










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkRevista mexicana de ciencias geológicas
On-line version ISSN 2007-2902Print version ISSN 1026-8774
Rev. mex. cienc. geol vol.29 n.1 Ciudad de México Apr. 2012
A new specimen of nyctosaurid pterosaur, cf. Muzquizopteryx sp. from the Late Cretaceous of northeast Mexico
Nuevo espécimen de un pterosaurio nyctosaurido, cf. Muzquizopteryx sp. del Cretácico tardío del noreste de México
Eberhard Frey1, Ross A. Elgin1*, Wolfgang Stinnesbeck2, José Manuel Padilla–Gutiérrez3, Christina Ifrim2, Samuel Giersch1, and Arturo H. González–González4
1 Geowissenschaftliche Abteilung, Staatliches Museum für Naturkunde Karlsruhe, Erbprinzenstraβe 13, 76133 Karlsruhe, Germany.
2 Institut für Geowissenschaften, Universität Heidelberg, Im Neuenheimer Feld 234, D–69120 Heidelberg, Germany.
3 Colección Paleontológica de Coahuila, Museo del Desierto, Prol. Peréz Treviño 3745, Parque Las Maravillas, 25015 Saltillo, Coahuila, Mexico.
4 Museo del Desierto, Prol. Peréz Treviño 3745, Parque Las Maravillas, 25015 Saltillo, Coahuila, Mexico. *dinofrey@aol.com.
Manuscript received: June 9, 2011
Corrected manuscript received: September 9, 2011
Manuscript accepted: September 16, 2011
ABSTRACT
Here we describe a second nyctosaurid pterosaur from the Late Cretaceous laminated limestone deposits of northeast Mexico. The specimen was discovered in the Múzquiz quarry area in northern Coahuila and comprises an isolated right wing skeleton including the humerus, radius/ulna, carpus, wing finger metacarpus and the proximal segment of wing finger phalanx I. The specimen is likely to be a primarily isolated wing, at least the basal wing finger phalanx of which was complete prior to collection. The specimen is referred to cf. Muzquizopteryx sp. on account of its humerus morphology, which is coincident with that of Muzquizopteryx coahuilensis, although a lack of diagnostic characters at the species level prevents any further identification. Its discovery from the Late Turonian deposits of northern Coahuila near Muzquíz, confirms it as the oldest nyctosaurid discovered to date.
Keywords: Muzquizopteryx, Nyctosauridae, Pterosauria, Late Cretaceous, northeast Mexico.
RESUMEN
Se describe un segundo pterosaurio nyctosaurido de los depósitos laminares del Cretácico tardío del Noreste de México. El espécimen fue descubierto en calizas laminares del área de Múzquiz, al norte de Coahuila, México, y consiste de un ala incompleta derecha que incluye: el húmero, radio/ulna , carpo, pteroide, metacarpo del dedo del ala y la porción proximal de la primera falange del dedo del ala o dedo IV. El espécimen es una parte del esqueleto postcraneano, preservado en dos lajas, y al menos la primera falange del dedo del ala estaba completa antes de la colecta. El espécimen es referido a cf. Muzquizopteryx sp. con base en la morfología del húmero, el cual concuerda con la de Muzquizopteryx coahuilensis, conocido del Turoniano Tardío del norte de Coahuila cerca de Muzquiz. Sin embargo, debido a la falta de caracteres diagnósticos a nivel de especie, no es posible llegar a una determinación más exacta. Este hallazgo del Turoniano Tardío proveniente de los depósitos del noreste, cerca de Muzquiz, correspondería al registro más antiguo de Nyctosauridae descubierto hasta la fecha.
Palabras clave: Muzquizopteryx, Nyctosauridae, Pterosauria, Cretácico tardío, noreste de México.
INTRODUCTION
Nyctosauridae comprises two genera, Nyctosaurus (Marsh, 1876) and Muzquizopteryx (Frey et al., 2006). They were edentulous pterodactyloid pterosaurs identified by the hatchet–shaped deltoid crest of the humerus, a straight pteroid with an articular head almost perpendicular to the shaft, a wing finger composed of three instead of four phalanges, the completely reduced three preaxial digits in the wing, and a scapula that does not contact the supraneural plate (Williston, 1903; Bennett, 2003). Further characters have been proposed by Unwin (2003), who placed the nyctosaurs within Pteranodontidae, and Kellner (2003, 2004), who retains Nytosauridae as a clade of its own, following Bennett (1994). Because the controversy on the phylogenetc position of nyctosaurid pterosaurs is unresolved to date, the character states presented in the cladistic analysis of Unwin (2003) and Kellner (2003, 2004) cannot be regarded as diagnostic to date.
Nyctosauridae appear to have been endemic to the North American Western Interior Seaway (WIS) south into the Palaeogulf of Mexico. While one single species, Nyctosaurus lamegoi (Price, 1953) has been reported from the Gramame Formation (Late Cretaceous, Early Maastrichtian) of the Paraiba Basin, northeast Brazil, its identification as Nyctosaurus has not been universally accepted (Kellner, 2003). N. lamegoi would be the largest nyctosaur with a wingspan estimated four metres (Price, 1953).
The majority of nyctosaur specimens come from the Santonian (Late Cretaceous) Niobrara Formation, Kansas, U.S.A., and are all referred to the genus Nyctosaurus (Marsh, 1876). Three different species have been described: N. gracilis (Marsh, 1876), N. bonneri (Miller, 1972a, 1972b), and N. nanus (Marsh, 1881). Bennett (1994) suggests N. bonneri is a junior subjective synonym of N. gracilis, both of which are from the Upper Chalk of the Smoky Hills. N. nanus comes from the Lower Chalk of the same region and was identified as Nyctosaurus by Schoch (1984). Bennett (2003) described two nyctosaurid specimens from the Lower Chalk of Trego County, which both have a huge branching crest on top of the skull. These specimens were referred to N. gracilis and the crest was supposed to grow during late ontogeny. However, the specimens have a wing–span of only two metres and could therefore also represent adult N. nanus (Everhart, 2009). Before the discovery of the crested specimens, nyctosaurids were thought to have occipital crests similar to that of a Pteranodon longiceps or P. ingens (e.g., Wellnhofer, 1991).
In 2006 a small, nearly complete nyctosaurid pterosaur was described as Muzquizopteryx coahuilensis (UNAM IMG 8621; Frey et al., 2006). The adult individual had a wingspan of two metres and a bluntly rounded and straight caudally directed occipital crest. Coming from the Lower Coniacian (middle Late Cretaceous) of the El Rosario Quarry, approximately 170 km WNW of the town of Múzquiz (northern Coahuila, Figure 1) Muzquizopteryx represented the oldest known nyctosaur prior to the discovery of the new specimen (Figure 2).
Here we report on a second specimen of Muzquizopteryx comprising a partial wing skeleton. It is about 20 % smaller than the first specimen and was obtained by a prívate collector from a local quarry worker. Muzquizopteryx coahuilenis is the only valid pterosaur taxon from the Cretaceous of Mexico.
MATERIAL AND METHODS
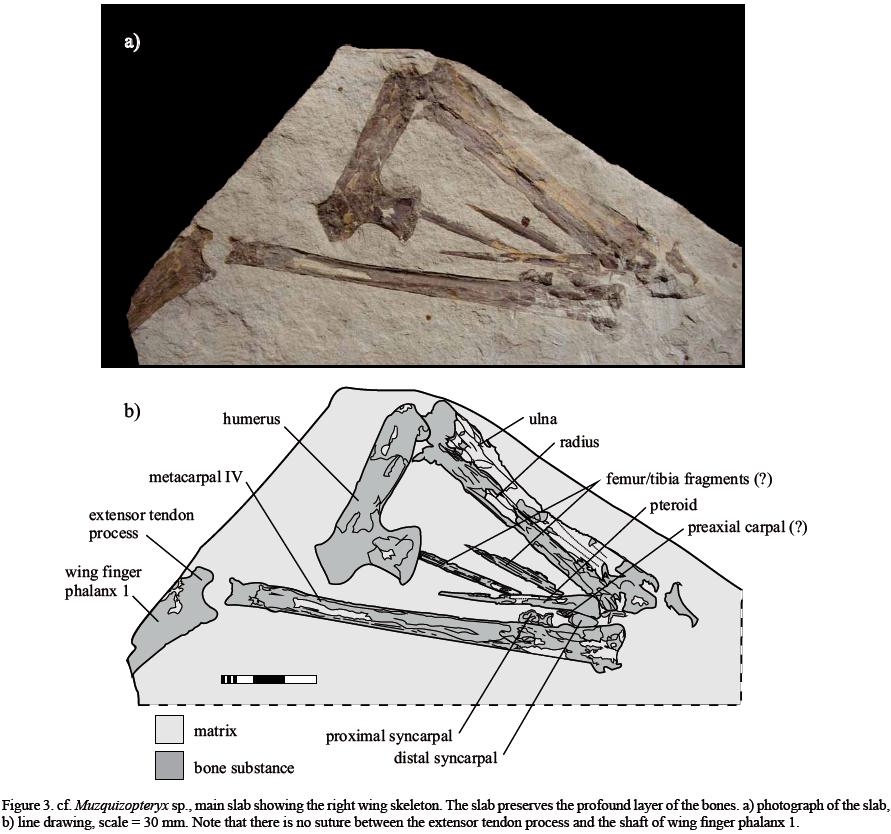
The specimen is housed in the collection of the Museo del Desierto Saltillo (MUDE), Coahuila (collection number CPC–484). The original plate was split into two parts, but because the original orientation cannot be reconstructed, i.e. , which of the two slabs was the bottom one, the plate with the better preservation is here defined as the main slab (Figure 3) and the other one as the counter slab (Figure 4). Besides the initial splitting of the slab, no preparation has been undertaken. Photographs were taken with a Canon D 400 SLR camera and traced with Corel Graphics Suite 12. The traced image of the counter slab was projected below that of the main slab in order to provide an image of the entire specimen (Figure 5).
GEOLOGY AND LITHOLOGY
The Múzquiz area in northern Coahuila, Mexico, is known for its evenly layered laminated limestone and fissile marly limestone that, owing to their platy fabric, are intensively quarried in the region for building faces, flagstones and wall tiles. These deposits are of Cenomanian to Early Coniacian age (Late Cretaceous) and contain vertebrate fossils with exceptionally well–preserved anatomical details of their soft tissues, as well as a diverse number of invertebrates (Stinnesbeck et al., 2005). Sediment deposition was in an open marine shelf environment near the southern opening of the WIS, several hundreds of kilometres south of the North American coast line, at a depth of at least 50–100 m (Stinnesbeck et al., 2005).
The new specimen is embedded within a yellow tan slab and counterslab, each about 20 mm thick, of micritic, evenly layered, laminated limestone with internal millimetric bedding which is best described by the German word Plattenkalk. According to the lithology and microfossil contents of the host matrix, the specimen most likely comes from the El Rosario or La Carranza quarry field about 150 km north of Múzquiz, northern Coahuila (Figure 1). The matrix of the new specimen coincides with the lower part of the Yellow Member of the El Rosario section (Stinnesbeck et al., 2005; Frey et al., 2006; Figure 2). A micropaleontological analysis of the host matrix of cf. Muzquizopteryx revealed the following planktonic foraminifers: Archaeoglobigerina cretacea, Dicarinella hagni, D. imbricata, D. primitiva, Heterohelix reussi, Marginotruncana schneegansi, M. sigali, Whiteinella cf. baltica, and W. brittonensis. D. primitiva and M. sigali are diagnostic taxa of a Late Turonian age. This, combined with the absence of H. helvetica and D. concavata dates the new specimen of cf. Muzquizopteryx to a Late Turonian age (Premoli Silva and Sliter 1999) making it the oldest known nyctosaurid pterosaur (Figure 2).
Preservation (Figures 3, 4, 5)
Due to the manner in which the plate was split, the humerus, the proximal two thirds of radius and ulna, the distal two thirds of the wing finger metacarpal and the proximal fragment of the wing finger are distributed in the centre of the slab (Figures 3, 4). Most of the carpus and the adjacent thirds of the radius/ulna and wing finger metacarpal are preserved exclusively on the main slab having left only external moulds on the counter slab (Figure 3). Pieces of compacta are only preserved on the counter slab (Figure 4). Overlapping projections of the main and counter slab indicates that a few chips of bone were lost in the distal third of the ulna and the proximal fifth of the wing finger metacarpal during splitting and possibly subsequent refitting attempts (Figure 5).
The slabs preserve the proximal portion of a right wing skeleton including the humerus, radius and ulna, carpus and the pteroid, wing finger metacarpal (mc IV) and the proximal fragment of the first wing finger phalanx which terminates at the border of the slab and was likely complete prior to collection (Figures 3, 4). All elements lie almost in natural articulation but are bent beyond bone lock. While the elbow joint is in articulation, the carpus has disarticulated with the proximal and distal syncarpals lying along the ventral margin of the wing finger metacarpal. The proximal syncarpal therefore now lies distal to the other carpals (Figures 3, 5) while the distal syncarpal and the preaxial carpal are wedged between the articular end of the pteroid and the wing finger metacarpal (Figures 3, 5). The pteroid parallels the wing finger metacarpal and has rotated along its long axis so that its articular condyle is now facing cranially. The wing finger metacarpal has rotated 90° around its long axis and is now seen in cranial view lying slightly separated from the fragmented basal wing finger phalanx. Isolated fragments, likely the ipsilateral hind limb, lie parallel to each other between the pteroid bone and radius/ulna at an angle of about 10° to the latter elements (Figures 3, 4, 5). All bones show traces of impaction with typical longitudinal break lines. Due to a late diagenetic compaction, the preserved diameter of the bones is regarded as representing their original dimensions. The humerus lies at an angle of about 75° to radius and ulna, which in turn lie parallel to each other. The metacarpal IV defines an angle of about 40° with radius and ulna and 135° with the first wing finger phalanx (Figure 5).
SYSTEMATIC PALAEONTOLOGY AND DESCRIPTION
Pterosauria Kaup, 1834
Pterodactyloidea Plieninger, 1901
Nyctosauridae Marsh, 1876
cf. Muzquizopteryx Frey et al., 2006
cf. Muzquizopteryx sp.
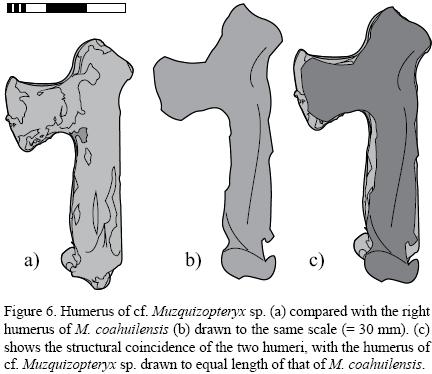
The specimen comes from a pterosaur that was 81 % the size of the holotype of M. coahuilensis (Figure 6). The humerus is hatchet–shaped with a deltopectoral crest displaying concave proximal and distal margins while the pteroid is straight with a semi–circular articular head, which is off set at almost 90° to the pteroid shaft. Both these features are diagnostic for Nyctosauridae Marsh, 1876 (Figures 3, 4, 5). The shape of the humerus coincides with that of M. coahuilensis to such detail (Figure 6) that both specimens are likely conspecific, however, the concavity of the caudomedial margin of the pteroid is slightly shallower than in that of M. coahuilensis. All other bones are too badly preserved for a direct comparison, but do not show any significant morphological differences when compared to their respective elements in M. coahuilensis. Furthermore, the ratios between the humerus and radius/ulna as well as the humerus and pteroid show only minor differences in the two specimens and these may be solely due to their preservation (Table 1). In contrast to the holotype of M. coahuilensis there are no mineralized tendons in the new specimen. As the described specimen does not preserve any diagnostic features to refer it to M. coahuilensis, but is otherwise morphologically coincident, we refer the new specimen to cf. Muzquizopteryx sp.

Description
The description of the material is based on the overlapping projection of the slab and the counter slab (Figure 5). The anatomical directions refer to the inferred flight position of the animal with the wing bones fully laterally extended. The caudal margin of the humeral shaft is straight while the cranial edge is slightly convex, with the deltopectoral crest emerging from the proximal third of this margin of the humeral shaft (Figures 3, 4, 5, 6a). The distal margin of the deltopectoral crest is straight but offset from the cranial margin of the shaft by an angle of 55° (Figure 6). The proximal margin of the deltopectoral crest stands at a right angle to the cranial margin of the shaft but forms an angle of about 70° with the caudally inclined neck of the humerus. The cranial margin of the humeral neck is slightly convex. The humeral neck measures one fourth of the length of the shaft and is angled at about 130° against this (Figure 6a). The proximal articular face of the humerus is straight and orientated at 90° to the humeral neck. The distal articulation of the humerus is damaged but shows a fragmentary distally facing articular condyles, which likely protruded cranially by a maximum of one fifth of the distal diameter of the shaft (Figures 3, 4, 5, 6a).
The ulna overlies the cranial margin of the radius although both bones are damaged such that no anatomical features are visible. The absolute length of both bones can only be estimated due to their damaged distal articular ends (Figures 3, 4, 5). Due to the partial coverage of the distal syncarpal and the preaxial carpal by the pteroid and the wing finger metacarpal, nothing can be said about their features. The proximal syncarpal is visible in cranial view. The distal margin lies free and shows the convex articulation face for the cranioventral part of the proximal articulation facet of distal syncarpal and the dorsolateral process that articulates with the dorsal face of the distal syncarpal (Figures 3, 4, 5).
Metacarpal IV is exposed in cranial view and shows an S–shaped articular facet for the distal syncarpal, whereby the ventral margin is medially convex and the dorsal margin is slightly concave. The dorsal margin of the shaft is straight while the ventral margin is slightly concave converging distally towards the distal articular end, which is half as wide as the proximal articular surface (Figures 3, 4, 5).
The pteroid shows the typical elongate triangular shape diagnostic of nyctosaurids (Figures 3, 5). Its cranial margin is straight while the caudal one is slightly concave. The concavitiy of the caudal margin gradually increases proximally. The distal margin of the bone is straight, forming an angle of 104° with the cranial margin, and the transition between the two margins is evenly convex. The articular surface is a tiny semicircular protrusion at the caudodistal corner of the pteroid, which forms one sixth of the lateral margin of the bone.
Of the first wing finger phalanx only the proximal segment is preserved. The extensor tendon process is internally fused with the shaft of the phalanx (Figure 3) but superficially shows a suture (Figure 4). Otherwise there are no remarkable features on the bone fragment (Figures 3, 4, 5).
Two unidentified bone fragments lying parallel to the radius are too thick to represent parts of the distal wing finger phalanges and thus likely come from the ipsilateral hind limb. However, the elements are fragmentary and preserve no osteological features (Figures 3, 4, 5).
Only one feature allows a statement on the age of the cf. Muzquizopteryx specimen: near complete fusion of the extensor tendon process with the shaft of the first wing finger phalanx (Figures 3, 4). Because the described specimen is only 81 % the size of the holotype of M. coahuilensis, a late fusion of the extensor tendon process with the basal wing finger phalanx occurred during the ontogeny Muzquizopteryx (Frey and Martill 1998; Bennett 1993; Elgin and Frey 2008). Therefore, the new specimen is osteologically immature and thus likely subadult individual at the time of death.
Taphonomy
The absence of any shoulder girdle remains, the grade of disarticulation and lack of mineralised tendons indicate that the specimen likely represents a primarily isolated fragment (Figures 3, 4, 5). The wing presumably separated from a pterosaur carcass in an advanced stage of decay where the main flight musculature was no longer viable. The cardinal ligaments of the wing were, however, still able to hold the bones in a near to natural position. From the status of decay and a comparison with the fully articulated holotype of M. coahuilensis, conclusions on the ligament stability are possible at least for this individual. Here the elbow joint is the most stable (Figures 3, 5). The humerus and the radius/ulna remain in tight articulation with an angle of 75° between the bones. This is less than the articulation angle of the same bones in M. coahuilensis and may represent maximum humeroantebrachial flexion. The carpus has been completely disrupted (Figures 3, 5). The position of the syncarpals, the rotation of pteroid and wing finger metacarpal all indicate an advanced state of decay of the carpal ligaments. However, the separation of the carpals suggests that either at least some of the ligaments were isolated from each others and thus could react independently during advancing decay, or that the carpus was damaged by the bite of a predator or scavenger prior to deposition. The separation of the first wing finger phalanx from its articulation with metacarpal IV is explainable by muscular decay where only the massive extensor tendon was still viable. Following this, it is likely that the remains of the dermis and the wing membrane, connecting the fore arm to the body and hind limbs, would have prevented the complete decay of the wing finger. Examples of this style of preservation are common throughout the pterosaur fossil record (Frey et al., 2003; Martill and Frey, 1998; Unwin and Martill, 2007). If the wing membrane was still viable, the wing skeleton would occasionally detach from the body along with the ipsilateral hind limb. It is for this reason that the indeterminate long bone fragments are interpreted here as belonging to the left hind limb.
CONCLUSIONS
The new nyctosaurid specimen, comprised of an isolated fragmentary wing, likely originates from the Turonian limestone quarries of El Rosario or La Carranza 150 km north of Múzquiz, northern Coahuila (Figures 1, 2). While several bones are preserved close to their natural articulations, the carcass is interpreted as having floated on the surface waters for an extended period, allowing for a relatively advanced state of decay. Due to the presence of potential scavengers, however, the scenario of prolonged surface decay remains more unlikely than dismembering by either a scavenger or a predator. Due to the lack of diagnostic characters, the specimen can only be determined on a comparative level, using bone biometry and morphological coincidence. Despite this, it is sufficiently similar to the holotype of M. coahuilensis to warrant a referral to cf. Muzquizopteryx sp. making it the geologically oldest nyctosaurid discovered pterosaur to date. The specimen also provides evidence of the potential of the Múzquiz area to yield more nyctosaurid pterosaurs in the future.
ACKNOWLEDGMENTS
Our special thanks go to Veronica Ruiz Santos, Múzquiz, for allowing us to describe the specimen and granting access to it through the Museo del Desierto, where the specimen is registered and housed. For critical, helpful and constructive reviews of the manuscript we cordially thank Laura Codorniu (Universidad Nacional de San Luis, Argentina) and David M. Martill (Portsmouth, United Kingdom). We also wish to thank the chief editors of the Revista Mexicana de Ciencias Geológicas for their assistance with the manuscript. The publication is a result of the DFG projects FR 1314/15, STI 128/9, and we are grateful to the DFG for funding these projects.
REFERENCES
Bennett, S.C., 1993, The ontogeny of Pteranodon and other pterosaurs: Paleobiology, 19, 92–106. [ Links ]
Bennett, S.C., 1994, Taxonomy and systematics of the Late Cretaceous pterosaur Pteranodon (Pterosauria, Pterodactyloidea): Occasional papers of the Natural History Museum, The University of Kansas, 169, 1–70. [ Links ]
Bennett, S.C., 2003, New crested specimens of the Late Cretaceous pterosaur Nyctosaurus: Stuttgart, Paläontologische Zeitschrift, 77(1), 61–75. [ Links ]
Elgin, A.R., Frey, E. 2008, You show me your sutures and I'll tell you your age. Age traces in a young Anhanguera skeleton: 6th Meeting of the European Association of Vertebrate Palaeontologists, 30th June – 5th July 2008, Volume of Abstracts, Spisská Nová Ves: Spisská Nová Ves, Slovak Republic, Múzeum Spisa, 33–34. [ Links ]
Everhart, M.J. 2009, Pteranodons, flying reptiles of the Late Cretaceous Western Interior Sea – a photographic atlas; internet publication: http://www.oceansofkansas.com/Pteranodon.html. [ Links ]
Frey, E., Martill, D.M., 1998, Late ontogentic fusion of the processus tendinis extensoris in Cretaceous pterosaurs from Brazil: Neues Jahrbuch für Geologie und Paläontologie Monatshefte, 10, 587549, Stuttgart. [ Links ]
Frey, E., Tischlinger, H., Martill, D.M., Buchy, M.–C., 2003, New specimens of Pterosauria (Reptilia) with soft parts, with implications for pterosaurian anatomy and locomotion, in Buffetaut, E., Mazin, J.–M. (eds.), Evolution and palaeobiology ofpterosaurs: Proceedings of the Geological Society of London, Special Publication, 217, 233–266, London. [ Links ]
Frey, E., Buchy, M.–C., Stinnesbeck, W., González, A.G., di Stefano, A., 2006, Muzquizopteryx coahuilensis n.g., n. sp., a nyctosaurid pterosaur with soft tissue preservation from the Coniacian (Late Cretaceous) of northeast Mexico (Coahuila): Oryctos, 6, 19–39. [ Links ]
Kaup, J., 1834, Versuch einer Einteilung der Sãugetiere: Jena, Isis von Oken, 315 pp. [ Links ]
Kellner, A.W.A., 2003, Pterosaur phylogeny and comments on the evolutionary history of the group, in Buffetaut, E., Mazin, J.–M. (eds.), Evolution and Palaeobiology of Pterosaurs: London, Proceedings of the Geological Society of London, Special Publications, 217, 105–137. [ Links ]
Kellner, A.W.A., 2004, New information on the Tapejaridae (Pterosauria, Pterodactyloidea) and discussion of the relationships of this clade: Ameghiniana, 41, 521–534. [ Links ]
Marsh, O.C., 1876, Notice on a new sub–order of Pterosauria: American Journal of Science, 11 (3), 507–509. [ Links ]
Marsh, O.C., 1881, Note on American pterodactyls: American Journal of Science, 21 (3), 342–343. [ Links ]
Martill, D.M., Frey, E., 1998, Soft tissues in pterosaurs: Leicester Transactions of the Leicester Literary and Philosophical Society, 92, 25–26. [ Links ]
Miller, H.W., 1972a, A skull of Pteranodon (Longicepia) longiceps Marsh associated with wing and body bones: Transactions of the Kansas Academy of Science, 74, 20–23. [ Links ]
Miller, H.W., 1972b, The taxonomy of the Pteranodon species from Kansas: Transactions of the Kansas Academy of Science, 74, 1–19. [ Links ]
Plieninger, F., 1901, Beiträge zur Kenntnis der Flugsaurier: Paläontographica, 48, 65–90, plates 4–5. [ Links ]
Premoli Silva, I, Sliter, W.V., 1999, Cretaceous paleoceanography: evidence from planktontic foraminiferal evolution, in Berrera, E., Johnson, C.C. (eds.), Evolution of the Cretaceous ocean–climate system: Boulder, Geological Society of America, Special Paper 332, 301–328. [ Links ]
Price, L.I., 1953, A presença de pterosáuria no Cretáceo Superior do Estado da Paraíba: Rio de Janeiro, Divisão de Geologia e Mineralogía, Notas preliminares e estudos, 71, 5–11. [ Links ]
Schoch, R. M., 1984, Notes on the type specimens of Pteranodon and Nyctosaurus (Pterosauria, Pteranodontidae) in the Yale Peabody Museum Collections: Postilla, 194, 1–23. [ Links ]
Stinnesbeck, W., Ifrim, C., Schmidt, H., Rindfleisch, A., Buchy, M.–C., Frey, E., González González, A.H., Porras–Múzquiz, H., Cavin, L., Smith, K.T., 2005, A new lithographic limestone deposit in the Upper Cretaceous (Coniacian) Austin Group at El Rosario, county of Muzquíz, state of Coahuila, northeastern Mexico: Revista Méxicana de Ciencias Geológicas, 22(3), 401–418. [ Links ]
Wellnhofer, P., 1991, The illustrated encyclopedia of pterosaurs: New York, Crescent Books, 192 pp. [ Links ]
Williston, S.W., 1903, On the osteology of Nyctosaurus (Nyctodactylus), with notes on American pterosaurs: Chicago, Field Columbian Museum, Publication 78, Geological Series 2(3), 125–163. [ Links ]
Unwin, D.M., 2003. On the phylogeny and evolutionary history of pterosaurs, in Mazin, J.M., Buffetaut, E. (eds.), Evolution and Palaeobiology of Pterosaurs: London, Geological Society of London, Special Publication, pp. 139–190. [ Links ]
Unwin, D.M., Martill, D.M., 2007, Pterosaurs of the Crato Formation, in Martill, D. M., Bechly, G., Loveridge, R. F. (eds.), The Crato Fossil Beds of Brazil: Window into an Ancient World: Cambridge ,Cambridge University Press, 475–524. [ Links ]


{kind=link}