











 text new page (beta)
text new page (beta) Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO 
 Permalink
PermalinkIntroducción
En la actualidad, a través del uso de técnicas dendrocronológicas se han logrado generar múltiples cronologías para diversas especies arbóreas de México, principalmente para los géneros Pinus, Pseudotsuga, Abies y Taxodium (Villanueva-Díaz et al., 2015; Acosta-Hernández et al., 2017). Dichas cronologías han sido desarrolladas a partir de los procesos de fechado, medición y estandarización de anillos de crecimiento (Fritts, 1976; Cook y Holmes, 1986; Cook, 1987).
Las cronologías de anillos de crecimiento aportan datos sumamente relevantes sobre las fluctuaciones del clima, influencia de fenómenos océano-atmosféricos de escala global como el Niño Oscilación del Sur (ENOS), la Oscilación Decadal del Pacífico (ODP) y la Oscilación Multidecadal del Atlántico (OMA), eventos que quedan registrados en el crecimiento radial anual (Villanueva-Díaz et al., 2015; Carlón et al., 2021; Manzanilla-Quiñones et al., 2020), así como registros históricos de sequías y de incendios forestales (Cerano-Paredes et al., 2019; Martínez-Sifuentes, Villanueva-Díaz y Estrada-Ávalos, 2020).
La red de investigación dendrocronológica en México ha aumentado significativamente en la última década, siendo las zonas más estudiadas; las regiones norte y centro del país (Villanueva-Díaz et al., 2015; Acosta-Hernández et al., 2017; Carlón et al., 2021). Mucha de esta información generada se basa en la experiencia del investigador y en el conocimiento de los dueños de los predios forestales, quienes eligen los rodales a muestrear, por lo que no se suele realizar un análisis espacial previo de las condiciones biofísicas de esos potenciales sitios potenciales de muestreo (Carlón et al., 2015).
En la actualidad los modelos de nicho ecológico (MNE) se han convertido en uno de los métodos más comunes y eficientes en la delimitación de las áreas de distribución de especies (Soberón y Peterson, 2005; Phillips et al., 2006; Martínez-Méndez, et al., 2016). En México los MNE se han utilizado en la delimitación de los nichos ecológicos de los taxa de la familia Pinaceae, lo que ha permitido identificar los requerimientos ambientales que delimitan la distribución de esas especies (Martínez-Méndez et al., 2016; Manzanilla-Quiñones, Aguirre-Calderón et al., 2019).
La aplicabilidad de los MNE no se ha limitado exclusivamente a cuestiones biogeográficas, biológicas y ecológicas (Phillips et al., 2006; Soberón y Peterson, 2005; Martínez-Sifuentes, Villanueva-Díaz, Manzanilla-Quiñones et al., 2020) sino que su uso ha servido como una herramienta auxiliar en las propuestas de manejo y conservación de los recursos naturales (Correa Ayram, et al., 2019; Méndez et al., 2020). Por lo que se sugiere el uso de técnicas de MNE en la identificación de sitios potenciales para la toma de datos dendrocronológicos de una región y/o especies de interés.
El estado de Michoacán alberga 22 de los 72 taxa de la familia Pinaceae registrados para México (Madrigal, 1982; Perry, 1991), la mayoría de estas especies se encuentran sin un análisis espacial previo de las condiciones biofísicas desde una perspectiva dendrocronológica. El único estudio de este tipo es el realizado por Carlón et al. (2015) para Pinus pseudostrobus y Abies religiosa en la Reserva de la Biosfera Mariposa Monarca (RBMM). En ese trabajo se obtuvieron buenos resultados en la identificación espacial de sitios y en la posterior evaluación dendrocronológica de las muestras colectadas en campo de ambas especies. Por lo anterior, el potencial para la identificación de sitios y especies para la toma de datos dendrocronológicos en Michoacán es formidable.
Objetivos
Analizar bases de datos geográficas y variables ambientales con el propósito de identificar sitios potenciales para la toma de datos dendrocronológicos en especies de la familia Pinaceae del estado de Michoacán de Ocampo, México. Los objetivos específicos fueron 1) modelar la idoneidad ambiental de especies de pináceas de Michoacán, 2) describir las variables ambientales que más influyen en la distribución potencial de cada especie analizada y 3) identificar los sitios y especies con las características bioclimáticas, topográficas y edáficas potenciales para la posterior toma de datos dendrocronológicos.
Materiales y métodos
Área de estudio
La zona de estudio corresponde al estado de Michoacán, localizado en las coordenadas geográficas 20° 23’ 40’’ y 17° 54’ 54’ latitud norte y los 100° 03’ 47’’ y 103° 44’ 17’’ longitud oeste. El estado colinda al norte con Jalisco y Guanajuato, al noroeste con Querétaro, al este con el Estado de México y Guerrero, al oeste con el Océano Pacífico y Colima y Jalisco y al sur con Guerrero (Fig. 1). El área de estudio abarca una superficie de 58 836.95 km2 (Instituto Nacional de Estadística, Geografía e Informática [Inegi], 2017).

Clima
Los climas registrados en Michoacán corresponden a los tipos A(w) cálido subhúmedo con lluvias en verano, ACm semicálido húmedo con abundantes lluvias en verano, ACw semicálido subhúmedo con lluvias en verano, C(m) templado húmedo con abundantes lluvias en verano, C(w) templado subhúmedo con lluvias en verano, C(E)(m) semifrío húmedo con lluvias en verano, C(E)(w) semifrío subhúmedo con lluvias en verano, BS1(h’) semiseco muy cálido y cálido y BS(h’) seco muy cálido y cálido. Siendo ACm (34.08%), C(w) (28.19%) y AC(w) (20.6%) los climas dominantes (Inegi, 2017).
Edafología
Los suelos corresponden a los tipos Andosol (AN), Arenosol (AR), Calcisol (CL), Cambisol (CM), Durisol (DU), Fluvisol (FL), Histosol (HS), Kastañozem (KS), Leptosol (LP), Luvisol (LV), Phaeozeom (PH), Planosol (PL), Regosol (RG), Umbrisol (UM) y Vertisol (VR). Siendo los suelos dominantes LP (19.62%), LV (17.56%) y AN (13.55%) (Inegi, 2017).
Datos de presencia
Los registros de presencia disponibles de los taxa registrados en el estado de Michoacán (Madrigal, 1982) fueron descargados de las plataformas GBIF (Global Biodiversity Information Facility) y del herbario del Instituto de Biología (MEXU) de la Universidad Nacional Autónoma de México (UNAM).
Los registros geográficos correspondieron a las especies Abies religiosa (H.B.K.) Schlt. Et Cham., Pinus douglasiana Martínez, P. hartwegii Lindl, P. herrerae Martínez, P. lawsonii Roezl., P. leiophylla Schlt. et Cham., P. martinezii Larsen, P. maximinoi Moore, P. devoniana Lindl, P. montezumae Lamb., P. oocarpa Schiede, P. pringlei Shaw, P. pseudostrobus Lindl., P. rzedowskii Madrigal et Caballero y P. teocote Schlt. et Cham. Asimismo, se obtuvieron registros de expediciones realizadas en campo durante 2012-2020 para P. martinezii, P. montezumae, P. pseudostrobus y P. rzedowskii (Delgado et al., 2015).
Limpieza de datos
El proceso de depuración consistió en eliminar los registros de presencia que se localizaban fuera de su distribución natural (Perry, 1991), así como aquellos que no contaban con una fuente de validación oficial. Se eliminó la autocorrelación espacial entre registros de las especies en la plataforma Niche Toolbox (Osorio‐Olvera et al., 2020) lo que ayudó a prevenir el sobreajuste en los modelos por efecto de la autocorrelación espacial (Peterson y Nakazawa, 2008) dejando un solo registro por cada píxel de 90 m. Finalmente se obtuvieron 160 registros (A. religiosa), 30 (P. devoniana), 26 (P. douglasiana), 24 (P. hartwegii), 14 (P. herrerae), 26 (P. lawsonii), 176 (P. leiophylla), 12 (P. martinezii), 13 (P. maximinoi), 94 (P. montezumae), 49 (P. oocarpa), 17 (P. pringlei), 175(P. pseudostrobus), 77 (P. rzedowski) y 28 (P. teocote).
Variables predictoras
Los datos utilizados en la generación de los modelos de idoneidad ambiental fueron de tres tipos: I) bioclimáticas, las cuales representan el comportamiento climático promedio del periodo 1910-2009 en México y tienen una resolución espacial de 90 m (Cuervo-Robayo et al., 2014); II) topográficas: pendiente (Pen) en porcentaje (%), orientación de laderas (Ori) en grados (°) y elevación en metros, generadas a partir de un modelo digital de elevación de 90 m de resolución, y III) edáficas: potencial de hidrógeno (pH) en escala de 1 a 14 y pedregosidad del suelo (Roc) en porcentaje (%) a una escala de 250 m, estas últimas fueron descargadas de la plataforma de Inegi (Inegi, 2014; 2013). Todas las variables fueron homogenizadas a una resolución espacial de 90 m y en formato ascii.
Selección de variables
La selección de variables empleadas en la modelación de idoneidad ambiental se efectuó mediante un análisis del factor de inflación de la varianza (VIF) en el programa R versión 4.0 (R Core Team, 2020) que permitió descartar las variables redundantes (correlación de Pearson con valores mayores o iguales a 0.85 positivos y negativos, p<0.01) y evitar que la multicolinealidad sesgara los resultados de los modelos de distribución (Peterson y Nakazawa, 2008).
Las variables seleccionadas para cada especie fueron 12 (P. leiophylla y P. pseudostrobus), 11 (P. maximinoi y P. teocote), 10 (A. religiosa, P. devoniana, P. douglasiana, P. lawsonii, P. montezumae y P. oocarpa), 9 (P. pringlei), 8 (P. hartwegii y P. martinezii), 6 (P. rzedowski) y 5 (P. herrerae). El número de variables seleccionadas fue mayor en especies de distribución amplia en Michoacán, mientras que las especies de distribución restringida presentaron menos variables (Tabla 1).
Tabla 1 Descripción de las variables ambientales empleadas en la modelación de las 15 especies de estudio.
| Clave | Especie | |||||||||||||||
| Variable | ||||||||||||||||
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 | ||
| Oscilación anual de la temperatura (°C) | Bio7 | * | * | * | * | * | * | * | * | * | * | * | ||||
| Temperatura promedio del trimestre más lluvioso (°C) | Bio8 | * | * | * | ||||||||||||
| Temperatura promedio del trimestre más seco (°C) | Bio9 | * | * | |||||||||||||
| Temperatura promedio del trimestre más cálido (°C) | Bio10 | * | * | * | * | * | * | * | ||||||||
| Precipitación acumulada anual (mm) | Bio12 | * | * | * | * | |||||||||||
| Precipitación del periodo más seco (mm) | Bio14 | * | * | * | * | * | * | * | * | |||||||
| Estacionalidad de la precipitación (%) | Bio15 | * | * | * | * | * | * | * | * | * | ||||||
| Precipitación del trimestre más lluvioso (mm) | Bio16 | * | * | * | * | |||||||||||
| Precipitación del trimestre más seco (mm) | Bio17 | * | * | * | * | * | * | * | * | * | * | * | * | * | ||
| Precipitación del trimestre más cálido (mm) | Bio18 | * | * | * | * | * | * | * | * | * | * | * | * | |||
| Precipitación del trimestre más frío (mm) | Bio19 | * | * | * | * | * | * | * | * | * | ||||||
| Orientación (°) | Ori | * | * | * | * | * | * | * | * | * | * | * | * | * | * | * |
| Pendiente (%) | Pen | * | * | * | * | * | * | * | * | * | * | * | * | * | * | * |
| Potencial Hidrogeno (0-14) | pH | * | * | * | * | * | * | * | * | * | * | * | * | * | * | * |
| Rocosidad (%) | Roc | * | * | * | * | * | * | * | * | * | * | * | * | * | * | * |
Especie: 1. A. religiosa, 2. P. devoniana, 3. P. douglasiana, 4. P. hartwegii, 5. P. herrerae, 6. P. lawsonii, 7. P. leiophylla, 8. P. martinezii, 9. P. maximinoi, 10. P. montezumae, 11. P. oocarpa, 12. P. pringlei, 13. P. pseudostrobus, 14. P. rzedowski y 15. P. teocote.
*Variable empleada en la modelación de la especie
Generación de modelos
La modelación de la idoneidad ambiental de las condiciones biofísicas donde se localizan las especies de estudio se realizó en el programa MaxEnt ver. 3.3.3 (Phillips et al., 2006) esto debido a la disponibilidad y naturaleza de los datos, los cuales fueron exclusivamente registros de presencia. Se generaron 10 réplicas para cada taxa analizado, empleando 75% de los registros y 25 % para la validación de los modelos (Phillips et al., 2006) y las variables ambientales seleccionadas en formato ascii.
Los criterios de modelación fueron 1000 iteraciones, un límite de convergencia de 10-5 (valor por omisión), un replicado interno por Bootstrap (Manzanilla-Quiñones, Delgado et al., 2019). Se desactivaron las casillas Extrapolate y Do clamping, esto con el fin de evitar extrapolaciones artificiales en los valores extremos de las variables y sobreajuste de los modelos (Elith et al., 2011) y se aplicó un umbral mínimo de presencia (Pearson et al., 2007). El formato de salida fue acumulativo, el cual proporciona un intervalo de valores de 0% a 100% (Phillips et al., 2006). El porcentaje de contribución de cada variable utilizada en la generación de los modelos fue determinado mediante la prueba de Jackknife (Phillips et al., 2006).
Calibración de modelos
La calibración de los modelos fue realizada a partir de la delimitación del área accesible M para las especies (diagrama de BAM), la cual ha sido definida como el área geográfica y ambiental que posee las condiciones ecológicas idóneas para el crecimiento, desarrollo y movilidad de las especies (Soberón y Peterson, 2005). La calibración de las áreas M se efectuó a partir de las subprovincias fisiográficas publicadas por Inegi (2001) donde se localizan los registros de presencia de las especies analizadas en Michoacán (Tabla 2).
Tabla 2 Áreas M empleadas en la calibración de los modelos de idoneidad ambiental para la toma de datos dendrocronológicos de las 15 especies analizadas.
| Especie | |||||||||||||||
| Subprovincia fisiográfica | |||||||||||||||
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 | |
| Llanuras y Sierras de Querétaro e Hidalgo | * | * | * | * | * | * | * | * | * | * | * | * | * | ||
| Mil Cumbres | * | * | * | * | * | * | * | * | * | * | * | * | * | ||
| Depresión del Balsas | * | * | * | ||||||||||||
| Sierras y Bajíos Michoacanos | * | * | * | * | * | * | * | * | * | ||||||
| Chapala | * | * | * | ||||||||||||
| Neovolcánica Tarasca | * | * | * | * | * | * | * | * | * | * | * | * | * | ||
| Escarpa Limítrofe del Sur | * | * | * | * | * | * | * | * | * | ||||||
| Depresión de Tepalcatepec | * | * | * | ||||||||||||
| Cordillera Costera del Sur | * | * | * | * | * | * | |||||||||
Especie: 1. A. religiosa, 2. P. devoniana, 3. P. douglasiana, 4. P. hartwegii, 5. P. herrerae, 6. P. lawsonii, 7. P. leiophylla, 8. P. martinezii, 9. P. maximinoi, 10. P. montezumae, 11. P. oocarpa, 12. P. pringlei, 13. P. pseudostrobus, 14. P. rzedowski y 15. P. teocote. *registros de presencia
Evaluación de los modelos
El desempeño estadístico de los modelos fue evaluado mediante las métricas obtenidas de la prueba del área bajo la curva AUC de ROC estándar (Receiver Operating Characteristic), los resultados del análisis Roc parcial (Peterson et al., 2008), así como los valores de la prueba de Z entre las proporciones AUC de Roc parcial el programa R versión 4.0 (R Core Team, 2020), estas últimas pruebas se consideran más robustas y de mayor precisión y confiabilidad que el AUC de ROC estándar.
Identificación de sitios potenciales
La estimación de los sitios potenciales para la colecta de datos dendrocronológico se llevó a cabo mediante la reclasificación de los valores continuos de la salida acumulativa de MaxEnt en tres categorías con intervalos iguales; baja, moderada y alta. Estas categorías representan la idoneidad ambiental de las condiciones biofísicas de los sitios donde crecen las especies de pináceas de Michoacán. Empleando el valor de la categoría de idoneidad ambiental alta se transformaron los modelos continuos a binarios (condiciones idóneas y no idóneas) en el programa ArcMap versión 10.3 (Environmental Systems Research Institute [ESRI], 2014).
La identificación de los sitios potenciales para la toma de datos de tipo dendrocronológicos se efectuó al localizar las áreas de mayor idoneidad ambiental predichas por los modelos en los municipios y áreas naturales protegidas de Michoacán. Asimismo, estos fueron divididos en tres zonas: oriente, centro y occidente. Por último, la elaboración y edición de los mapas de la distribución potencial de la idoneidad ambiental de las especies de pináceas de Michoacán se realizó con el programa ArcMap versión 10.3 (ESRI, 2014).
Resultados
Desempeño de los modelos
Los resultados de los modelos de idoneidad ambiental de las 15 especies evaluadas presentaron valores en el área bajo la curva (AUC) de 0.82 a 0.99, para los datos de entrenamiento; y de 0.87 a 0.99, para los datos de validación. Por otra parte, los resultados de las pruebas de Roc parcial oscilaron de 1.28 a 1.96, mientras que los valores del estadístico Z fueron de 32.55 a 4226.70 (p < 0.01). En la Tabla 3 se muestran los resultados de las pruebas de AUC, Roc parcial y Z, así como el número de réplica que mejor desempeño estadístico presentó para cada especie analizada.
Tabla 3 Resultados de las pruebas de AUC, Roc parcial y Z de los modelos de idoneidad ambiental para la toma de datos dendrocronológicos de las 15 especies analizadas.
| Especie | Réplica | AUC | Valor Roc parcial | Valor Z | |||
| Entrenamiento | Validación | ||||||
| A. religiosa | 5 | 0.97 | 0.97 | 1.92 | 1695.20* | ||
| P. devoniana | 7 | 0.93 | 0.92 | 1.73 | 304.63* | ||
| P. douglasiana | 2 | 0.97 | 0.96 | 1.82 | 381.12* | ||
| P. hartwegii | 8 | 0.99 | 0.99 | 1.91 | 333.80* | ||
| P. herrerae | 10 | 0.95 | 0.97 | 1.67 | 111.05* | ||
| P. lawsonii | 8 | 0.94 | 0.97 | 1.81 | 398.40* | ||
| P. leiophylla | 10 | 0.95 | 0.94 | 1.87 | 2076.50* | ||
| P. martinezii | 10 | 0.82 | 0.98 | 1.40 | 51.96* | ||
| P. maximinoi | 10 | 0.93 | 0.90 | 1.28 | 32.55* | ||
| P. montezumae | 10 | 0.94 | 0.94 | 1.84 | 885.93* | ||
| P. oocarpa | 4 | 0.95 | 0.92 | 1.73 | 307.90* | ||
| P. pringlei | 8 | 0.94 | 0.87 | 1.74 | 241.64* | ||
| P. pseudostrobus | 7 | 0.96 | 0.96 | 1.91 | 1858.80* | ||
| P. rzedowskii | 7 | 0.99 | 0.98 | 1.97 | 4226.70* | ||
| P. teocote | 3 | 0.95 | 0.90 | 1.71 | 303.26* | ||
*significativo; p<0.01
Variables relevantes
Las variables ambientales relevantes de los modelos de idoneidad ambiental fueron: oscilación anual de la temperatura (Bio7, 67.7%), pendiente (Pen, 10.6%) y precipitación del trimestre más seco (Bio17, 9.4%) para A. religiosa; precipitación del trimestre más lluvioso (Bio16, 54.7%), oscilación anual de la temperatura (Bio7, 14.9%) y precipitación del trimestre más frío (Bio19, 8.3%) para P. devoniana; precipitación del trimestre más lluvioso (Bio16, 44.2%), precipitación del trimestre más seco (Bio17, 25.2%) y pendiente (Pen, 22.5%) para P. douglasiana; temperatura promedio del trimestre más cálido (Bio10, 96%), precipitación del trimestre más frío (Bio19, 2%) y orientación (Ori, 0.9%) para P. hartwegii; rocosidad (Roc, 78.3%), pendiente (Pen, 11.6%) y orientación (Ori, 9.7%) para P. herrerae; precipitación del trimestre más seco (Bio17, 26.8%), pendiente (Pen, 21.1%) y estacionalidad de la precipitación (Bio15, 15.8%) para P. lawsonii; temperatura promedio del trimestre más lluvioso (Bio8, 37.2%), pendiente (Pen, 19%) y precipitación del trimestre más seco (Bio17, 15.8%) para P. leiophylla; estacionalidad de la precipitación (Bio15, 58.5%), precipitación del trimestre más seco (Bio17, 41.4%) y pendiente (Pen, 0.1%) para P. martinezii; precipitación del trimestre más lluvioso (Bio16, 59.1%), orientación (Ori, 15.8%) y oscilación anual de la temperatura (Bio7, 8.8%) para P. maximinoi; precipitación del trimestre más frío (Bio19, 31.1%), pendiente (Pen, 15.6%) y precipitación acumulada anual (Bio12, 13%) para P. montezumae; rocosidad (Roc, 23.4%), precipitación del trimestre más seco (Bio17, 14.8%) y precipitación del trimestre más cálido (Bio18, 14.7%) para P. oocarpa; precipitación del trimestre más frío (Bio19, 62%), oscilación anual de la temperatura (Bio7, 12.8%) y potencial hidrogeno (pH, 9.5%) para P. pringlei; temperatura promedio del trimestre más lluvioso (Bio8, 58.4%), precipitación del trimestre más frío (Bio19, 19.6%) y (Pen, 7.6%) para P. pseudostrobus; precipitación del periodo más seco (Bio14, 55.6), oscilación anual de la temperatura (Bio7, 31.8%) y rocosidad (Roc, 7.7%) para P. rzedowskii; y pendiente (Pen, 36.6%), orientación (Ori, 23.4%) y precipitación acumulada anual (Bio12, 13%) para P. teocote.
Áreas potenciales
Las superficies de idoneidad ambiental potencial por especie oscilaron de 753.97 ha (P. hartwegii) a 166 689.35 ha (P. lawsonii) (Tabla 4), distribuidas en tres zonas: 1) oriente, en la cual se distribuyen A. religiosa, P. douglasiana, P. hartwegii, P. leiophylla, P. maximinoi, P. pringlei, P. pseudostrobus y P. teocote (Fig. 2) y P. lawsonii y P. martinezii (Fig. 3); 2) centro, con A. religiosa, P. douglasiana, P. hartwegii y P. leiophylla (Fig. 4), P. montezumae y P. lawsonii (Fig. 5), P. maximinoi, P. pseudostrobus, P. teocote y P. martinezii (Fig. 6), P. oocarpa, P. pringlei y P. devoniana (Fig. 7); y 3) occidente, con P. rzedowskii, P. herrerae, P. oocarpa, P. douglasiana y P. leiophylla (Fig. 8).
TABLA 4 Superficies estimadas de idoneidad ambiental potencial para la toma de datos dendrocronológicos de las 15 especies de estudio.
| Especie | Superficie (ha) |
| A. religiosa | 6591.92 |
| P. douglasiana | 18 672.65 |
| P. devoniana | 75 600.75 |
| P. hartwegii | 753.97 |
| P. herrerae | 8874.07 |
| P. lawsonii | 71 687.87 |
| P. leiophylla | 75 954.59 |
| P. martinezii | 99 723.64 |
| P. maximinoi | 84 337.52 |
| P. montezumae | 21 639.73 |
| P. oocarpa | 48 317.33 |
| P. pringlei | 166 689.35 |
| P. pseudostrobus | 51 868.08 |
| P. rzedowskii | 3003.36 |
| P. teocote | 37 003.06 |

Figura 2 Distribución espacial de la idoneidad ambiental potencial para la toma de datos den-drocronológicos de A. religiosa, P. douglasiana, P. hartwegii, P. leiophylla, P. maximinoi, P. pringlei, P. pseudostrobus y P. teocote en la zona oriente de Michoacán.
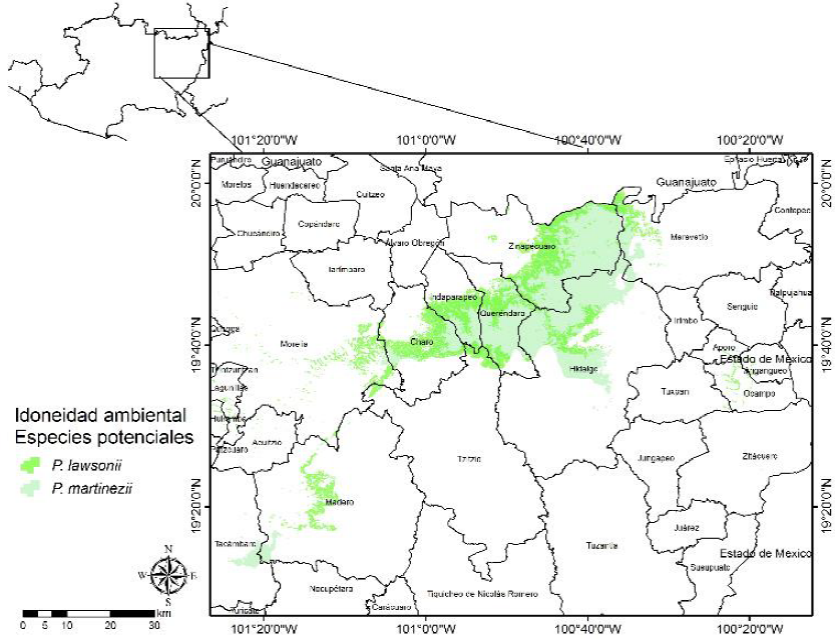
Figura 3 Distribución espacial de la idoneidad ambiental potencial para la toma de datos dendrocronológicos de P. lawsonii y P. martinezii en la zona oriente de Michoacán.

Figura 4 Distribución espacial de la idoneidad ambiental potencial para la toma de datos dendrocronológicos de A. religiosa, P. douglasiana, P. hartwegii y P. leiophylla en la zona centro de Michoacán.

Figura 5 Distribución espacial de la idoneidad ambiental potencial para la toma de datos dendrocro-nológicos de P. montezumae y P. lawsonii en la zona centro de Michoacán.

Figura 6 Distribución espacial de la idoneidad ambiental potencial para la toma de datos dendrocronológicos de P. maximinoi, P. pseudostrobus, P. teocote y P. martinezii en la zona centro de Michoacán.
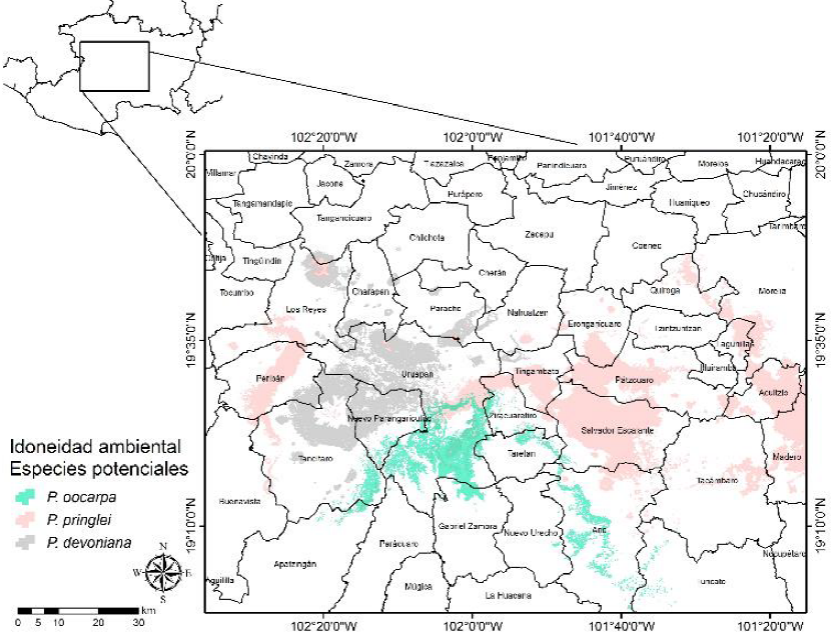
Figura 7 Distribución espacial de la idoneidad ambiental potencial para la toma de datos dendrocronológicos de P. oocarpa, P. pringlei y P. devoniana en la zona centro de Michoacán.

Figura 8 Distribución espacial de la idoneidad ambiental potencial para la toma de datos dendrocronológicos de P. rzedowskii, P. herrerae, P. oocarpa, P. douglasiana y P. leiophylla en la zona occidente de Michoacán.
La mayoría de las áreas con mayor idoneidad ambiental potencial para la toma de datos dendrocronológicos se localizaron en los municipios de Tancítaro (3916.77 ha), Angangueo (737.26 ha) y Senguio (583.10 ha) para A. religiosa; Uruapan (25 694.74 ha), Tancítaro (18 521.42 ha) y Nuevo Parangaricutiro (12 199 ha) para P. devoniana; Uruapan (5057.26 ha), Nuevo Parangaricutiro (3644.85 ha) y Tancítaro (2429.80 ha) para P. douglasiana; Tancítaro (624.84 ha) para P. hartwegii; Coalcomán (4064.44 ha), Arteaga (2624.09 ha) y Tumbiscatío (372.79 ha) para P. herrerae; Zinapécuaro (10 896.33 ha), Uruapan (10 343.01 ha) y Charo (7428.46 ha) para P. lawsonii; Tancítaro (12 335.42 ha), Nuevo Parangaricutiro (8090.88 ha) y Pátzcuaro (5940.82 ha) para P. leiophylla; Zinapécuaro (33 441.78 ha), Ciudad Hidalgo (29 494.91 ha) y Queréndaro (14 283.37 ha) para P. martinezii; Uruapan (28 972.17 ha), Tancítaro (15 777.08 ha) y Nuevo Parangaricutiro (7700.31 ha) para P. maximinoi; Tancítaro (9061.95 ha), Uruapan (4392.34 ha) y Nuevo Parangaricutiro (3590.38 ha) para P. montezumae; Uruapan (22 231.91 ha), Ario de Rosales (6268.71 ha) y Tancítaro (3069.06 ha) para P. oocarpa; Salvador Escalante (31 634.55 ha), Pátzcuaro (16 614.15 ha) y Acuitzio (11 195.98 ha) para P. pringlei; Tancítaro (9864.91 ha), Uruapan (7954.69 ha) y Nuevo Parangaricutiro (3935.59 ha) para P. pseudostrobus; Coalcomán (2056.26 ha) y Aguililla (845.57 ha) para P. rzedowskii; y Morelia (5975.23 ha), Uruapan (3733.25 ha) y Nuevo Parangaricutiro (1172.59 ha) para P. teocote. Los municipios de Tancítaro, Uruapan y Nuevo Parangaricutiro fueron los que mayor número de especies potenciales presentaron para la toma de datos dendrocronológicos (Tabla 5).
Tabla 5 Municipios con mayor número de especies potenciales para la toma de datos dendrocronológicos en Michoacán.
| Municipio | Especies |
| Tancítaro | A. religiosa, P. devoniana, P. douglasiana, P. hartwegii, P. leiophylla, P. maximinoi, P. montezumae, P. oocarpa, P. pringlei y P. pseudostrobus |
| Uruapan | P. devoniana, P. douglasiana, P. lawsonii, P. maximinoi, P. montezumae, P. oocarpa, P. pseudostrobus, P. pringlei y P. teocote |
| Nuevo Parangaricutiro | P. devoniana, P. douglasiana, P. leiophylla, P. maximinoi, P. montezumae, P. pseudostrobus, P. pringlei y P. teocote |
La Reserva de la Biosfera Mariposa Monarca presentó una superficie de idoneidad ambiental potencial para la toma de datos dendrocronológicos de 1690.34 ha para A. religiosa, 1235.02 ha para P. maximinoi, 162.2 ha para P. pseudostrobus y 89.58 ha para P. hartwegii (Fig. 9). Mientras que el Pico de Tancítaro (PDT) presentó una superficie de idoneidad ambiental potencial para la toma de datos dendro-cronológicos de 17906.7 ha para P. devoniana, 13279.19 ha para P. leiophylla, 11289.47 ha para P. montezumae, 9395.29 para P. pseudostrobus, 4415.57 ha para A. religiosa, 817.58 para P. teocote, 662.94 para P. maximinoi, 569.38 ha para P. hartwegii, 198.11 ha para P. pringlei; debido al alto traslape observado entre estas especies, se optó por generar un mapa de riqueza de pináceas para el Pico de Tancítaro (Fig. 10).
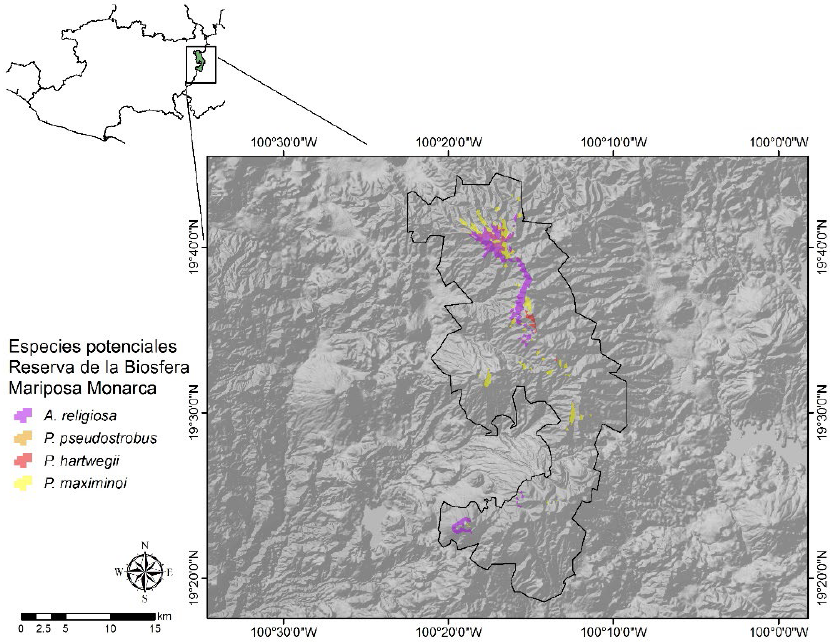
Figura 9 Distribución espacial de la idoneidad ambiental potencial para la toma de datos dendrocronológicos de A. religiosa, P. pseudostrobus, P. hartwegii y P. maximinoi en la Reserva de la Biosfera Mariposa Monarca.

Discusión
Desempeño de los modelos
Acorde con los resultados de las pruebas de AUC (> 0.84), Roc parcial (> 1.28) y de Z (p < 0.01), los modelos de idoneidad ambiental presentaron un desempeño estadístico que osciló de bueno a excelente (Peterson et al., 2008; Martínez-Méndez et al., 2016) por lo tanto, estos se consideraron como modelos confiables para los propósitos planteados en esta investigación.
Variables relevantes
Las variables que más influyeron en la generación de los modelos fueron del tipo bioclimáticas: precipitación del trimestre más lluvioso (Bio16), precipitación del trimestre más seco (Bio17), precipitación del trimestre más frío (Bio19), oscilación de la temperatura anual (Bio7) y temperatura promedio del trimestre más lluvioso (Bio8), las cuales presentaron un alto porcentaje de relevancia en 13 de las 15 especies analizadas. Resultados similares han sido observados por Martínez-Méndez et al. (2016), quienes hallaron que las variables Bio19, Bio17, Bio16 y Bio7 presentaron un mayor grado de importancia en el modelado de las áreas de distribución de las especies de oyamel registradas para México. Por su parte Martínez-Sifuentes, Villanueva-Díaz, Manzanilla-Quiñones, et al. (2020))señalaron a Bio19 y Bio7 como las variables que más influyeron en el modelado de nicho ecológico de P. greggii en México.
A su vez Cruz-Cárdenas et al. (2016) mencionaron a Bio5, Bio6, Bio7 y Bio17 como las variables que mayor ponderación presentaron en la elaboración de los modelos de distribución potencial de 12 especies de pináceas de Michoacán. Acorde con estos resultados, la combinación de las variables bioclimáticas indicaría una preferencia de estas especies de coníferas mexicanas a climas templados y húmedos; la precipitación y la temperatura registrada durante primavera-verano fueron las más relevantes en el crecimiento radial de dichas especies.
Los estudios dendrocronológicos realizados en México coinciden en que la variabilidad de la precipitación y la temperatura que se presenta durante las estaciones de primavera-verano influye en la etapa de reactivación celular del crecimiento radial de las pináceas mexicanas (Biondi et al., 2005; Villanueva-Díaz et al., 2015; Manzanilla-Quiñones et al., 2020; Carlón et al., 2021). Esta variabilidad climática es similar a los valores de las variables Bio16, Bio17, Bio19 y Bio8 para 13 de 15 de las especies analizadas en este estudio. Por lo que, acorde con las variables relevantes identificadas para cada especie, el modelado de idoneidad ambiental de las condiciones biofísicas serviría como una herramienta auxiliar en la identificación de sitios potenciales para la toma de datos dendrocronológicos en Michoacán y en otras regiones geográficas de la República mexicana.
Diversos estudios efectuados sobre modelado de distribución potencial de especies han demostrado que su aplicabilidad en temáticas ajenas a cuestiones biológicas, ecológicas y geográficas como la predicción de brotes de enfermedades e invasión de especies han presentado resultados sumamente confiables (Qiao et al., 2017; Yañez-Arenas et al., 2017). Por lo tanto, se esperaría que el modelado de las condiciones biofísicas presentara resultados buenos en la predicción de sitios potencialmente idóneos para la posterior toma de datos dendrocronológicos. Sin embargo, esto último necesitaría ser corroborado mediante la toma y el análisis de muestras en campo.
Identificación de áreas potenciales
La mayoría de los estudios dendrocronológicos realizados en México se basan en la experiencia acumulada del investigador y en el conocimiento de los dueños de los predios forestales de los sitios por muestrear, descartando el análisis espacial previo de las condiciones biofísicas donde crecen los árboles.
En general, los municipios de Tancítaro, Uruapan y Nuevo Parangaricutiro fueron los sitios que presentaron el mayor número de especies y áreas potencialmente idóneas para la toma de datos dendrocronológicos. La zona es considerada como un área prioritaria para la conservación y conectividad del paisaje ecológico de P. pseudostrobus y P. montezumae (Molina et al., 2019), así como para especies de bosque mesófilo de montaña como A. religiosa y P. hartwegii (Correa Ayram et al., 2017), por lo que la realización de estudios dendrocronológicos ayudaría en el análisis histórico de la variabilidad climática y en la identificación de eventos ambientales ocurridos en la región de la Meseta Purépecha de Michoacán.
A escala nacional, el único estudio en el cual se incorporó el análisis espacial de las características ambientales fue el realizado por Carlón et al. (2015) en la RBMM para A. religiosa y P. pseudostrobus. En esos estudios se identificaron los sitios potenciales, en los cuales posteriormente se tomaron muestras en campo para analizarlas en el laboratorio. Se concluyó que la predicción del análisis espacial de las condiciones biofísicas presentó resultados fiables en la identificación de sitios potenciales para la toma de datos dendrocronológicos. Acorde con lo mencionado por Carlón et al. (2015) y con los resultados obtenidos en esta investigación, la RBMM presenta condiciones ambientales favorables para la toma de datos dendrocronológicos de las especies A. religiosa, P. pseudostrobus, P. maximinoi y P. hartwegii, cuyos sitios potenciales se localizan en el norte y centro de la RBMM.
Investigaciones dendrocronológicas realizadas en el Pico de Tancítaro por Cerano-Paredes et al. (2014) para A. religiosa indican que es sensible a los cambios en la precipitación registrada durante enero-mayo, lo cual es similar a la variable precipitación del trimestre más seco (Bio17) registrado en este estudio para la misma especie.
Por su parte, Carlón et al. (2021) observaron una respuesta fisiológica negativa de la temperatura máxima promedio, registrada durante las estaciones de primavera-verano, en el crecimiento radial anual de P. hartwegii de las partes altas del Pico de Tancítaro. Resultado que es similar a la principal variable (temperatura promedio del trimestre más cálido, Bio10) que más influye en la idoneidad ambiental de esa misma especie en este estudio.
Además de A. religiosa y P. hartwegii analizadas por Cerano-Paredes et al. (2014) y Carlón et al. (2021) el Pico de Tancítaro presenta superficies de idoneidad potencial altas para la toma de datos dendrocronológicos de P. devoniana, P. teocote, P. leiophylla, P. maximinoi, P. montezumae, P. pringlei y P. pseudostrobus. Dichas especies no han sido analizadas desde una perspectiva dendrocronológica para la zona y/o el área natural protegida y esa información serviría para conocer su respuesta fisiológica a los cambios en el clima, incendios forestales, ataques de plagas y la influencia de fenómenos océano-atmosféricos de circulación global.
Conclusiones
Mediante la aplicación de los modelos de distribución de especies fue posible modelar las condiciones biofísicas de los sitios donde crecen los árboles de 15 especies de pináceas del estado de Michoacán.
Las variables ambientales identificadas en este estudio son similares a las respuestas fisiológicas del crecimiento señaladas por diversos estudios dendrocronológicos.
Los municipios de Tancítaro, Uruapan y Nuevo Parangaricutiro fueron los que presentaron el mayor número (12) de especies potenciales para la toma de datos dendrocronológicos en Michoacán.
La presente propuesta pretende emplear el uso de los modelos de distribución potencial como una herramienta auxiliar en la identificación de sitios potenciales para la toma de datos dendrocronológicos. Sin embargo, esto último necesitaría ser corroborado mediante la toma y el análisis de muestras en campo.
La aplicabilidad del presente método puede extrapolarse hacia otras especies arbóreas y regiones geográficas de México.

