










Services on Demand
Journal
Article
 text in
text in  English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkRevista Chapingo. Serie horticultura
On-line version ISSN 2007-4034Print version ISSN 1027-152X
Rev. Chapingo Ser.Hortic vol.26 n.1 Chapingo Jan./Apr. 2020 Epub June 15, 2020
https://doi.org/10.5154/r.rchsh.2019.06.011
Artículo científico
Dosis y número de aplicaciones de etefón en la reversión sexual de flores de calabacita
1São Paulo State University. Via de acessos Prof. Paulo Donato Castellane s/n, Jaboticabal, São Paulo, 14884-900, BRAZIL.
2Tocantins Federal University. Rua Badejos, lote 7 s/n, Chácaras 69/72, Zona Rural - Jardim Cervilha, Gurupi, Tocantins, 77404-970, BRAZIL.
El etileno puede cambiar el patrón de floración de algunas cucurbitáceas, lo que favorece la aparición de flores femeninas. El objetivo de este estudio fue determinar la influencia de las dosis y el número de aplicaciones de etefón en la reversión sexual de flores de calabacita (Cucurbita pepo L.). El experimento se realizó bajo un arreglo factorial 4 x 4, más un testigo (floración natural); el primer factor fue la dosis de etefón (125, 250, 375 y 500 mg·L-1), y el segundo fue el número de aplicaciones (1, 2, 3 y 4). El etefón influyó en la expresión sexual de la calabacita, pero no lo suficiente como para lograr una reversión sexual completa. La dosis de 500 mg·L-1 de etefón proporcionó la mayor feminización en todas las aplicaciones; por lo que, el etefón podría ser una herramienta útil en los programas de reproducción de calabacita.
Palabras clave: Cucurbita pepo; etileno; fitohormonas; expresión sexual en cucurbitáceas
Ethylene can change the flowering pattern of some cucurbits, favoring the emergence of female flowers. The objective was to determine the influence of doses and number of ethephon applications in sex reversal of zucchini (Cucurbita pepo L.). The experiment was conducted in a 4 x 4 factorial arrangement, plus a control (natural flowering), the first factor being the ethephon doses (125, 250, 375 and 500 mg·L-1) and the second the number of applications (1, 2, 3 and 4). Ethephon influenced the sexual expression of zucchini, but not enough to achieve complete sex reversal. The 500 mg·L-1 dose provided the largest feminization for all applications. Ethephon might be a useful tool for zucchini breeding programs.
Keywords Cucurbita pepo; ethylene; phytohormones; sex expression in Cucurbits
Introducción
La expresión sexual de la calabacita (Cucurbita pepo L.) ‘Caserta’ es monoica, con flores anaranjadas, grandes y vistosas, que están presentes en puntos separados de la misma planta y están aisladas en las axilas de las hojas (Filgueira, 2008). Las flores femeninas, las cuales presentan un ovario alargado, suelen ser menos numerosas que las masculinas (Filgueira, 2008).
La calabacita tiene dos o tres fases sexuales, comenzando con la producción de flores masculinas de manera exclusiva; posteriormente, ocurre la producción de flores masculinas y femeninas de manera alterna, y finalmente se producen flores femeninas (Manzano et al., 2010a; Manzano, Martínez, Megías, Garrido, & Jamilena 2013; Martínez et al., 2014; Peñaranda, Payán, Garrido, Gómez, & Jamilena, 2007).
La genética, el medio ambiente y los factores hormonales influyen en la expresión sexual de las cucurbitáceas (Bhowmick & Jha, 2015; Campbell, Luo, & Mercer, 2013; Manzano, Martínez, García, Megías, & Jamilena, 2014). En la calabacita, las temperaturas bajas inhiben el desarrollo de las flores masculinas y estimulan el desarrollo de las flores femeninas, mientras que las temperaturas altas favorecen la reversión parcial o completa de las flores femeninas a flores bisexuales y estimulan la producción de flores masculinas (Peñaranda et al., 2007).
El etileno es la principal hormona que regula la expresión sexual en las cucurbitáceas (Manzano et al., 2011; Papadopoulou, Little, Hammar, & Grumet, 2005; Yamasaki, Fujii, & Takahashi, 2005). La aplicación exógena de etileno puede modificar la proporción de flores masculinas/femeninas (Manzano et al., 2010a; Payán et al., 2006; Wien, 2006). En el melón (Cucumis melo L.), el pepino (Cucumis sativus L.) y la calabacita, el etileno reduce el desarrollo inicial de las flores masculinas y disminuye la proporción de flores masculinas/femeninas, lo que no sucede en el caso de la sandía (Citrullus lanatus (Thunb.) Matsum & Nakai) (Manzano et al., 2014).
Para aumentar el rendimiento y facilitar los programas de reproducción, algunos estudios sugirieren el uso de reguladores de crecimiento basados en etileno para lograr la reversión sexual en algunas cucurbitáceas. Las yemas florales femeninas requieren una cantidad mínima de etileno para completar su desarrollo y maduración, sin que se produzca un aborto prematuro (Manzano, Martínez, Gómez, Garrido, & Jamilena, 2010b; Manzano et al., 2011; Manzano et al., 2013). El etefón (Ethrel®), cuando se aplica a las plantas, conduce a la liberación inmediata de etileno en los tejidos de las plantas, lo que causa la supresión de flores masculinas, las cuales, después de ser abortadas, son reemplazadas por flores femeninas que se desarrollan en brotes secundarios (Nascimento, Pinheiro, & Freitas, 2007). Esta inducción ginoica, con el uso de etefón, puede ser una tecnología prometedora comparada con otros reguladores, esto debido a sus efectos positivos y facilidad de aplicación (Nascimento, Innecco, Medeiros-Filho, & Oliveira, 2005). No obstante, la intensidad de la feminidad causada por el etefón puede depender de la dosis, el número de aplicaciones y las condiciones ambientales (Friedlander, Atsmon, & Gal, 1997).
La región norte de Brasil es un gran productor de cucurbitáceas, como la sandía y el melón, aunque otras cucurbitáceas, como la calabacita, han ido ganando importancia; sin embargo, los cultivares disponibles no están adaptados a las condiciones de la región, como el estado de Tocantins que tiene un clima cálido. Para producir calabacita de manera más eficiente, se deben desarrollar cultivares adaptados o mejorar las técnicas de manejo. Por ello, el objetivo de este estudio fue determinar la influencia de las dosis y el número de aplicaciones de etefón en la reversión sexual de flores de calabacita.
Materiales y métodos
El experimento se realizó de noviembre de 2013 a enero de 2014 en suelo latosol rojo (Empresa Brasileira de Pesquisa Agropecuaria [Embrapa], 2013), en el Sector de Cultivos Vegetales de la Universidad Federal de Tocantins, Campus Gurupi, Brasil, ubicado a 11° 43’ 45’’ latitud sur y 49° 04’ 07’’ longitud oeste, a 280 msnm. El clima de la región es húmedo con un déficit hídrico moderado, clasificado como Aw (Köppen, 1948).
El experimento se estableció bajo un diseño de bloques completos al azar con tres repeticiones y un arreglo factorial 4 x 4, más un testigo (floración natural). El primer factor fue la dosis de etefón (125, 250, 375 y 500 mg·L-1), utilizando el producto comercial Ethrel®, y el segundo fue el número de aplicaciones (1, 2, 3 y 4).
La primera aplicación de etefón se realizó 10 días después de la emergencia de las plántulas (dde), y el resto cada siete días. El cultivar Caserta se utilizó en siembra directa, para lo cual se colocaron dos semillas por hoyo de plantación y se realizó un aclareo cuando las plantas tuvieron dos hojas verdaderas, dejando una planta por hoyo. Las camas elevadas fueron de 0.7 m de ancho y 0.2 m de alto. La distancia entre plantas fue de 0.5 m bajo una disposición triangular. Cada parcela constó de cinco plantas, y las tres plantas centrales se utilizaron para la recolección de datos.
La fertilización se realizó con 150 g·m-1 de 5N-25P-15K sintético y 20 g·m-1 de micronutrientes (FTE). La capa de abono final se realizó 15 dde con 5 g de 5N-25P-15K formulado por planta. El riego por aspersión y todas las prácticas de cultivo se realizaron de acuerdo con las necesidades del cultivo.
Las dosis de los diferentes tratamientos se prepararon a partir de una solución madre, la cual se diluyó para obtener las diferentes concentraciones. Para todos los tratamientos se prepararon 2 L para su aplicación, y a cada uno se le adicionó 1 % de aceite mineral.
Se realizaron seis conteos de flores masculinas y femeninas cada tres días en seis ocasiones. El primer conteo se realizó 22 dde, y el último a los 36 dde. Las características evaluadas fueron el número de flores masculinas y femeninas, la proporción de flores masculinas/femeninas, el amarre de los frutos y el rendimiento (Mt·ha-1). La tasa de amarre de los frutos se calculó con base en el número total de flores femeninas convertidas en frutos.
Los datos obtenidos se sometieron a análisis de varianza en el programa estadístico SISVAR (Ferreira, 2011). Como el número de flores femeninas y masculinas no cumplió con los supuestos del ANOVA (independencia, homocedasticidad y distribución normal), los valores se expresaron por medio de errores estándar. Para el resto de las variables, se elaboraron modelos de regresión para explicar la variación de los datos observados.
Resultados y discusión
En el tratamiento control (sin aplicación de etefón) hubo aproximadamente el doble de flores masculinas que de flores femeninas, como ocurre generalmente en la floración natural (Figura 1). Por su parte, la aplicación de etefón cambió el patrón de floración, ya que en algunos tratamientos el número de flores masculinas y femeninas fue similar cuando se aplicó la dosis más baja (Figura 1), mientras que con la mayor concentración de etefón (500 mg·L-1) se observaron más flores femeninas en todas las aplicaciones. Los resultados obtenidos coinciden con los reportados por otros autores en calabacita (Iozi, Rodrigues, Goto, & Ono, 1999; Yongan, Bingkui, Enhui, & Zunlian, 2002), en C. moschata (Cardoso, da Silva, & Vecchia, 1998), en calabacita híbrida de Tetsukabuto (Nascimento et al., 2007) y en melón (Girek et al., 2013). Daryono, Prasetya, Sumarlina, Sartika, y Subiastuti (2018) confirmaron que, a partir de 75 ppm, el etefón puede incrementar la producción de flores femeninas y hermafroditas, reducir el número de flores masculinas y aumentar el diámetro de las flores en algunos cultivares de melón.

Figura 1 Número de flores masculinas y femeninas en el cultivo calabacita (Cucurbita pepo L.) ‘Caserta’ bajo diferentes dosis de etefón y número de aplicaciones.
En general, la dosis de etefón afectó la intensidad del cambio de sexo de la flor. A medida que aumentó la concentración de etefón, incrementó el número de flores femeninas y disminuyó el número de flores masculinas. De hecho, una aplicación fue suficiente para cambiar el patrón de floración, y la máxima reversión sexual ocurrió con dos o tres aplicaciones. Manzano et al. (2011) mencionan que una aplicación sería suficiente para influir en la expresión sexual, ya que el etefón reduce la primera fase de desarrollo de las flores masculinas. No obstante, Peixoto, Casali, Almeida, y Seraphin (1989) encontraron que para la calabaza, el grupo C. pepo Baianinha, se requieren al menos dos aplicaciones de 400 mg·L-1 para revertir el sexo de las plantas.
Aunque la dosis de 125 mg·L-1 influyó en la expresión sexual de la calabacita, principalmente con cuatro aplicaciones, el efecto no fue suficiente para inducir la aparición de más flores femeninas que masculinas. Yongan et al. (2002), al aplicar etefón a plantas de calabacita en la etapa de cotiledón, encontraron que 50 mg·L-1 fue la concentración más efectiva para la reversión sexual, y 100 mg·L-1 causó daño o muerte de las plantas, lo cual no fue el caso en este estudio.
A partir de la dosis de 250 mg·L-1 se obtuvieron más flores femeninas que masculinas, excepto por el tratamiento de 250 mg·L-1 con dos aplicaciones y el de 375 mg·L-1 con una aplicación. La dosis de etefón que resultó en el efecto feminizador más alto fue la de 500 mg·L-1, sin importar el número de aplicaciones. Ninguna dosis proporcionó una reversión sexual completa, es decir, plantas con flores femeninas únicamente, lo que concuerda con lo reportado por Shannoon y Robinson (1979), y Cardoso et al. (1998) en calabacita.
Cardoso et al. (1998) también observaron que el etefón no fue capaz de inducir la reversión sexual completa, por lo que propusieron utilizar las dosis y el número de aplicaciones que resultaron en un menor número de flores masculinas en una planta que sería un progenitor femenino en la producción de semillas híbridas F1. Lo anterior ayudaría a reducir la mano de obra necesaria para extraer manualmente las flores masculinas antes de la antesis, lo que facilitaría la reproducción de híbridos de calabacita.
A medida que aumentó la concentración de etefón, se redujo la proporción de flores masculinas/femeninas (Figura 2). La aplicación de etefón promovió una disminución lineal de este rasgo en todos los tratamientos (dosis y número de aplicaciones), lo que se debió a un mayor número de flores femeninas y un menor número de flores masculinas, o al efecto combinado de estos dos elementos.
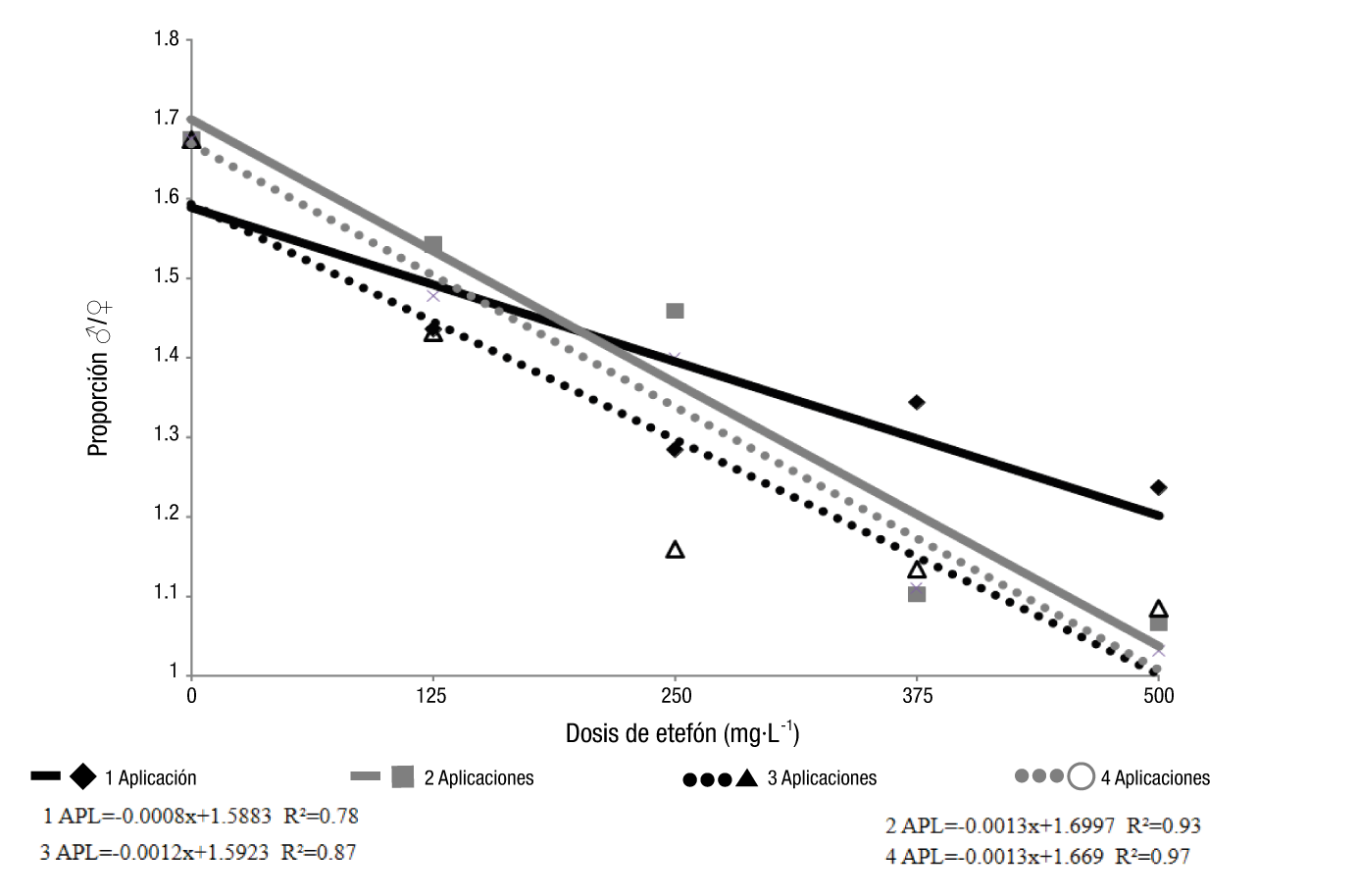
Figura 2 Proporción de flores masculinas/femeninas en el cultivo de calabacita (Cucurbita pepo L.) ‘Caserta’ en función de la dosis de etefón y el número de aplicaciones. Datos transformados a √(x+1).
La dosis de 500 mg·L-1, con tres o cuatro aplicaciones, condujo a una proporción menor de flores masculinas/femeninas. El efecto fue menos marcado cuando el etefón se aplicó una vez, aunque se produjo un comportamiento similar al de otros tratamientos, lo que indica que el etefón afecta la regulación sexual en calabacita.
En condiciones de campo, el porcentaje de amarre de frutos no es total debido a factores ambientales que interfieren con la polinización, la sanidad vegetal y la expresión fenotípica. Al comparar la aplicación de etefón en las dosis de 125 y 250 mg·L-1, se observó que el etefón afectó el amarre del fruto incluso cuando se aplicó la concentración más baja (Figura 3).

Figura 3 Porcentaje de amarre de frutos en calabacita (Cucurbita pepo L.) ‘Caserta’ en función de la dosis de etefón y el número de aplicaciones.
Con el incremento en la dosis de etefón y el número de aplicaciones, a concentraciones más altas, el etefón no ejerce influencia directa en la conversión de flores femeninas en el fruto. Además, debido a que el efecto del etefón estará ligado a su poder feminizador, se reduce la cantidad de polen y la probabilidad de una polinización eficiente, ya que la proporción de flores masculinas/femeninas es más baja de lo normal, así como el número de flores masculinas.
De manera general, la aplicación de etefón afectó el rendimiento de la calabacita de manera adversa. Para una, tres y cuatro aplicaciones se observó un comportamiento cuadrático, y fue lineal con dos aplicaciones (Figura 4). El rendimiento estuvo relacionado con el número de frutos por planta, el cual se vio afectado por el amarre del fruto. La aplicación de etefón es una alternativa de interés agronómico únicamente cuando se realizan cuatro aplicaciones, ya que bajo esta condición se observaron rendimientos más altos; por lo que, si el objetivo es aumentar el número de frutos, se pueden recomendar cuatro aplicaciones.

Figura 4 Rendimiento de calabacita (Cucurbita pepo L.) ‘Caserta’ en función de la dosis de etefón y el número de aplicaciones. Datos transformados a √(x+1).
Con la disminución en la proporción de flores masculinas/femeninas, se esperaría un mayor rendimiento debido a un mayor número de flores femeninas; sin embargo, el rendimiento de los diferentes tratamientos fue menor que el del testigo. Esto puede estar vinculado con la tasa de amarre de los frutos, que a su vez puede estar relacionada con un número de flores masculinas insuficiente para polinizar todas las flores, o con la influencia del etefón en la viabilidad de las flores.
Conclusiones
El uso del etefón influye en la expresión sexual de la calabacita, pero no promueve una reversión sexual completa. La aplicación del etefón puede ser una herramienta útil para la reproducción de calabacita, ya que reduce la necesidad de eliminar las flores masculinas de los progenitores utilizados como líneas madre en cruzas dirigidas o en la producción de semillas.
References
Bhowmick, B. K., & Jha, S. (2015). Dynamics of sex expression and chromosome diversity in Cucurbitaceae: a history in the making. Journal of Genetics, 94(4), 793-808. doi: 10.1007/s12041-015-0562-5 [ Links ]
Campbell, L. G., Luo, J., & Mercer, K. L. (2013). Effect of water availability and genetic diversity on flowering phenology, synchrony and reproductive investment in summer squash (Cucurbita pepo). The Journal of Agricultural Science, 151(6), 775-786. doi: 10.1017/S0021859612000731 [ Links ]
Cardoso, A. I., da Silva, N., & Vecchia, P. T. (1998). Use of ethephon on a summer squash line. Horticultura Brasileira, 16(2), 140-143. doi: 10.1590/S0102-05361998160000200010 [ Links ]
Daryono, B. S., Prasetya, E., Sumarlina, S., Sartika, D., & Subiastuti, A. S. (2018). The effect of ethepon treatment on the formation of flower in melon (Cucumis melo L.). Proceeding of the 2nd International Conference on Tropical Agriculture. doi: 10.29037/digitalpress.21239 [ Links ]
Empresa Brasileira de Pesquisa Agropecuária (Embrapa). (2013). Sistema brasileiro de classificação de solos. Brasília: Author. Retrieved from http://livimagens.sct.embrapa.br/amostras/00053080.pdf [ Links ]
Ferreira, D. F. (2011). Sisvar: a computer statistical analysis system. Ciência e Agrotecnologia, 35(6), 1039-1042. doi: 10.1590/S1413-70542011000600001 [ Links ]
Filgueira, F. A. (2008). Novo Manual de Olericultura. Viçosa, Brazil: Universidade Federal de Viçosa. [ Links ]
Friedlander, M., Atsmon, D., & Galun, E. (1997). The effect of grafting on sex expression in cucumber. Plant and Cell Physiology, 18(6), 1343-1350. doi: 10.1093/oxfordjournals.pcp.a075556 [ Links ]
Girek, Z., Prodavonic, S., Zdravkovic, J., Zivanovic, T., Ugrinovic, M., & Zdravkovic, M. (2013). The effect of growth regulators on sex expression in melon (Cucumis melo L.). Crop Breeding and Applied Biotechnology, 165(13), 165-171. doi: 10.1590/S1984-70332013000300003 [ Links ]
Iozi, R. N., Rodrigues, J. D., Goto, R., & Ono, E. O. (1999). Uso de fitorreguladores na expressão sexual de abobrinha (Cucurbita pepo var. melopepo). Biotemas, 12(2), 47-53. doi: 10.5007/%25x [ Links ]
Köppen, W. (1948). Climatología: con un estudio de los climas de la tierra. México: Fondo de Cultura Económica. [ Links ]
Manzano, S., Martínez, C., Domínguez, V., Avalos, E., Garrido, D., Gómez, P., & Jamilena, M. (2010a). A major gene conferring reduced ethylene sensitivity and maleness in Cucurbita pepo. Journal of Plant Growth Regulation, 29(1), 73-80. doi: 10.1007/s00344-009-9116-5 [ Links ]
Manzano, S., Martínez, C., Gómez, P., Garrido, D., & Jamilena, M. (2010b). Cloning and characterisation of two CTR1-like genes in Cucurbita pepo: regulation of their expression during male and female flower development. Sexual Plant Reproduction, 23(4), 301-313. doi: 10.1007/s00497-010-0140-1 [ Links ]
Manzano, S., Martínez, C., Megías, Z., Gómez, P., Garrido, D., & Jamilena, M. (2011). The role of ethylene and brassinosteroids in the control of sex expression and flower development in Cucurbita pepo. Plant Growth Regulation, 65(2), 213-221. doi: 10.1007/s10725-011-9589-7 [ Links ]
Manzano, S., Martínez, C., Megías, Z., Garrido, D., & Jamilena, M. (2013). Involvement of ethylene biosynthesis and signaling in the transition from male to female flowering in the monoecious Cucurbita pepo. Journal ofPlant Growth Regulation , 32(4), 789-798. doi: 10.1007/s00344-013-9344-6 [ Links ]
Manzano, S., Martínez, C., García, J. M., Megías, Z., & Jamilena, M. (2014). Involvement of ethylene in sex expression and female flower development in watermelon (Citrullus lanatus). Plant Physiology and Biochemistry, 85, 96-104. doi: 10.1016/j.plaphy.2014.11.004 [ Links ]
Martínez, C., Manzano, S., Megías, Z., Garrido, D., Picó, B., & Jamilena, M. (2014). Sources of parthenocarpy for Zucchini breeding: Relationship with ethylene production and sensitivity. Euphytica, 200(3), 349-362. doi: 10.1007/s10681-014-1155-8 [ Links ]
Nascimento, I. B., Innecco, R., Medeiros-Filho, S., & Oliveira, V. H. (2005). Precocidade da colheita e produtividade do meloeiro submetida a soluções de ethephon. Revista Ciencia Agronomica, 36(3), 295-298. Retrieved from http://www.ccarevista.ufc.br/seer/index.php/ccarevista/article/view/240 [ Links ]
Nascimento, W. M., Pinheiro, F., & Freitas, R. A. (2007). Utilização de ethephon para a produção de sementes de híbrido de abóbora tipo tetsukabuto. Revista Brasileira de Sementes, 29(2), 10-14. doi: 10.1590/S0101-31222007000200002 [ Links ]
Papadopoulou, E., Little, H. A., Hammar, S. A., & Grumet, R. (2005). Effect of modified endogenous ethylene production on sex expression, bisexual flower development and fruit production in melon (Cucumis melo L.). Sexual Plant Reproduction, 18(3), 131-142. doi: 10.1007/s00497-005-0006-0 [ Links ]
Payán, M. C., Penaranda, A., Rosales, R., Garrido, D., Gomez, P., Jamilena, M., & Holmes, G. J. (2006). Ethylene mediates the induction of fruits with attached flower in zucchini squash. Proceedings Cucurbitaceae. Retrieved from https://www.researchgate.net/publication/285289123_Ethylene_mediates_the_induction_of_fruits_with_attached_flower_in_zucchini_squash [ Links ]
Peñaranda, A., Payán, M. C., Garrido, D., Gómez, P., & Jamilena, M. (2007). Production of fruits with attached flowers in zucchini squash is correlated with the arrest of maturation of female flowers. The Journal of Horticultural Science and Biotechnology, 82(4), 579-584. doi: 10.1080/14620316.2007.11512276 [ Links ]
Peixoto, N., Casali, V. W., Almeida, A. H., & Seraphin, J. C. (1989). Effect of ethephon on sexual expression of pumpkin. Horticultura Brasileira, 7(1), 20-21. [ Links ]
Shannoon, S., & Robinson, R. W. (1979). The use of ethephon to regulate sex expression of summer squash for hybrid seed production. Journal of the American Society for Horticultural Science, 104(5), 674-677. [ Links ]
Wien, H. C. (2006). Ethephon treatment may alleviate the suppression of female flowers of Cucurbita pepo under high temperatures. American Society for Horticultural Science, 41(6), 1421-1422. doi: 10.21273/HORTSCI.41.6.1421 [ Links ]
Yamasaki, S., Fujii, N., & Takahashi, H. (2005). Hormonal regulation of sex expression in plants. Vitamins & Hormones, 72, 79-110. doi: 10.1016/S0083-6729(05)72003-3 [ Links ]
Yongan, C., Bingkui, Z., Enhui, Z., & Zunlian, Z. (2002). Control of sex expression in summer squash (Cucurbita pepo L.). Cucurbit Genetics Cooperative Report, 25, 51-53. Retrieved from http://cuke.hort.ncsu.edu/CGC/cgc25/cgc25-18.pdf [ Links ]
Recibido: 05 de Junio de 2019; Aprobado: 12 de Septiembre de 2019
 This is an open-access article distributed under the terms of the Creative Commons Attribution License
This is an open-access article distributed under the terms of the Creative Commons Attribution License
