











 text new page (beta)
text new page (beta) Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO 
 Permalink
PermalinkIntroducción
México tiene aproximadamente 138 millones de hectáreas de vegetación, lo que representa 70% de su territorio; de esa superficie, 17% corresponde a bosques templados (Comisión Nacional Forestal [Conafor], 2014). La función ecológica de este tipo de vegetación incluye la regulación del clima al actuar como sumideros de gases de efecto invernadero (Food and Agriculture Organization of the United Nations [FAO], 2018). La excesiva acumulación de estos gases en la atmósfera está provocando cambios en el clima, evidencia de ello es el aumento de temperatura de 0.87 °C registrado en el periodo de 2006 a 2015; de continuar esta tendencia, se podrían alcanzar los 1.5 °C para al año 2040 (Allen et al., 2018). La alteración de la composición del ecosistema por la extracción de especies, la conversión de uso de suelo, y los incendios forestales, entre otros, provocan debilitamiento y estrés en los árboles, lo que reduce su eficiencia reguladora y favorece la incidencia de plagas forestales (Salinas et al., 2010).
Los insectos descortezadores son parte natural de la estructura y del funcionamiento de los bosques (Wood, 1982). No obstante, existe evidencia clara sobre la magnitud, frecuencia e intensidad de los brotes de descortezadores por efectos directos o indirectos del clima (Pureswaran, Roques y Battisti, 2018). En el caso de especies como Dendroctonus rufipennis Kirby y Dendroctonus ponderosae Hopkins, incrementarían su población bajo escenarios de calentamiento global (Bentz et al., 2010). Otros investigadores demuestran que con una temperatura de 1.6 °C, los descortezadores podrían: aumentar el número de generaciones por año (Logan y Powell, 2001; Waring et al., 2009), ampliar su distribución y modificar sus hábitos (Régnière, 2009; Kocmánková et al., 2010), además de incrementar su tasa de supervivencia al ubicarse en mayores latitudes y altitudes (Pureswaran et al., 2018).
Los modelos de distribución de especies relacionan las condiciones del ambiente con su posible distribución geográfica (Guisan y Zimmermann, 2000). A través del diagrama «BAM» pueden determinarse los factores causales de los cambios de distribución de una especie con base en: condiciones bióticas favorables («B»), condiciones abióticas («A») y las variables accesibles a la dispersión de la especie desde la región de su origen histórico («M») (Soberón y Nakamura, 2009; Soberón, Osorio-Olvera y Peterson, 2017). Para modelar la distribución de especies se han desarrollado distintos algoritmos tales como: BIOCLIM (Busby, 1991), GARP (Stockwell, 1999) y Maxent (Phillips, Dudík y Schapire, 2004). Este último es de los más robustos y el que mejor predice la idoneidad climática para las especies (Elith et al., 2006); también puede incluir un gran número de variables escenopoéticas (bioclimáticas) de diferente índole indicadas en el diagrama de «BAM» (Soberón et al., 2017) y que se han utilizado para modelar la distribución de las especies (Beaumont, Hughes y Poulsen, 2005).
El desarrollo de modelos en E.E.U.U. se ha enfocado en D. ponderosae (Régnière, 2009), Dendroctonus frontalis Zimmermann y D. ponderosae (Williams y Liebhold, 2002) y en Dendroctonus valens LeConte (Maldonado et al., 2015). Mientras que, en México, existe un trabajo relevante que modeló con BIOCLIM la distribución actual de 12 especies de descortezadores, entre ellas Dendroctonus mexicanus Hopkins, revelando que esta es la especie con más amplia distribución en los bosques de México y coloniza hasta 20 especies de pino (Salinas et al., 2010). Sosa et al. (2018), modelaron la distribución potencial de descortezadores a nivel género, mientras que Mendoza, Salinas, Olivo y Zúñiga (2011) hicieron lo propio con Dendroctonus rizophagus Thomas & Bright, en la Sierra Madre Occidental. Por los impactos económicos, ecológicos y daños irreversibles que ocasiona a los ecosistemas, D. mexicanus es considerada la especie de descortezador más importante en México (Cibrián et al., 1995); de aquí la importancia de conocer su distribución actual y potencial.
A través del tiempo, se ha incrementado el número de registros de presencia de las diferentes especies, ampliando el conocimiento de sus límites de distribución (Armendáriz, Torres, López, Villa y Zúñiga, 2012), han mejorado los algoritmos de modelación (Cobos, Peterson, Barve y Osorio, 2019) y se han creado nuevas superficies bioclimáticas y modelos de circulación general (MCG´s) (Karger et al., 2017), lo que da la pauta para generar más estudios sobre la distribución de especies.
Objetivos
Los objetivos de este estudio fueron modelar la distribución actual y potencial de D. mexicanus, bajo diferentes escenarios de cambio climático para el año 2050, usando cinco MCG´s: MIROC5, HadGEM2-AO, CSIRO-Mk3-6-0, CSM4 y CNRM-CM5 y dos «trayectorias de concentración repre-sentativas» (RCP) 4.5 y 8.5, así como conocer las variables climáticas asociadas a su presencia, los impactos del cambio climático sobre la distribución actual y futura de esta especie y posibles riegos de invasión. Se hipotetiza que esta especie es sensible al clima y por tanto su distribución geográfica será modificada al 2050.
Materiales y métodos
Área de estudio
El estudio se llevó a cabo en el área de distribución actual de D. mexicanus, la cual se ubica en bosques de coníferas de México, que abarcan una superficie de 164 422.7 km2 (FAO, 2018), con predominancia de especies del género Pinus, Quercus, Juniperus y Abies (Flores y Gerez, 1994). Se localiza en las principales serranías montañosas, Sierra Madre Oriental (SMO), Occidental (SMOc), Eje Neolvolcánico Transversal (ENT), en altitudes de 50 m a 3500 m snm, temperaturas medias anuales entre 10 °C y 20 °C y precipitación de 600 mm a 1000 mm (Rzedowski, 1978).
Obtención registros y variables bioclimáticas
Los registros de presencia de D. mexicanus se obtuvieron del portal GBIF (Global Biodiversity Information Facility), de artículos científicos y de colectas propias. Los datos se depuraron con base en lo sugerido por Cobos, Jiménez, Nuñez, Romero y Simoes (2018), eliminando registros duplicados, de baja precisión, fuera del límite de la distribución de la especie (en altitud, latitud y longitud), con errores de identificación etc.; permitiendo al menos un registro por celda de las capas ambientales. La autocorre-lación espacial entre los registros se eliminó en R usando una distancia de 5 km a través del paquete spThin (Aiello, Boria, Radosavljevic, Vilela y Anderson, 2015). Las variables bioclimáticas (~1 km2 de resolución) utilizadas fueron las propuestas por Karger et al. (2017), excluyendo de las 19, Bio 8, Bio 9, Bio 18 y Bio 19 (Tabla 1) para disminuir la incerti-dumbre y por combinar información de precipitación y tempera-tura en la misma capa (Escobar, Lira, Medina y Peterson, 2014).
Tabla 1 Variables bioclimáticas utilizadas para generación de modelos de distribución actual y potencial de Dendroctonus mexicanus Hopkins.
| Variable | Descripción |
| Bio 1 | Temperatura media anual (°C) |
| Bio 2 | Intervalo medio diurno (media por mes [temp. máx. - temp. mín.]) |
| Bio 3 | Isotermalidad (Bio 2 / Bio 7)*(100) |
| Bio 4 | Estacionalidad de la temperatura (desviación estándar * 100) |
| Bio 5 | Temperatura máxima del mes más cálido (°C) |
| Bio 6 | Temperatura mínima del mes más frío (°C) |
| Bio 7 | Intervalo de temperatura anual (Bio 5 - Bio 6) |
| Bio 10 | Temperatura media del 1/4 anual más cálido (°C) |
| Bio 11 | Temperatura media del 1/4 anual más frío (°C) |
| Bio 12 | Precipitación anual (mm) |
| Bio 13 | Precipitación del mes más húmedo (mm) |
| Bio 14 | Precipitación del mes más seco (mm) |
| Bio 15 | Temporalidad de la precipitación (coeficiente de variación, %) |
| Bio 16 | Precipitación de 1/4 anual más húmedo (mm) |
| Bio 17 | Precipitación de 1/4 anual más seco (mm) |
Área de calibración y espacio ambiental
La delimitación del área de calibración, denotada por «M» en el diagrama de «BAM», es fundamental en la modelación de nichos y distribución de especies, así como en la genera-ción y transferencia del modelo (Soberón et al., 2017). Esta delimitación se realizó en ArcMap 10.1, aplicando un buffer de 70 km de radio al total de los registros (n = 283), identi-ficando “outliers” tanto en el espacio geográfico («G») como en el espacio ambiental («E»), hasta definir «M», aplicando la llamada «dualidad de Hutchinson» (Colwell y Rangel, 2009).
Selección de variables bioclimáticas
Las variables bioclimáticas se limitaron al área de calibración; para seleccionarlas, se realizó una modelación previa en Maxent 3.3.3k, con los parámetros predeterminados del programa (Phillips, Anderson y Schapire, 2006), usando todos los registros (50% para entrenamiento y el resto para validación) para incluir toda la geografía y climas actuales ocupados por D. mexicanus. La importancia de cada variable se evaluó mediante pruebas jackknife, implementadas en Maxent (Phillips et al., 2006). En ArcMap 10.1, se realizó correlación de Pearson entre variables bioclimáticas en formato raster. Finalmente, la selección de las variables se realizó considerando la biología de la especie, el valor de importancia de la variable y el aporte individual y en conjunto hacia el modelo, además de elegir aquellas no correlacionadas entre sí (r < 0.8) (Elith et al., 2006). Tres grupos de variables fueron probados para modelar la distribución de D. mexicanus. Para evitar la sobrestimación del modelo y representaciones erróneas de los límites de la realidad biológica (Beaumont et al., 2005; Cobos et al., 2019), cada grupo se integró de cinco variables.
Calibración, creación y evaluación del modelo
La calibración, creación y evaluación del modelo se realizó en Maxent, dentro de R Core Team (2019), utilizando el paquete kuenm (Cobos et al., 2019), el cual genera un gran número de modelos candidatos, evaluándolos de acuerdo con: significancia estadística, ROC parcial (curvas operadas por el receptor), tasas de omisión, criterio de información Aikaike o complejidad del modelo (AICc) (Phillips et al., 2006; Peterson, Papeş y Soberón, 2008) y pruebas de jackknife (Cobos et al., 2019). Se empleó ROC parcial para determinar el ajuste del modelo, excluyendo el estadístico AUC (Area Under Curve) ya que no se recomienda porque su predicción no ocupa todo el espectro del área de la curva ROC (Peterson et al., 2008).
Transferencia de modelos y riesgo de extrapolación
Los modelos calibrados se transfirieron en tiempo (al año 2050) y espacio (todo el mundo) a través de tres enfoques: i) sin extrapolación, ii) con extrapolación y iii) con clamping (Escobar et al., 2014), utilizando cinco MCG´s: MIROC5, HadGEM2-AO, CSIRO-Mk3-6-0, CSM4 y CNRM-CM5, bajo dos RCP´s: 4.5 y 8.5; cuyo análisis utilizó las medias de cada uno de ellos. El formato de salida fue de tipo logístico, donde los modelos resultantes (mapas) representan valores de idoneidad (0 - 1) de la especie (Phillips et al., 2006). El riesgo de extrapolación del modelo se realizó usando ExDet (Mesgaran, Cousens y Webber, 2014) basada en distancias de Mahalanobis, dentro del paquete ntbox de R (Osorio, Narayani, Soberón y Falconi, 2016).
Idoneidad actual y futura
Los mapas de idoneidad actual de D. mexicanus se estratificaron en tres categorías: alta, media y baja, usando el método de Dalenius y Hodges (1959), consistente en formar estratos lo más homogéneos posible (varianza mínima intra estrato y máxima inter estrato), buscando inicialmente qué distribución estadística se ajustó a los datos de idoneidad, esta fue obtenida en R con el paquete fitdistrplus (Delignette-Muller y Dutang, 2015) e integrada en Mathematica5, versión 5.0.0.0 y, posteriormente, se obtuvo la superficie por entidad federativa y por estrato. Los mapas de idoneidad actual y futura de D. mexicanus se transformaron a mapas binarios tomado como base el umbral de mínima presencia, después se analizaron mediante álgebra de rasters, para determinar: i) nuevas áreas idóneas en 2050, ii) sobreposición de áreas (actual y futura) y iii) áreas idóneas actuales que ya no lo son en 2050.
Resultados
Se obtuvieron 283 registros totales de ocurrencia de D. mexicanus, de los cuales, 67 se utilizaron para modelación. En «M», la Bio 1 se correlacionó positivamente (0.20 < r < 0.95, p ≤ 0.05) con la mayoría de las variables, contrario a Bio 2, Bio 3 y Bio 15, que no se correlacionaron con las variables evaluadas. En «E», también llamado «espacio ambiental o modelo de nicho» (Fig. 1), se observan registros dispersos, indicando probablemente que D. mexicanus pudiera no encontrarse en equilibrio, o quizá a la falta de monitoreo de esta especie en ciertas regiones climáticas. En este modelo de nicho, la especie se registra desde -1.9 hasta 33.6 °C, con medias que van desde 9 °C hasta 21 °C y desde 450 mm hasta 1755 mm.

Distribución actual de Dendroctonus mexicanus Hopkins
Se generaron un total de 1479 modelos candidatos por kuenm (Cobos et al., 2019), reflejando todas las combinaciones de 17 multiplicadores de regularización, 29 tipos de respuesta de modelado y 3 distintos conjuntos de variables ambientales. Acorde con los estadísticos de selección, el modelo con mayor robustez fue el tipo de respuesta lineal+cuadrático (lq), con un regulizador 0.2 (Tabla 2) y el conjunto de variables Bio 3, Bio 7, Bio 10, Bio 11 y Bio 17. Los resultados muestran buen ajuste de los modelos de D. mexicanus, con valores de ROC parcial de 0.9314 (modelo de calibración) hasta 0.9485 (transferencia al 2050, RCP 4.5). La tasa de AUC fluctuó desde 1.8629 (calibración) hasta 1.8971 (transferencia al 2050, RCP 8.5).
Tabla 2 Estadísticas generales de los modelos candidatos de Dendroctonus mexicanus Hopkins.
| Criterio | Número de modelos |
| Total de modelos candidatos | 1479 |
| Modelos estadísticamente significantes (mES) | 1463 |
| Modelos que cumplen con el criterio tasa de omisión (mTO) | 15 |
| Modelos que cumplen con el criterio AICc (mAIC) | 1 |
| Número de mES y mTO | 15 |
| Número de mES y mAIC | 1 |
| Número de mES, mTO y mAIC | 1 |
Independientemente del enfoque de la modelación (con extrapolación, sin extrapolación y clamping), los datos de idoneidad del modelo de distribución actual de D. mexicanus se ajustaron a una distribución Gamma. Los valores de los parámetros de ´forma´ de la distribución Gamma fueron: 0.6003 y 2.7178; 0.6158 y 2.1976; 0.5373 y 1.6480 para el RCP 4.5 y 8.5, y los valores de idoneidad fueron desde 0.0001 hasta 0.8309, 0.8052 y 0.8357, respectivamente al enfoque. La idoneidad alta estimada para D. mexicanus, acorde con el enfoque sin extrapolación (Fig. 2a), representa 11.2% del total «M». Además, se distribuye en forma continua, principalmente en el Eje Neolvolcánico Transversal, destacando que en esta parte del país la idoneidad pasa directamente del estrato alto al bajo (Fig. 2a). La idoneidad media se observa hacia mayores latitudes, constituye 12.7% de «M», pero es discontinua. La modelación con los enfoques extrapolación (Fig. 2b) y clamping (Fig. 2c) muestran patrones geográficos similares al hacerlo sin extrapolación.
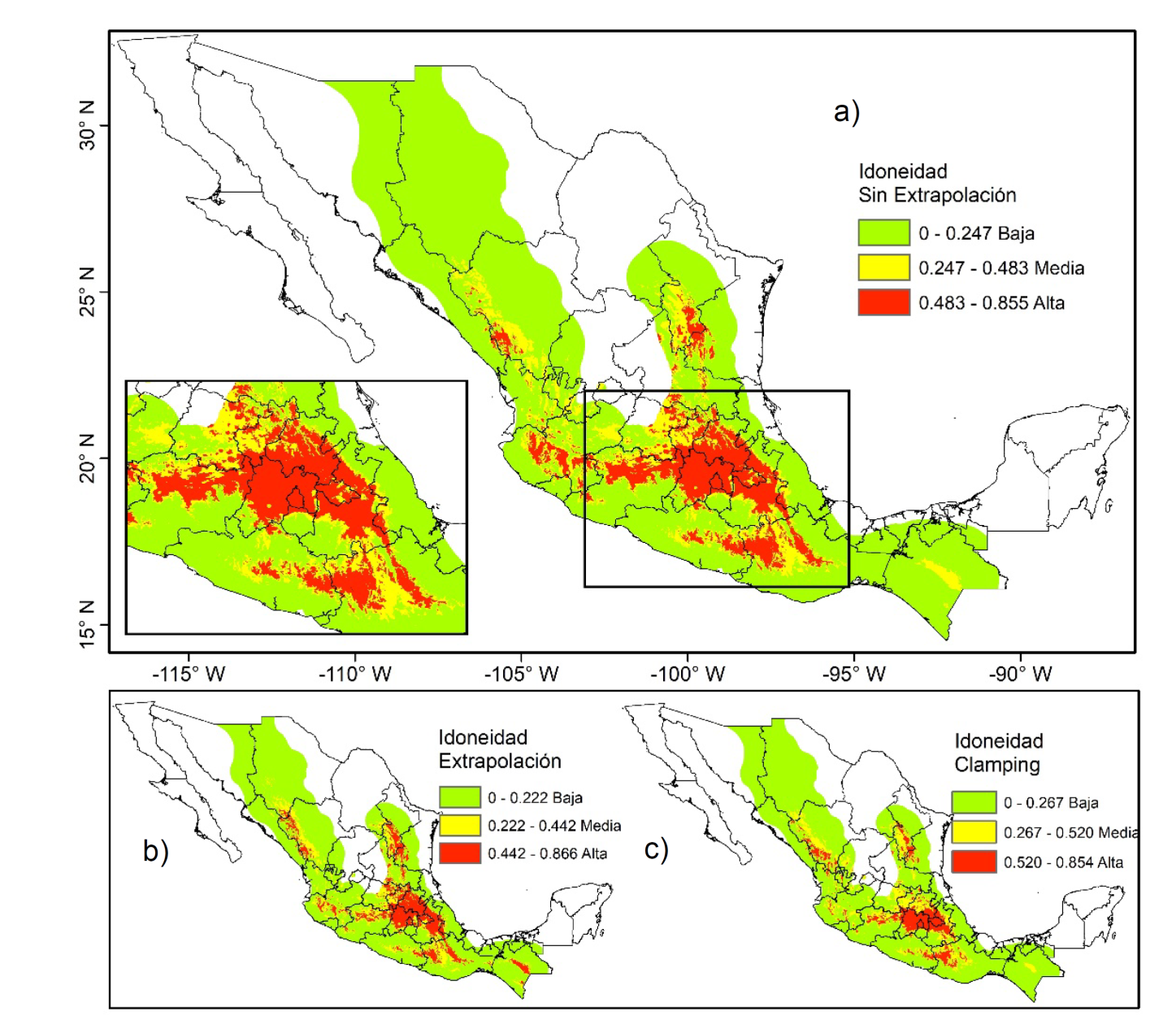
Figura 2 Áreas de idoneidad actual de Dendroctonus mexicanus Hopkins, estratificada en tres niveles (alto, medio y bajo) mediante la técnica de varianza mínima al interior de cada estrato y máxima entre ellos, en tres enfoques de modelación: sin extrapolación (a), con extrapolación (b) y con clamping (c).
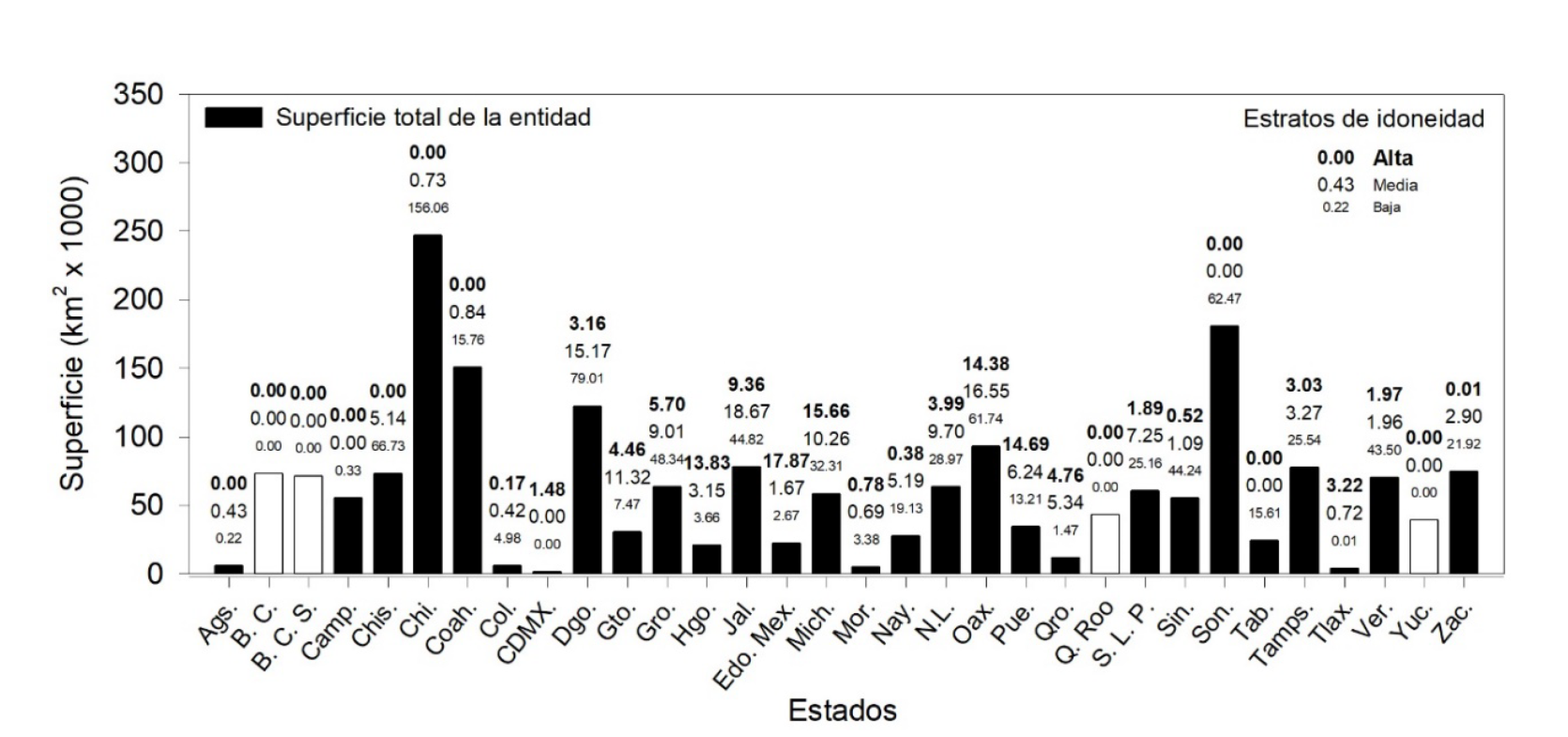
En México, el área de idoneidad alta de D. mexicanus es de 121 310.7 km2; la idoneidad media es de 137 710.4 km2 y la baja de 828 710.2 km2. Las entidades: Estado de México (17 879.7), Michoacán (15 669.8), Puebla (14 693.1), Oaxaca (14 380.1) e Hidalgo (13 830 km2), representan más de 63% de la superficie de idoneidad alta estimada en «M» (Fig. 2a y 3). La idoneidad media ocurre en: Jalisco (18 672.47), Oaxaca (16 558.64) y Durango (15 173.74 km2), equivalente a 37% del total de este estrato (Fig. 2a y 3).
Perfil bioclimático actual y futuro de Dendroctonus mexicanus Hopkins
Las variables bioclimáticas que mejor explican la distribución actual de D. mexicanus son las derivadas de temperatura: la media del 1/4 anual más cálido (Bio 10) y el intervalo de temperatura anual (Bio 7), contribuyendo con 76.6 y 12.5% respectivamente. La contribución de la Bio 10 es ligeramente mayor al modelar al 2050 (~ 80% en ambos RCP´s), el resto de las variables (Bio 7, Bio 17, Bio 3 y Bio 11) aportan al modelo solo 23% (Tabla 3).
Tabla 3 Contribución de las variables bioclimáticas al modelo actual, transferido al 2050 en dos trayectorias de concentración representativas RCPS´s 4.5 y 8.5 y perfil bioclimático de Dendroctonus mexicanus Hopkins.
| Contribución al modelo (%) | Perfil bioclimático (mínimo y máximo) | |||||
| Variable | Actual | RCP 4.5¥ | RCP 8.5¥ | Actual¶ | RCP 4.5¶ | RCP 8.5¶ |
| Bio 10 | 76.6 | 79.9 ± 1.2 | 80.58 ± 0.6 | 110.0 - 283.0 | 126.6 - 300.0 | 130.4 - 303.4 |
| Bio 7 | 12.5 | 11.1 ± 0.8 | 10.8 ± 0.8 | 134.0 - 275.0 | 153.8 - 297.6 | 141.4 - 279.4 |
| Bio 17 | 5.5 | 4.4 ± 0.1 | 4.32 ± 0.3 | 11.0 - 110.0 | 9.0 - 100.6 | 7.6 - 97.0 |
| Bio 3 | 2.3 | 3.1 ± 0.9 | 2.9 ± 0.6 | 442.0 - 590.0 | 53.0 - 69.0 | 48.2 - 63.8 |
| Bio 11 | 3.1 | 1.5 ± 0.4 | 1.4± 0.5 | 110.0 - 283.0 | 65.6 - 237.4 | 69.8 - 240.8 |
¥Promedio ± desviación estándar de cinco modelos de circulación general.
¶En las mismas unidades indicadas en tabla 1.
La variable de mayor contribución (Bio 10) muestra una distribución bimodal (Fig. 4a) en ambos periodos, actual y al 2050, y RCPS´s. Sesenta por ciento de los registros de D. mexicanus se ubican actualmente en 18 °C, acorde con la mediana1+[1.33x log 10 (n)] propuesto por (Sturges, 1926). Se estima que, en las mismas zonas de registro, la temperatura aumente 1.6 °C y 2.0 °C en el año 2050 en RCP 4.5 y 8.5, respectivamente. En Bio 7, 78.8% de los registros se ubican entre 17 °C y 19 °C (Fig. 4b), el desplazamiento al 2050 es 0.8 °C (RCP 4.5) y 2.2 °C (RCP 8.5) más. Los MCG´s predicen al 2050 que la precipitación del 1/4 anual más seco (Bio 17) disminuirá entre 8.7% y 19.1% (RCP´s 4.5 y 8.5) en el área de distribución actual de D. mexicanus. En términos de precipitación 94.7% de los registros se ubican entre 10 mm y 40 mm (Fig. 4c) en Bio 17. En términos de altitud, 92.1% de los registros se ubican entre 1600 m y 2600 m snm (Fig. 4d).

Figura 4 Histograma de frecuencia elaborado con 283 registros de Dendroctonus mexicanus Hopkins, distribuidos sobre tres variables ambientales de mayor contribución Bio 10 (a), Bio 7 (b) y Bio 17 (c), en el periodo actual (azul) y en el promedio de cinco Modelos de Circulación General (líneas) en dos trayectorias de concentración (RCP´s) 4.5 y 8.5 al 2050. Altitud (d).
Distribución potencial de Dendroctonus mexicanus Hopkins
Al realizar la trasferencia del modelo actual a todas las regiones del mundo, se predicen nuevas áreas de idoneidad de D. mexicanus en México (climas análogos acorde a ExDet), e idoneidad, pero climas no análogos en Sudamérica (Bolivia) y en África (Etiopía, Angola y Sudáfrica, mapas no presentados). En ese sentido, la transferencia del modelo al 2050 en México muestra una posible expansión de D. mexicanus (color rojo, Fig. 5a), en total 24 267 km2 y 25 002 km2 estimadas para el RCP 4.5 y 8.5 respectivamente, superficies localizadas principalmente en la parte norte de la SMOc (Durango) y en menor proporción en la SMO (Nuevo León), lo que no se observó para el sur del país (Fig. 5a y 5b); esto implica una posible redistribución geográfica de D. mexicanus, migrando en promedio 30 km ± 15 km hacia latitud norte y longitud este, únicamente en el norte del país.

Figura 5 Áreas de idoneidad actual y futura de Dendroctonus mexicanus Hopkins para el año 2050, promedio estimado de cinco Modelos de Circulación General en dos trayectorias de concentración representativas (RCP), 4.5 (a) y 8.5 (b).
Algunas áreas predichas actualmente como idóneas en el centro del país (color amarillo) podrían no serlo en el 2050; estas representan 47 687 km2 y 51 585 km2 para el RCP 4.5 y 8.5, respectivamente (Fig. 5a y 5b). Al obtener la altitud en un total de 100 000 puntos distribuidos aleatoria-mente en los mapas de idoneidad actual y futura de D. mexicanus, reclasificados al umbral de 0.3639 (mínima presencia), se obtiene un promedio y desviación estándar de 2062 m ± 444.51 m snm (distribución actual); al año 2050, los valores son de 2172 m ± 434.5 m y 2180 m ± 456.1 m snm para los RCP´s 4.5 y 8.5; es decir, acorde con el promedio, esta especie podría migrar hasta 100 m en altitud para el 2050.
Discusión
Perfiles bioclimáticos
El perfil bioclimático de las especies es de suma importancia para conocer sus tolerancias climáticas (Beaumont et al. 2005); aun así, pocos son los estudios que lo presentan. Mendoza et al. (2011) muestran que D. rhizophagus se distribuye entre 10.2 °C y 19.2 °C y desde 305 mm hasta 1406 mm de precipitación (obtenidos con Bio 1 y Bio 12). Con estas mismas variables bioclimáticas, se observa que el perfil de D. mexicanus es más amplio (9 °C - 21 °C), incluso donde la precipitación alcanza 1700 mm. Acorde con Bio 10 (variable que contribuye más al modelo de idoneidad de esta especie), 60% de los registros se presenta a una temperatura de 18 °C, oscilando de 13 °C a 25 °C. Sin embargo, en D. valens (California, E.E.U.U.) la temperatura del mes más cálido (Bio 5) y la precipitación del mes más seco (Bio 14) explicaron su distribución potencial (Maldonado et al., 2015), mientras que la temperatura media (Bio 1) y altitud influyeron significativamente en la modelación de áreas idóneas de D. rhizophagus en el noroeste de México (Mendoza et al., 2011).
En general, se observa que la temperatura es la variable que más influye en la distribución de especies de Dendroctonus spp. (Logan y Powell, 2001; Waring et al., 2009; Bentz et al., 2010; Pureswaran et al., 2018), como se ha evidenciado para esta especie; además, Varela, Lima-Ribeiro y Terribile (2015), indican que los modelos calibrados con variables de temperatura son más robustos comparados con aquellos que solo usan precipitación.
Los histogramas de frecuencias de las Bio 10 y Bio 7 (temperatura), construidos con n = 283, muestran posible ausencia de registros de D. mexicanus en los intervalos que van desde 19 °C a 21 °C (Fig. 4a y 4b), mientras que el histograma de Bio 17 sigue una distribución sesgada a la derecha (Fig. 4c). Beaumont et al. (2005) demostraron que, si las variables poseen una distribución normal o sesgada, ejercen gran influencia en el perfil bioclimático de la especie, aunque construir un histograma debe tener una base estadística, puesto que puede manipularse dependiendo de cómo se determine el número de clases (Sturges, 1926).
Modelos de idoneidad
La distribución potencial de D. mexicanus en el presente estudio es semejante al modelo propuesto para Dendroctonus spp. en México (Sosa et al., 2018), el cual utilizó también Maxent, observándose gran similitud en las áreas de idoneidad predichas para D. mexicanus, demostrando que la mayor idoneidad de este género ocurre donde hay mayor diversidad de especies forestales y condiciones climáticas similares entre hospederos y hospedantes. De igual forma, se aprecian coincidencias con el modelo desarrollado por Salinas et al. (2010), el cual utilizó el algoritmo BIOCLIM, aunque dicha metodología no se recomienda por proporcionar predicciones imprecisas y ajustes bajos (Elith et al., 2006). Salinas et al. (2010) indican que 38% de incidencias de D. mexicanus se presentan en Pinus leiophylla Schl. & Cham., en este sentido, Aceves, Méndez, García y Nájera (2018) muestran que la Bio 10 es variable importante en determinar la distribución potencial de nueve especies de Pinus, incluyendo P. leiophylla, cuyo modelo de idoneidad es similar al de D. mexicanus, distribuyéndose en los mismos intervalos de temperatura desde 11.7 °C hasta 21.1 °C en Bio 10. Al modelar la distribución actual y futura de P. leiophylla, Cruz et al. (2016) revelan que esta es vulnerable al cambio climático, el modelo de idoneidad revela un decremento en su distribución en el sur de México, esto podría indicar que D. mexicanus busque nuevas especies hospederas.
Migración altitudinal
La distribución altitudinal actual de D. mexicanus de esta investigación es la misma que la señalada por Salinas et al. (2010) desde 800 m (Chiapas) hasta 3600 m snm (Estado de México), evidenciándose preferencia de esta especie hacia mayores altitudes (Fig. 4c). Bajo escenarios moderados (RCP 4.5) y drásticos (RCP 8.5) de emisiones de CO2 al 2050, se observa alta idoneidad climática para D. mexicanus, 100 m snm más allá del límite superior altitudinal actual, lo que supone que esta especie podría emigrar hacia esas altitudes. La preferencia de esta especie a migrar hacia mayores altitudes fue observada por Wood (1982) desde hace más de 30 años. Para Pinzón, Cuellar, Jurado y González (2018), la altitud resulta indicadora de la expansión de D. mexicanus, lo cual concuerda con los resultados obtenidos aquí. Régnière (2009) registró nuevos brotes de D. ponderosae en la parte occidental de Canadá, hacia mayores altitudes. Mendoza et al. (2011) muestran que D. rhizophagus tiene preferencias por hábitats más templados y por ende hacia mayores altitudes. En el noreste de E.E.U.U., por cada incremento de 1 °C, los brotes de D. ponderosae han incrementado 30.2 m en elevación (Williams y Liebhold, 2002).
Migración latitudinal
En este estudio se encontró que D. mexicanus podría desplazarse entre 15 km y 45 km hacia el Norte y Este, respecto a su distribución actual; es posible que el aumento de temperatura estimado entre 1.6 °C y 2.0 °C para el 2050, en la temperatura media del 1/4 anual más cálido, sea el responsable de este movimiento. En otras especies de descortezadores, el desplazamiento geográfico de su distribución podría ser más amplio, e. g. Ungerer, Ayres y Lombardero (1999) muestran que, ante un aumento de 3 °C en la temperatura mínima anual, D. frontalis podría extender los límites de distribución de hasta 170 km. Armendáriz et al. (2012) evidenciaron que D. rhizophagus ha ampliado su distribución geográfica 250 km hacia el Sur de la SMOc. En este mismo contexto, Régnière (2009) encontró evidencias de D. ponderosae sobre mayores latitudes, en la parte occidental de Canadá. Moser, Fitzgibbon y Klepzig (2005) documentaron nuevos regis-tros de D. mexicanus al sur de E.E.U.U. (Arizona), esto da evidencia de latitud más alta de esta especie; sin embargo, es importante resaltar que ningún grupo de variables empleadas aquí predijo idoneidad en la zona geográfica indicada por estos autores (Fig. 6a). La idoneidad de D. mexicanus en Sudamérica (Fig. 6b) no presenta clima análogo al área de calibración, según el error tipo 1 de ExDet (Fig. 6c), en especial Bio 3. Áreas idóneas, predichas por el modelo en el resto del mundo, tampoco presentan similitud climática.

Figura 6 Áreas de idoneidad de Dendroctonus mexicanus Hopkins registradas en el mundo, obtenidas a través de la transferencia del modelo a), idoneidad de la especie predicha en Sudamérica b) y evaluación del riesgo de extrapolación de la idoneidad climática de la especie en Sudamérica, basada en distancias de Mahalanobis calculada con ExDet c).
De la misma forma, Logan y Bentz (1999) sugieren que las especies de descortezadores podrían moverse hacia mayores elevaciones y más hacia el norte de su distribución. La revisión exhaustiva de Pureswaran et al. (2018) sobre insectos y cambio climático demuestra que los descortezadores tienen mayor tasa de supervivencia y reproducción al migrar hacia mayores altitudes y latitudes, se expanden hacia nuevas áreas, hospedando incluso nuevas especies, posiblemente porque los MCG´s predicen los mayores incrementos de temperatura en mayores latitudes. Como se puede constatar en la literatura, la respuesta al calentamiento global será diferente entre y dentro de especies de descortezadores (Bentz et al., 2010).
Conclusiones
Las variables que constituyeron el modelo de idoneidad de D. mexicanus fueron: temperatura de los meses más cálidos (Bio 10), oscilación entre la máxima y mínima (Bio 7), dentro de los meses con poca precipitación (Bio 17). Por la alta idoneidad predicha por el modelo, los estados en riesgo de ataque por esta especie son: Estado de México, Michoacán y Puebla. Bajo escenarios moderados (RCP 4.5) y drásticos (RCP 8.5) de emisiones de CO2, al 2050 se espera idoneidad de D. mexicanus en nuevas regiones en el norte del país (24 000 km2 y 25 000 km2), pero pérdida de áreas idóneas de casi el doble en el centro de México (47 000 km2 y 51 000 km2) respectivamente. Probable ampliación de su distribución altitudinal de hasta 100 m y un desplazamiento de entre 15 km y 45 km hacia la latitud Norte y longitud Este. Los resultados hacen suponer que un aumento de temperatura (1.6 °C - 2.0 °C al 2050) en áreas de distribución actual de D. mexicanus podrían tener efectos importantes en la redistribución de esta especie, a pesar de poseer un perfil bioclimático más amplio que otras especies.

