólo se indica el género y los datos del colector aparecen en blanco. Para analizar la presencia de malezas entre las especies registradas, se revisó la obra de Villaseñor y Espinosa-García (1998) y la plataforma de las malezas de México (CONABIO, 2019). Para describir las características fisonómicas de las localidades en “El Espinazo” se midió la longitud del paredón y la altura en tres puntos diferentes, mientras que en “La Venta” se definieron cinco puntos a lo largo de la cañada en los cuales se calculó el ancho y alto.
Resultados
Descripción del suelo
Se observó que L. villaregalis crece en suelos franco arenosos, poco profundos (promedio de 1.5 cm) con un pH que oscila entre 4.5 y 5.5. Este tipo de suelo por su naturaleza arenosa es altamente permeable por lo que tienen poca retención de agua y nutrientes, así como baja inercia térmica (Castellanos et al., 2000). De acuerdo con SAGARPA y FAO (2012) y Castellanos et al. (2000) en ambas localidades, la planta crece en suelos con porcentajes bajos de agua aprovechable; niveles bajos de nitrógeno nítrico, nitrógeno amoniacal, calcio y magnesio; niveles medios de fósforo; y niveles altos y medio altos de potasio y manganeso (Cuadro 1). En general, los valores de fertilidad entre ambas localidades son similares. Sin embargo, con excepción del nitrógeno y la materia orgánica, los niveles de nutrientes fueron más altos en “La Venta que en “El Espinazo”. Desafortunadamente, dado el bajo número de replicas por localidad (n = 1) no se puedo establecer si tales diferencias son estadísticamente significativas. Al comparar entre los sitios donde crece la planta y donde está ausente en cada una de las localidades no se observaron diferencias significativas (p > 0.05).
Cuadro 1 Características del suelo de los sitios donde hay presencia y ausencia de L. villaregalis dentro de las dos localidades del estudio. El valor de V y su probabilidad fueron calculados empleando la prueba Mann-Withney pareada. ND: no detectable.
| Variable | V | La Venta | El Espinazo | p | ||
|---|---|---|---|---|---|---|
| - | Presente | Ausente | Presente | Ausente | - | |
| Clase textural | - | Fa | Fa | Fa | Af | NA |
| Agua aprovechable % | 0 | 13 | 12 | 9 | 6 | 0.5 |
| Materia orgánica % | 2 | 1.88 | 2.26 | 4.24 | 2.19 | 0.9 |
| pH | 0 | 5 | 5.5 | 4.5 | 4.7 | 0.5 |
| Nitrógeno nítrico ppm | 0 | 0.5 | 1 | 1 | 1 | 0.9 |
| Nitrógeno amoniacal ppm | 0 | 6 | 12 | 12 | 12 | 0.9 |
| Fósforo ppm | 0 | 25 | 25 | 25 | 25 | 1 |
| Potasio ppm | 0 | 250 | 250 | 250 | 250 | 1 |
| Calcio ppm | 1 | 500 | 1200 | 500 | 500 | 0.9 |
| Magnesio ppm | 0 | 25 | 25 | 12 | 12 | 0.9 |
| Manganeso ppm | 3 | 8 | 5 | 2 | ND | 0.5 |
Asociación micorrícica
Se encontró que L. villaregalis tiene una asociación micorrícica del tipo arbuscular, la cual se establece entre hongos del Phyllum Glomeromycota y el 80% de las familias de las plantas vasculares (Smith y Read, 2008). En este estudio, se registró un %RC del 36.5 ± 5.5 en “La Venta” y 31.3 ± 9.8 en “El Espinazo” (Cuadro 2), sin diferencia significativa entre localidades (t8 = 0.459, p > 0.05). Por otra parte, al comparar el %RC entre diferentes épocas del año en la localidad de “El Espinazo” tampoco se detectaron diferencias significativas (t8 = 0.303, p > 0.05; Cuadro 3).
Cuadro 2 Porcentaje de colonización en la raíz de L. villaregalis por hongos micorrícicos arbusculares en cada localidad. Los valores representan el promedio y error estándar de la media (n = 5). El valor de t y su probabilidad fueron calculados empleando la prueba T-student. RC: colonización total de las raíces.
| Estructura | t | La Venta | El Espinazo | p |
|---|---|---|---|---|
| Arbúsculo % | 0.2 | 0.92 ± 0.33 | 5.35 ± 3.37 | 0.82 |
| Vesículas % | 0.2 | 5.30 ± 1.15 | 3.56 ± 0.58 | 0.83 |
| Hifas % | 0.3 | 30.29 ± 4.42 | 22.40 ± 6.33 | 0.74 |
| RC % | 0.6 | 36.52 ± 5.51 | 31.32 ± 9.89 | 0.53 |
Cuadro 3 Porcentaje de colonización en la raíz de L. villaregalis por hongos micorrícicos arbusculares en la localidad “El Espinazo” en dos diferentes estaciones del año. Los valores representan el promedio y error estándar de la media (n = 5). El valor de t y su probabilidad fueron calculados empleando la prueba T-student. RC: Colonización total de las raíces.
| Estructura | t | Estiaje | Lluvias | p |
|---|---|---|---|---|
| Arbúsculo % | 0.6 | 5.35 ± 3.37 | 2.85 ±1.20 | 0.58 |
| Vesículas % | -0.4 | 3.56 ± 0.58 | 4.04 ± 1.07 | 0.71 |
| Hifas % | -1.9 | 22.40 ± 6.33 | 41.98 ± 5.16 | 0.13 |
| RC % | -1.2 | 31.32 ± 9.89 | 48.88 ± 5.10 | 0.28 |
Descripción de la vegetación
En el conjunto de las dos localidades estudiadas, se registró un total de 44 familias, 102 géneros y 140 taxa de plantas vasculares, incluyendo cinco categorías intraespecíficas (Apéndice). Asteraceae concentra la mayor riqueza de especies con 44 taxa (32% del total), le siguen Poaceae y Pteridaceae con 6 y 5% de las especies, respectivamente. Los géneros con mayor número de especies corresponden a Ageratina con siete y Pseudognaphalium con cinco, ambos se incluyen en Asteraceae. La forma biológica mejor representada es la herbácea con 102 taxa (73% del total), le sigue la arbórea con 18 (13%), la arbustiva con 16 (11%) y sólo cuatro taxa son trepadoras. De las especies registradas, 22 (16%) son consideradas como malezas, no obstante, sólo tres son de origen exótico o incierto. Entre las plantas no vasculares, se registra la presencia de musgo (Bryopsida) en las dos localidades, pero con mayor abundancia en “El Espinazo”.
Respecto a la composición de especies, se encontró que 31 taxa (22.1%) se comparten entre las dos localidades, de las cuales 19 son hierbas, 7 son árboles y el resto corresponden a arbustos y trepadoras. Se consideran malezas cinco de los taxa compartidos (Lobelia laxiflora, Lopezia racemosa, Oplismenus burmannii, Paspalum humboldtianum y Wigandia urens). La localidad de “La Venta” alberga 101 taxa (72% del total) de los cuales 70 (50%) son exclusivos y en “El Espinazo” se registran 70 (50% del total), de ellos 39 (28%) no se comparten (Apéndice). De las especies exclusivas de “La Venta” 84% (59 taxa) corresponden a herbáceas que en su mayoría (64%) pertenecen a la familia Asteraceae y a siete familias de helechos y afines. Se consideran malezas 14 taxa. Destacan tres especies de árboles; Clethra rosei, Morella cerifera y Oreopanax peltatus. Por otra parte, en “El Espinazo” los taxa exclusivos incluyen 24 hierbas (61.5%) de las cuales casi la mitad forman parte de las Asteraceae. Las especies arbustivas y arbóreas conforman el restante 38.5%. Sólo tres taxa exclusivos se reconocen como malezas. Entre los árboles se listan; Pinus douglasiana, P. maximinoi, Clethra hartwegii, Agarista mexicana var. pinetorum, Arbutus madrensis y Comarostaphylis glaucescens.
Las dos localidades donde crece L. villaregalis se encuentran en áreas de bosque de encino (Quercus) y bosque mixto de encino-pino, sin embargo, existen diferencias en la composición florística. A continuación, se describen las características de las dos localidades y la composición de la vegetación: La localidad de “La Venta” se ubica entre los 1530-1565 m s.n.m. donde L. villaregalis se encuentra en la parte alta y media de la cañada creciendo de forma dispersa a lo largo de un km del estrecho cañón con exposición noroeste, de forma que sólo recibe luz solar en el invierno durante algunas horas de la mañana. La cañada alcanza los 10.5 m en su parte más alta y en otras porciones no pasa de los 4 m. Lo ancho varía entre 3 y 1.30 m; la cañada se abre hacia la parte alta donde llega a los 7.5 m (Fig. 2D). En el fondo de esta cañada los árboles son escasos y se trata de especies características de lugares húmedos como Fraxinus uhdei, Morella cerifera, Oreopanax peltatus y Prunus serotina. En el estrato arbustivo se observan Ageratina areolaris, A. dolichobasis, Cestrum nitidum, Hyptis albida y Cunila pycnantha. El estrato herbáceo que cubre las paredes se compone de algunos helechos y afines como Asplenium monanthes, Thelypteris rudis, Cheilanthes angustifolia, Adiantum capillus-veneris y Selaginella pallescens. También se encuentran algunos pastos como Muhlenbergia tenella, M. brevifolia, Oplismenus burmannii y Paspalum humboldtianum; además de Begonia gracilis, Drymaria gracillima, Moussonia elegans, Ageratina malacolepis y Heuchera orizabensis.
Respecto a la localidad “El Espinazo”, esta se localiza entre los 2060-2080 m s.n.m. y es un paredón con exposición noroeste que debe su existencia a la apertura de un camino que asciende al cerro Planillas donde se localiza una torre de vigilancia. Presenta en forma general, la misma composición de suelo y rocas que la localidad de “La Venta”, pero la población de L. villaregalis es quizá 20 veces más numerosa aquí y concentrada en una superficie más reducida, creciendo principalmente en la parte media y alta del paredón. Éste mide 50 m de largo y su altura es irregular alcanzando 17 m en la parte más alta, disminuyendo hasta los 3 m (Fig. 2C). Es una pared casi vertical que en el invierno recibe poca radiación solar directa en las mañanas, el resto del día está bajo sombra dada en parte por otro paredón más pequeño ubicado frente a éste y por los árboles y arbustos que la coronan o que crecen entre la población de Lobelia. Los árboles que se encuentran adyacentes a esta población de L. villaregalis son: Clethra hartwegii, Comarostaphylis glaucescens, Pinus devoniana, P. oocarpa, P. maximinoi, Quercus resinosa y Q. viminea. Entre los arbustos se registran a Calliandra hirsuta, Monnina schlechtendaliana, Gaultheria erecta, Vaccinium stenophyllum y Wigandia urens. En cuanto a las herbáceas sobresalen Dalea roseiflora, Cerastium brachypodon, Salvia lavanduloides, Heterocentron mexicanum, Lopezia racemosa, Heuchera orizabensis, Bessera elegans y los pastos Aegopogon cenchroides, Paspalum humboldtianum y Oplismenus burmannii. En esta zona es frecuente observar especies arbóreas de los géneros Clethra, Pinus y Prunus y arbustivas como Wigandia creciendo en oquedades del paredón. No obstante, la mayoría no llega a establecerse y permanecen por poco tiempo debido a la erosión del suelo, mientras que las especies herbáceas tienen más éxito.
Cabe mencionar que L. villaregalis se encuentra en comunidades típicas de cañadas húmedas, por lo menos como sucede en las cañadas que forman parte del APFFLP. Las especies que comparten las condiciones ambientales en suelo y humedad entre las dos localidades son 31, algunas se nombran a continuación: Fraxinus uhdei, Prunus serotina, Pinus devoniana, P. oocarpa, Quercus coccolobifolia, Q. resinosa y Q. viminea entre las arbóreas, y de las herbáceas Lopezia racemosa, Heuchera orizabensis, Paspalum humboldtianum, Oplismenus burmannii, Lobelia laxiflora y Moussonia elegans, entre otras (Apéndice).
Discusión
Ante la constante modificación de hábitats por razones antropogénicas, la generación de información sobre las características del hábitat donde las especies se desarrollan resulta urgente para su conservación, particularmente para aquellas que son endémicas restringidas. Las especies con distribuciones restringidas evolutivamente se han especializado con base en uno o más factores ambientales (Boisson et al., 2017; Irl et al., 2015). Esta especialización, las hace vulnerables a cambios en su hábitat y por lo tanto son especies prioritarias para la conservación (Niskanen et al., 2019). En este estudio, se generó información sobre algunos de los factores que conforman el hábitat donde se desarrolla L. villaregalis dentro del APFFLP, la cual se encuentra amenazada por actividades derivadas del crecimiento urbano de la Zona Metropolitana de Guadalajara.
Los resultados muestran que la especie se desarrolla en suelos con niveles bajos de nutrientes dada su alta permeabilidad debido a su naturaleza arenosa lo cual es característico de los suelos tipo regosol del APFFLP (CONANP, 2000). El regosol es un tipo de suelo joven presente en el 18.5% del territorio del país (SEMARNAT, 2002) principalmente en las regiones biogeográficas de la Sierra Madre Occidental, Eje Neovolcánico Transversal y Sierra Madre del Sur (SEMARNAT, 1998). El no encontrar diferencias significativas entre sitios donde L. villaregalis está presente y ausente, sugieren que la cañada y el paredón no presentan microhábitats definidos por características edáficas. Sin embargo, la baja retención de humedad del suelo, la presencia de taxa de sitios húmedos y la baja exposición solar de las localidades, indican que otros factores, por ejemplo, porcentaje de humedad relativa o cantidad de radiación solar podrían estar siendo de alguna manera determinantes en la distribución de la especie. Lo anterior contrasta con otros estudios donde se ha demostrado que el suelo es el factor que más influye en la diversidad vegetal (Huerta-Martínez et al., 2014, 2012). Asimismo, estos resultados sugieren que L. villaregalis podría estar presente en otras cañadas húmedas dentro del APFFLP y potencialmente en otras zonas montañosas del país, aunque debido al relieve geológico único del APFFLP (González Torreros et al., 2018) que pudo haber llevado a procesos de especiación y dispersión particulares al área, la probabilidad de que la especie se encuentre fuera del área protegida sea baja.
En ambas localidades, L. villaregalis estableció una asociación micorrícica del tipo arbuscular, aunque en un bajo %RC comparada con otras especies (Olivera-Morales et al., 2011; Montañez Orozco et al., 2010). De la misma manera, al parecer se mantiene el %RC, a pesar del cambio de estaciones durante el año como se pudo constatar con las plantas colectadas en “El Espinazo”. La estabilidad en la asociación micorrícica entre la época de estiaje y lluvias no es inusual, ya que se ha observado en otras especies como Coccothrinax readii Quero (Polanco et al., 2013) y Agave angustifolia Haw (Ochoa-Meza et al., 2009). Por un lado, se menciona que una estrategia para enfrentar la limitación por nutrientes y agua sería el establecimiento de relaciones micorrícicas (Smith y Read, 2008; Ruiz-Lozano, 2003), por otra parte, una alta colonización en suelos pobres podría convertirse en una desventaja, ya sea por un alto gasto metabólico de la planta al tener que retribuir con carbono a los hongos micorrícicos (Lendenmann et al., 2011; Lynch y Ho, 2005; Nielsen et al., 1998); o a que la planta y el hongo micorrícico compiten por nutrientes (Johnson et al., 2015). Sin embargo, el hecho de haber reportado que L. villaregalis presenta asociación micorrícica del tipo arbuscular es un paso importante en el conocimiento de la biología de esta planta, donde el identificar las especies de hongos micorrícicos que se asocian, así como diseñar experimentos para probar su función y contribución en el desarrollo y reproducción de la planta son apremiantes.
En cuanto a la composición de la vegetación acompañante, es de esperarse que Asteraceae sea la familia con mayor riqueza de especies en las dos localidades, ya que es un grupo altamente representado en toda el área protegida (CONANP, 2000) y en otras áreas con bosques templados en Jalisco (García-Martínez y Rodríguez, 2018; Harker et al., 2017). No obstante, los sitios estudiados mostraron diferencias tanto en la riqueza como en la composición de especies, lo cual puede explicarse por el proceso de formación de ambas localidades. En la localidad “El Espinazo” el paredón que ocupa L. villaregalis se formó por la fragmentación de un cerro para construir un camino angosto en el año de 1972 (A. Curiel, comunicación personal). Posteriormente, se reforestó con especies de pino a lo largo del camino construido de modo que, actualmente la vía es estrecha y los árboles han alcanzado cerca de los 10 m. Este hecho contrasta con la localidad de “La Venta”, donde al parecer la cañada se ha formado con el paso del tiempo y por los procesos naturales. Otra diferencia fundamental entre las localidades es la superficie que ocupan, ya que como se mencionó, “El Espinazo” mide apenas 50 m de largo, mientras que “La Venta” alcanza un km de longitud, aproximadamente.
En “La Venta” habitan cuatro especies de orquídeas y 13 helechos exclusivos, lo que aunado a la presencia de Oreopanax peltatus y Morella cerifera, entre otras, reflejan un estado más conservado de esta localidad. No obstante que alberga el mayor número de especies arvenses (14), su abundancia es baja. En tanto que en “El Espinazo” sus especies exclusivas se distribuyen en un mayor número de familias y el número de malezas exclusivas es menor (3 de 8 presentes) que en “La Venta”. Sin embargo, aquí se observan con mayor abundancia a Lobelia laxiflora, Oplismenus burmannii, Paspalum humboltianum, Baccharis salicifolia, Jaegeria hirta y Wigandia urens las cuales representan especies pioneras que han colonizado el sitio como se ha encontrado en otros bosques mixtos de pino-encino donde miembros de Asteraceae y Poaceae conforman principalmente el contingente de pioneras (Ramírez-Marcial et al., 1992).
Por otra parte, dado que el origen de “El Espinazo” fue una perturbación que implicó el desmonte y la apertura de material parental, puede decirse que L. villaregalis consigue colonizar sitios que, aunque han recibido cierto disturbio, con el tiempo en ellos llegan a crearse microhábitats (paredes con sustrato arenoso, permanentemente húmedas y con poca llegada de radiación solar) que favorecen a la planta. En este caso a L. villaregalis le tomó cerca de 50 años colonizar y establecerse exitosamente en “El Espinazo”. No obstante, eso fue posible gracias a la baja incidencia de perturbación en el sitio en el periodo posterior a la apertura del camino, lo cual se evidencia en parte por la acumulación de una gruesa capa de combustible en el suelo (ausencia de incendios) y la presencia de fauna como Crotalus basiliscus, especie endémica de México y protegida bajo la Norma Oficial Mexicana 059 de la SEMARNAT.
Por tanto, los resultados sugieren que L. villaregalis tiene preferencia por lugares húmedos (posiblemente a causa de desarrollarse en suelos con poca retención de agua) y en buen estado de conservación. Actualmente, ambas poblaciones de L. villaregalis están expuestas al impacto antropogénico, ya sea por su cercanía a asentamientos humanos irregulares o senderos recreativos por lo que es necesaria una regulación del paso de transeúntes, evitar cambios ilegales del uso de suelo y realizar actividades de prevención de incendios para la prevención de daños en ambas poblaciones. Se pretende que los datos generados en esta investigación sean de utilidad para la exploración de nuevas localidades potenciales, así como la realización de estudios ecológicos para orientar la promoción y generación de programas para la conservación y repoblación asistida de L. villaregalis. Estas iniciativas de investigación y acciones de manejo son urgentes, ya que los sitios de estudio aquí documentados, fueron impactados por los incendios forestales que se registraron en el APFFLP en la temporada de estiaje del 2019 y por tal motivo, la permanencia de L. villaregalis en esos sitios es ahora incierta.
Conclusiones
L. villaregalis se desarrolla en suelos franco arenosos, pobres, de poca profundidad y alta permeabilidad en los que establece asociaciones micorrícicas del tipo arbuscular, cuyo porcentaje de colonización se mantiene constante entre estaciones climáticas. La flora acompañante de esta especie y las condiciones generales de las localidades estudiadas reflejan su preferencia por hábitats húmedos y paredes o laderas pronunciadas con sustrato arenoso en buen estado de conservación. Las estrategias para la conservación de la especie, por tanto, deberán incluir acciones que eviten el deterioro de su hábitat. Asimismo, se sugiere que en las estrategias de propagación de esta especie se consideren la incorporación de microorganismos nativos del suelo (hongos micorrícicos) donde crece para aumentar su probabilidad de supervivencia.

