










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkRevista mexicana de ciencias geológicas
On-line version ISSN 2007-2902Print version ISSN 1026-8774
Rev. mex. cienc. geol vol.28 n.3 Ciudad de México Dec. 2011
Facies analysis of the Asmari Formation in central and north–central Zagros basin, southwest Iran: Biostratigraphy, paleoecology and diagenesis
Análisis de facies de la Formación Asmari en la región central y nor–central de la cuenca de Zagros, suroeste de Irán: Bioestratigrafía, paleoecología y diagénesis
Ali Seyrafian1* Hossein Vaziri–Moghaddam1, Naser Arzani2, and Azizolah Taheri3
1 Department of Geology, Faculty of Sciences, University of Isfahan, Isfahan, Iran.*seyrafian_ali@hotmail.com
2 Geology Department, University of Payame Noor, Isfahan, Iran.
3 Geology Department, Faculty of Earth Science, Shahrood University of Technology, Shahrood, Iran.
Manuscript received: November 29, 2010
Corrected manuscript received: May 31, 2011
Manuscript accepted: June 6, 2011
ABSTRACT
Three sections of the Oligocene–Miocene Asmari Formation, crossing central and north–central Zagros foreland basin in SW Iran, were measured and studied in order to interpret the biostratigraphy, paleoecology (based on distribution of benthic foraminifera) and diagenesis. Forty–three foraminifer genera and species were encountered in the studied areas and the following assemblage zones have been defined: 1) Nummulites vascus–Nummulites fichteli, 2) Lepidocyclina–Operculina–Ditrupa, 3) Archaias asmaricus–Archaias hensoni–Miogypsinoides complanatus, 4) Miogypsina–Elphidium sp. 14 – Peneroplis farsenensis, and 5) Borelis melo curdica–Borelis melo melo. According to this study, deposition of the Asmari Formation with association of hyaline, lamellar, perforate large and flat foraminifera first started in the basin and slope environments during the Rupelian in Dehdez and Tufe–Sefid areas. Lagoon depositional environment colonized by sea–grass epiphytic foraminifera was encountered during Chattian and Aquitanian in Bagh–e Malek and Dehdez areas and mostly lagoon and slope environments prevailed during Burdigalian in Bagh–e Malek and Dehdez areas, respectively. The main diagenetic processes that affected the Asmari Formation were dolomitization (replacement and cementation), compaction (stylolitization) and dissolution. The extent of these diagenetic overprinting seems to be mainly facies controlled.
Keywords: biostratigraphy, paleoecology, diagenesis, Asmari Formation, Zagros basin, Iran.
RESUMEN
Tres secciones de la Formación Asmari del Oligoceno–Mioceno, que cruza la region central y nor–central de la Cuenca de Zagros en el SE de Irán, fueron medidas y estudiadas, a fin de interpretar su bioestratigrafa, paleoecología (con base en la distribución de foraminíferos bentónicos) y diagénesis. Cuarenta y tres géneros y species de foraminíferos fueron encontrados en las áreas de estudio, y las siguientes zonas de composición fueron definidas: 1) Nummulites vascus–Nummulitesfichteli, 2) Lepidocyclina–Operculina–Ditrupa, 3) Archaias asmaricus–Archaias hensoni–Miogypsinoides complanatus, 4) Miogypsina–Elphidium sp. 14 – Peneroplis farsenensis y 5) Borelis melo curdica–Borelis melo melo. De acuerdo con este estudio, el depósito de la Formación Asmari, en asociación con grandes foraminiíferos aplanados, 4ialinos, lamelares y perforados, comenzó inicialmente en ambientes de cuenca y talud continental durante el Rupeliano en las áreas de Dehdez and y Tufe–Sefid. Un ambiente de depósito de laguna, colonizada porforaminíferos epifíticos de pastos marinos, fue encontrado durante el C4attiano y Aquitaniano en las áreas de Bagh–e Malek y Dehdez, y principalmente ambientes lagunares y de talud continental prevalecieron durante el Burdigaliano en las áreas de Bagh–e Malek y Dehdez, respectivamente. Los principales procesos diagenéticos que afectaron a la Formación Asmari fueron dolomitización (reemplazamiento y cementación, compactación (stilolitización) y disolución. La extensión de estos efectos diagenéticos, parece 4aber sido controlada principalmente por los tipos de facies.
Palabras clave: biostratigrafía, paleoecología, diagénesis, Formación Asmari, cuenca de Zagros, Irán.
INTRODUCTION
The Oligocene–Miocene Asmari Formation is well–known as a major hydrocarbon reservoir in southwestern Iran. Early studies of the Asmari Formation are due to Busk and Mayo (1918), Richardson (1924), Lees and Richardson (1940) and Thomas (1948). Later, the Asmari Formation studied by Wynd (1965), James and Wynd (1965), Adams and Bourgeois (1967), Jalali (1987), Kalantari (1986) and Motiei (1993) to review and improve the previous works and define the Asmari Formation throughout the Zagros basin. Recent works concerning the biostratigraphy of the Asmari Formation are Seyrafian et al. (1996), Seyrafian and Mojikhalifeh (2005), Hakimzadeh and Seyrafian (2008), Amirshahkarami (2008) and Sadeghi et al. (2009). Researches concerning the sedimentary and facies analysis and depositional environment of the Asmari Formation are Seyrafian and Hamedani (2003), Vaziri–Moghaddam et al. (2006), Amirshahkarami et al. (2007a, 2007b), Fakhari et al. (2008), and Mossadegh et al. (2009). Diagenesis of the Asmari Formation has been recently received more attention (e.g., Aqrawi et al, 2006; Ehrenberg et al, 2006; Al–Aasm et al, 2009). Also, recent works concerning to Zagros structural style and basin analy–sis are Sherkati et al. (2006), Ahmadhadi et al. (2008), and Heydari (2008).
The purposes of this study are (1) to present comprehensive biostratigraphic criteria based on newly strontium isotope stratigraphy and biozones (Laursen et al., 2009), and strontium isotope data obtained for three outcrops of the Asmari Formation examined through this research with respect to the International Stratigraphic Chart (ICS, 2010) and to revise the time interval framework for the used earlier, mostly based on the Adams and Bourgeois (1967) biostratigraphic classification; (2) to discuss the environmental controls on the distribution of Oligocene–Miocene benthic foraminifera and relate them to the depositional setting in the central and north–central parts of the Zagros basin; and (3) to investigate the main diagenetic processes and their extent, revealed in these outcrops, in order to gain insight on controlling factors in reservoir quality in the Asmari Formation.
STUDY AREA AND METHODS
This study is based on three sections of the Asmari Formation crossing the central and north–central Zagros region in the Izeh (sections 1and 2) and High Zagros (section 3) zones (Figure 1). Field work focused on detailed description of bed by bed biotic component, facies and diagenetic characteristics. 799 samples from three outcrops of the Asmari Formation with a total thickness of 855.5 m were collected for microscopic studies, to identify the distribution of foraminifera and biostratigraphical characteristics of each section. The diagenetic evolution has been studied on the basis of field observation and petrography of stained thin sections (alizarine red–S and K ferrisyanid; Dickson, 1965). Cathodoluminescence (CL) microscopy was also carried out for selected samples. The extent of dolomitization and porosity of each thin–section was visually estimated and presented as percentage based on standard charts (Flügel, 2004). Dolomite fabrics were described according to Sibley and Gregg (1987).
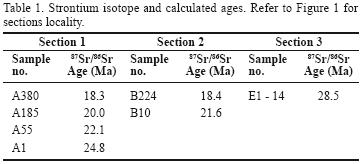
On the basis of identified biostratigraphic time intervals, limited samples were selected for strontium isotope dating. Strontium isotope sample preparation and analyses were done at CSIRO Division of Petroleum Research of Australia (Table 1).
REGIONAL SETTING
The Zagros region is located to the southwest of Iran. It is divided into six major tectonostratigraphic domains: 1) and 2) the interior and coastal Fars provinces, 3) Dezful embayment, 4) the Izeh zone, 5) the Lurestan province, and 6) the High Zagros zone (Figure 1). The Zagros basin was associated to the Gonwdwana supercontinent during the Paleozoic. It was a site of passive margin and convergent orogeny in the Mesozoic and Cenozoic eras, respectively (Motiei, 1993; Bahroudi and Koyi, 2004; Heydari, 2008).
Section 1 located in the Izeh zone, 12 km east–north–east of Bagh–e Malek town. Fieldwork concentrated along the Abol Abass valley at 31°31.76' N, 49°59.66' E (Figures 1 and 2). The Asmari Formation at the Bagh–e Malek is 384 m thick. At the base, it comprises medium– to thick–bedded grey limestone with bioclast remains and nodular structure. In the middle part, the thick beds of limestone are predominant. Bioturbated features, bioclasts remains (ostrea and coral debris) and nodular structures are common. To the top, the formation remains thick bedded and bioclasts debris (gastropod), bioturbation and stylolites are present.
Section 2 is also located in the Izeh zone, 16 km northwest of Dehdez town. The Asmari outcrops studied next to the Kolmat Shalloo village at 31°48.85'N, 50° 4.32' E (Figures 1 and 2). The Asmari Formation at the Dehdez is 427.5 m thick. At the base, it is predominantly thick to massive, light grey to cream limestone with bioclast debris including Lepidocyclina. The middle part of the Asmari Formation at section 2 is characterized by thick to massive bedded limestone. Nodular structure, bioclast debris (ostrea), coral fragments and horizontal burrows are present. Upward, inter–beds of thin, medium to thick limestone are exposed. Horizontal burrows, ripple mark features, and upward thickening of bed intervals are common.
Section 3 located in the High Zagros zone, at 32°35.86' N, 50°9.88' E, 3.5 km southwest of Tufe–Sefid town (Figures 1 and 2). It is 44 m thick and comprises thick to massive beds of light grey limestone. Bioclast debris (pecten, bryozoa, Lepidocyclina and coralline red algae) and stylolite features are present.
BIOSTRATIGRAPHY
Biostratigraphic criteria of the Asmari Formation were studied by Wynd (1965) and the following biozones were introduced: Globigerina spp. (zone 55), Lepidocyclina— Operculina–Ditrupa (zone 56), Nummulites intermedius—Nummulites vascus (zone 57), and Archaias operculiniformis (zone 58) assemblage zones for the Oligocene; Austrotrillina howchini—Peneroplis evolutus (zone 59) assemblage zones for Aquitanian and Borelis melo curdica assemblage zone for the Burdigalian times.
Adams and Bourgeois (1967) reviewed the previous biostratigraphic studies and suggested the following biozones for the Asmari Formation: Eulepidina–Nephrolepidina—Nummulites assemblage zone for the Oligocene; Miogypsinoides–Archaias–Valvulinid assemblage zone for the Aquitanian; Archaias Asmaricus–Archaias hensoni and Elphidium sp. 14–Miogypsina assemblage sub zones for the Early to Middle and Middle to Late Aquitanian ages respectively and Borelis melo group–Meandropsina iraniaca assemblage zone for the Burdigalian. Adams and Bourgeois (1967) did not separate Chattian from Rupelian. The biozones introduced by Wynd (1965) and Adams and Bourgeois (1967) were widely used throughout the Zagros and Central Iranian basins for the Asmari Formation and its age equivalent Qom formations, respectively (Daneshian and Ramezani Dana, 2007).
Cahuzac and Poignant (1997) defined the following assemblage zones: Nummulites vascus—Nummulites fichteli and Eulepidina formosoides assemblage zones for the Rupelian; Nummulites vascus–Nummulites fichteli and Eulepidina and Miogypsinoides–Eulepidina assemblage zones for the Early and Late Chattian respectively; Austotrillina howchini–Miogypsina–Miogypsinoides deharti for the Aquitanian and Borelis melo group–Miogypsina for the Burdigalian age. These biozones are mostly applicable for the European basin (Figure 3).
Ehrenberg et al. (2007) conducted strontium isotope dating to improve the biostratigraphic, depositional sequences and duration of sequences and paraseqeunces ages for the Asmari Formation. He introduced five biostratigraphic events based on index fossils, species of Nummulites and Spiroclypeus blankenhorni, genus of Miogypsina and Archaias and species of Borelis melo curdica. Based on this study, last Nummulites occurrences is about 1 Ma before the end of Rupelian time. Extinction of Nummulites near–end of the Rupelian was also stated by Racey (1995). This leads to recognize the Rupelian–Chattian boundary and to fulfill the biostratigraphic classification proposed earlier by Adams and Bourgeois (1967) for the Asmari Formation. The Burdigalian age suggested for the species Borelis melo curdica is unanimously supported by all researchers.
Van Buchem et al. (2010) applied strontium isotope dating for the Asmari Formation and proposed revised time intervals based on new biozones: Nummulites vascus–Nummulites fichteli assemblage zone for the Rupelian, Lepidocyclina–Operculina–Ditrupa assemblage zone for the Rupelian into Chattian, Archaias Asmaricus–Archaias hensoni–Miogypsinoides complanatus assemblage zone for the Aquitanian and Borelis melo curdica–Borelis melo melo assemblage zone for the Burdigalian age (Figure 4).
A total of 43 foraminifer genera and species were encountered in the studied areas and their distributions have been plotted (Figures 5, 6, 7). Assemblages associated to the Asmari Formation are correlated (Figure 8). Some selected foraminifera of the studied sections are presented (Figures 9 and 10). Five assemblages were identified and are discussed in ascending stratigraphic order as follows:
Assemblage 1 is present in section 2 (Dehdez) and section 3 (Tuf–e Sefid) areas. The most important and common foraminifera in both sections are: Eulepidina dilitata, Eulepidina elephantina, Nephrolepidina tournoueri, Lepidocyclina sp., Nummulites vascus, Nummulites sp., Operculina sp., Operculina complanata, Spiroclypeus sp., Amphistegina sp., Neorotalia viennoti, Heterostegina sp. and miliolids. This assemblage corresponds to the Nummulites vascus–Nummulitesfichteli assemblage zone of van Buchem et al. (2010) and is considered to be Rupelian in age (Figures 6 and 7).
Assemblage 2 occurs only at section 1 (Bagh–e Malek area). The most important fauna are: Nephrolepidina sp., Operculina sp., Operculina complanata, Heterostegina praecursor, Neorotalia sp. and Miogypsinoides sp. This assemblage can be correlated with the Lepidocyclina–Operculina–Ditrupa assemblage zone of van Buchem et al. (2010)).
Assemblage 3 is recorded only in section 2 (Dehdez area). This assemblage consists of Austrotrillina sp., Heterostegina costata, Heterostegina sp., Archaias sp., Peneroplis thomasi, Peneroplis sp., Neorotalia viennoti, Miogypsinoides complanatus, Miogypsinoides sp., Borelis haueri, Borelis pygmaea, Amphistegina sp. and miliolids. It represents the Archaias asmaricus–Archaias hensoni–Miogypsinoides complanatus assemblage zone of van Buchem et al. (2010) and indicates a Chattian age (Figure 6).
Assemblage 4 is present in section 1 (Bagh–e Malek) and section 2 (Dehdez) areas. The most important and common foraminifera in both sections are: Peneroplis sp., Penoroplis farsensis, Borelis sp., Miogypsina sp., Miogypsina irregularis, Miogypsina basraensis, Neorotalia viennoti, Miogypsinoides sp., Elphidium sp. and miliolids. These foraminifera are correlated with Miogypsina–Elphidium sp. 14 – Peneroplis farsenensis assemblage zone of van Buchem et al. (2010) and are attributed to the Aquitanian (Figures 5 and 6).
Assemblage 5 occurs in section 1 (Bagh–e Malek) and section 2 (Dehdez) areas. Common foraminifera in both sections are: Borelis melo curdica, Borelis sp., Borelispygmaea, Dendritina rangi, Meandropsina iranica, Meandropsina anahensis, Elphidium sp., Amphistegina sp., Asterigerina sp., Austrotrillina sp. and miliolids. These foraminifera correspond to Borelis melo curdica–Borelis melo melo assemblage zone of van Buchem et al. (2010) and indícate Burdigalian in age (Figures 5 and 6).
DISCUSSION
Eight samples (four from section 1, three from section 2 and one from section 1) were selected for isotope dating. Results are listed in Table 1 and presented in bold font in Figures 5, 6, 7. Selection was made with the aim to understand (1) the time equivalent of the lowest boundary of the Asmari Formation and time equivalent of the last occurrence of species Borelis melo curdica; (2) the age interval of the Asmari Formation across Izeh and High Zagros zones; and (3) the applicability of new biozonation established for the Asmari Formation throughout the Dezful embayment by van Buchem et al. (2010) in Izeh and High Zagros zones (central and north–central Zagros basin) with respect to the International Stratigraphic Chart (ICS, 2010).
Samples E1–14 and A1 were taken from the lowest part of the Asmari Formation in sections 3 and 1 (Tuf–e Sefid in High Zagros zone, northern part of the Izeh zone), respectively. Isotope dating reveals that deposition of the Asmari Formation started at 28.5 Ma in north and north–central High Zagros (Section 3). At the same time, the southern part of the Izeh zone next to Dezful embayment (Section 1) was still a site of a pelagic sedimentation of the Pabdeh Formation. This area did not prevailed by the carbonate deposits of the Asmari Formation until 24.8 Ma. In refer to ICS (2010), the Chattian time interval ranges from 28.4 to 23.03 Ma; therefore, assemblage 2 associated to the lower Asmari Formation in section 1 (Bagh–e Malek area) is Chattian in age. This confirms the younger Asmari base boundary and lack of assemblage 1 (Nummulites vascus–Nummulitesfichteli assemblage zone of van Buchem et al. (2010) in section one (southern Izeh zone next to Dezful embayment) (Figure 8).
Last occurrence of the species Borelis melo curdica is due to 18.3 and 18.4 Ma, associated to section 1 and 2 respectively. This reflects a similar time equivalent for the upper boundary of the Asmari Formation in the Izeh zone and supports the Burdigalian age suggested for the species Borelis melo curdica. Based on these data, with the exception of Rupelian time, we conclude that the Izeh zone was site of almost similar biostratigraphic characteristics from Aquitanian to Burdigalian. However, during Chattian the Izeh zone northward experienced the Lepidocyclina–Operculina–Ditrupa assemblage zone of van Buchem et al. (2010) and Archaias asmaricus–Archaias hensoni–Miogypsinoides complanatus assemblage zone of van Buchem et al. (2010) (from section 1 to section 2, respectively; Figure 8).
Isotope dating for this study supports the considered ages. Ages are almost coincident with biostratigraphic strontium isotope dating proposed for the Asmari Formation throughout the Dezful embayment by van Buchem et al. (2010) and International Stratigraphic Chart (ICS, 2010).
PALAEOECOLOGY
During the Oligocene–Miocene a nearly 300–500 m–thick sedimentary succession of mostly platform carbonates was deposited as the Asmari Formation in the Zagros basin, located at the southwest Iran (Motiei, 1993). The carbonate succession accumulated within a relatively stable, near equatorial setting, and limited terrigenous input was intermittently laid down within the carbonate intervals throughout this period. Thin section analysis of samples from the Asmari Formation indicate that much of the carbonate sediments are composed of larger benthic foraminifera and red algae, with subordinate amounts of small benthonic foraminifera, bryozoans, echinoderms, mollusks (gastropods, pelecypods), serpulid worm tubes, and non–skeletal grains (peloids, intraclasts and ooids).
The palaeoenvironmental distribution of foraminiferal assemblages and depositional conditions have been reconstructed, combining comparison with the depth range of recent foraminifera, analysis of the functional morphology of larger foraminifera and comparison with Oligocene–Miocene microfossils distribution of the Zagros basin (Vaziri–Moghaddam et al., 2006; Amirshahkarami et al., 2007a, 2007b). This biotic association is typical of shallow, benthic calcareous communities in modern tropical and subtropical marine carbonate environments. The presence of larger foraminifera in the carbonate rocks of the Asmari Formation show the persistence of an equatorial climate throughout deposition of the formation. As in other Tethyan carbonate platforms (Buxton and Pedley, 1989), nummulitids, amphisteginids and lepidocyclinids were especially important during the Paleogene in the Zagros basin, not only as rock–formers but also as biostratigraphic markers.
The distribution of larger foraminifera appears to have largely depended on light, salinity, temperature, substrate and water turbulence (Murray, 1991). Hence, larger foraminifera are excellent indicators used as valuable tools to reconstruct palaeoenvironmental models in warm, shallow marine environments, especially in monotonous carbonate platform successions (Geel, 2000).
It is known, for example, that symbiont–bearing larger perforate foraminifera (e.g., nummulitids and lepidocyclinids) require normal oceanic salinities, whereas imperforate forms tolerate higher salinities (Hallock and Glenn, 1986; Geel, 2000). Fossil of nummulitids, alveolinids and lepidocyclinids are supposed to bear photo–symbiotic micro–algae, which explains their maximum abundances in oligotrophic, tropical shallow marine environments. They are extreme K–strategists, characterized by slow growth, late maturity and low reproduction potential. In nutrient–rich waters this advantage disappears, and small, r–mode foraminifers flourish (Hallock, 1988).
As stated by Hallock and Glenn (1986), perforate living foraminifera exhibit change trends in test morphology that may be related to light and hydrodynamic forces in the environment, with flatter tests and thinner test walls being typical of low–energy and low–light environments and very thick, lamellate test walls being typical of higher energy environments.
The occurrence of a large number of porcelaneous imperforate foraminiferal tests may point to the depositional environment being slightly hypersaline. The biotic assemblages indicate deposition within the photic zone, in a sea–grass–dominated environment, as suggested by the presence of epiphytic porcellaneous foraminifera (Borelis, Archaias, Peneroplis). Such an assemblage has been interpreted as a shelf–lagoon environment (Wilson 1975; Flügel 1982, 2004; Vaziri–Moghaddam et al, 2006).
Generally, the upper photic zone is dominated by porcelaneous larger foraminifera (such as Peneroplis and Archaias), hosting dinoflagellate, rhodophycean and chlorophycean endosymbionts (Leutenegger 1984; Romero et al, 2002; Lee, 1990; Holzman et al., 2001).
Environmental factors and skeletal grain associations
Temperature and latitude appear to control skeletal grain associations that occur on continental shelves. A number of classifications can be recognized in modern marine carbonate environments on the basis of the biotic assemblages. Lees and Buller (1972) and Lees (1975) classified the distinctive skeletal grain assemblages along temperature gradients as chlorozoan, chloralgal and foramol. Carannante et al. (1988) carried out a detailed study of facies zonations on the modern Brazilian shelf, showing variations in carbonate facies associations both with latitude and depth. They introduced the terms rodalgal and molechfor. Recently, to deduce sediment composition from temperature, James (1997) has suggested the terms photozoan and heterozoan associations to classify carbonate sediments on light dependency.
The two distinctive biotic associations show a clear distribution in the modern ocean, both with respect to latitude and temperature. Photozoan associations are subject to dominate tropical and subtropical ocean basins, whereas heterozoan assemblages tend to occur on the cooler eastern sides of the oceans, impacted in part by upwelled nutrient–rich waters. Lees and Buller (1972), Lees (1975) and Carannante et al. (1988) concluded that parameters such as depth, nutrients and salinity additionally influence skeletal grains. Consequently, a certain skeletal grain may exist in various environments.
The compositional analysis of the Asmari Formation shows that the main carbonate producing biota is represented by red algal, small benthonic foraminifera (rotaliids and miliolids) and the large benthic foraminifera. These skeletal components belong to the rhodalgal association that dominated the Oligocene–Miocene carbonate systems in the Zagros region. The biotic assemblage (this study) and palaeolatitudinal reconstructions (Alavi, 2007) from the Asmari Formation clearly suggest a tropical depositional setting.
In the light of modern analogues, the occurrence of larger benthic foraminifera and zooxanthellate corals in the geologic record is interpreted as photozoan assemblages. Nevertheless, the Asmari Formation is dominated by heterozoan assemblages. The abundance of larger benthic foraminifera (Borelis, Archaias, Peneroplis, nummulitides, lepidocyclinides) and zooxanthellate corals on many platforms, however, argues against persistently high nutrient contents, since these foraminifera thrive in oligotrophic (Langer and Hottinger, 2000) to possibly slightly mesotrophic (Halfar et al, 2004) waters.
Heterozoan carbonates are also well developed in the Late Oligocene of Malta (Brandano et al, 2009). The microfacies of the Malta are quite similar to those described in the study area. In the carbonate platform of the Attard Member and Asmari Formation, zooaxanthellate corals did not form a reef framework. According to Pomar and Hallock (2007), until the Late Miocene, zooaxanthellate corals did not build framework structures in the Mediterranean and lived in the middle–lower part of the photic zone. Therefore, the spread of heterozoan assemblages in the Asmari Formation was related to the palaeoecology of zooaxanthellate corals. The interpretation used here may be confirmed by the interpretation of the Late Oligocene carbonate platform of Malta (Brandano et al., 2009). Here, the authors interpreted that the spread of heterozoan assemblages was related to low capacity of corals to thrive in high–light conditions and to form a wave–resistant reef, which promoted the diffusion of heterozoan assemblages.
Foraminiferal assemblage and depositional model
In the current paper, identification and functional morphological analysis of the larger foraminifera within the biogenic grain assemblages used to relate these organisms to their likely depositional environments within the Zagros basin during the Oligocene–Miocene.
The faunal data, obtained from thin sections, allowed the detailed biostratigraphy and characterization of a foraminiferal assemblage, containing both larger benthic imperforate foraminifera and hyaline foraminifera, to be applied to the interpretation of paleoenvironmental conditions. The absence of frame–building organisms capable of forming wave–resistant reefs leading to a steep–margined platform, the carbonates of Asmari Formation in this area, represents sedimentation on a carbonate ramp (see Flügel, 2004).
Three major depositional environments were identified in the Oligocene–Miocene succession in the study sections, on the basis of the distribution of the foraminifera and vertical facies relationships. These include inner ramp, middle ramp and outer ramp.
Inner ramp biotic assemblages represent a wider spectrum of marginal marine deposits indicating open lagoon and protected lagoon. The biotic assemblage of the restricted lagoon environment suggests deposition in the euphotic zone in a depositional environment colonized by seagrass as suggested by the presence of epiphytic foraminifera such as: Archaias, Peneroplis and Borelis (Brandano et al., 2009). Further indications of restricted lagoonal conditions include the presence of miliolids (Figure 11a–11d).
The predominance of mud–rich lithologies with oligotypic fauna (such as miliolids) and the presence of a low–diversity foraminiferal association indicate a very shallow subtidal environment with low to moderate energy. Although miliolids may be found in a variety of very shallow, hyposaline to hypersaline environments, or even in abundance on fore–reef slopes, they are generally taken as evidence for restricted lagoonal (Murray, 1991) and/or relatively nutrient–rich back reef environments (Geel, 2000).
Precise paleobathymetry based on the model developed by Hallock and Glenn (1986), show that the abundance of lenticular perforate specimens of larger rotaliids such as Amphistegina and Neorotalia with imperforate foraminifera suggests that sedimentation took place under shallower, more agitated inner ramp conditions. The biotic association of the inner ramp in this area originated from tropical and subtropical shallow water (Lee, 1990; Betzler et al., 1997; Holzmann et al., 2001; Brandano et al, 2009).
The middle ramp biotic assemblage is presented by nummulitids, lepidocyclinids and Amphistegina. The abundance of deeper water larger foraminifera (such as Operculina and Eulepidina) and Neorotalia specimens suggest sedimentation in the oligophotic zone of the middle ramp (Figure 11e–11g). Generally, the lower part of the upper photic zone is dominated by perforate hyaline foraminifera as lepidocyclinids, Nummulites, Operculina, and Hetrerostegina (Bassi et al., 2007; Brandano et al., 2009). Operculina complanata are found in the deepest photic zone on sandy substrates (Hohenegger, 2000; Hohenegger et al, 2000; Beavington–Penney and Racey, 2004). This species seems to tolerate a high amount of terrigenous sediment that reduces the light intensity (Bassi et al., 2007).
The outer ramp environment is characterized by abundant hyaline, lamellar, perforate large and flat foraminifera such as nummulitids and lepidocyclinids (Figure 11h). These assemblages often dominate in the lower part of photic zone (Leutenegger, 1984; Romero et al, 2002). The simultaneous occurrence of the large perforated foraminiferal tests such as large and flat symbiont bearing nummulitids, i.e., Operculina and Heterostegina with planktonic foraminifera represents the deepest environments of the lower limit of the photic zone (Geel, 2000; Romero et al., 2002).
DIAGENESIS
Diagenetic history and evolution of the studied Asmari Formation was affected by a series of processes including micritization and development of micritic envelopes, physical compaction, cementation, pressure dissolution, dolomitization, dissolution, late dolomitization and tectonic fracturing (Figure 12). Dolomoitization and dissolution, which are discussed below, were the main diagenetic processes that affected and changed the original texture, especially overprinting the porosity of this formation. However, their extent markedly varied in Izeh (Sections 1and 2) and High Zagros (Section 3) zones ( Figures 5, 6, 7).
Dolomitization
Dolomitization, as cement and replacement, is the most important diagenetic event and an important control on porosity and permeability development in the Asmari Formation. However, the degree of dolomitization is variable from minor (<10%) in the section 3 and basal parts of section 1 and 2 to pervasive (>50%) in middle to upper parts of sections 2 and 3 (Figures 5, 6, 7). This spectrum is mostly lithofacies controlled, with the lowest values in outer ramp, distal slope marls and wackestones and also biostromal coral–algal floatstones–rudstones, whereas the highest values are in inner ramp facies.
The dolomite cementation started relatively early in shallow burial and occurred after shell breakage, filling part of the primary porosities (Figure 13a–13c). However, fabric–preserving and very early dolomites (dolomicrites) are very rare in the studied area. In general, the main phase of dolomitization as dolomite cement and replacement started after chemical compaction and are concentrated along dissolution seams and stylolites. Five types of dolomite microfabrics were observed in the study areas.
Type 1: Planar–e microdolomite. They are the early replacive and the main components in the dolomicrite (mudstone) beds (Figure 13d). The dolomite crystals are <10µm in size, non–ferroan and brown to orange color in CL. These crystals have an iron–rich thin rim in some beds and they form a net fabric in partially dolomitized wackestones. The complete to partial dolomitization of the micritic matrix of the mudstones to wackestones resulted in dolomicrite microfacies. The incomplete replacement of the allochems and micritic matrix resulted in a pseudo–pellets (ghosts or calcitic blebs) floating in dolomudstone microfacies.
Type 2: Planar–e medium to coarse crystalline dolomite. They comprise the scatted dolomite crystals floating in the micritic matrix or completely or partially replacing the allochems and are polymodal in size (10–150µm, Figure 13e). They are euhedral, isolated dolomite rhombs which are thoroughly bright in CL and/or isolated dolomite rhoms with cloudy, inclusion–rich cores and limpid outer rims. The latter shows zoning with bright and dull zones. The calcitic core of these crystals is reddish in stained thin sections. The euhedral dolomite rhombs concentrated along the stylolites.
Type 3: Planar–s medium to coarse crystalline dolomite. They have a polymodal size distribution (10µ–>150µ), and the pervasive dolomitization, in this case, resulted in sucrosic and chalky textures with soft beds in the field (Figure 13b).
Type 4: Zoned, planar–e dolomite to anhedral, non–planar crystals (10–>200µm) replacing allochems (e.g., corals and bryozoan fragments). They grade into pore–filling dolomite cement (Figure 13g–13k).
Type 5: Drusy and saddle–like dolomite cement. They are fractured and pore–filling cement with large dolomite crystals, which are zoned and grade from mildly–ferroan (relatively dull luminescence) to ferroan (dull luminescence) dolomites (Figure 13l–13n).
Dissolution and porosity evolution
Other than dissolution seams and stylolites, which are not discussed in details here, dissolution as leaching is a major control on the evolution of porosity and is a very important diagenetic process in the studied sections. The porosity percentages, like degree of dolomitization, are variable from minor (<2.5%) in the section 3 and basal parts of section 1 and 2 to common (>7.5%) in middle to upper parts of the sections 2 and 3 ( Figures 5, 6, 7). This spectrum is lithofacies controlled, with the lowest values in outer ramp, distal slope marls and wackestones and also biostromal coral–algal floatstones–rudstones, and the highest values in inner ramp facies. The primary porosity is very rare, but the porous intervals exhibit mainly secondary porosity as solution enlarged, mouldic, vuggy and intercrystalline types that cross cut the allochems and partially dololmitized micritic matrix (calcitic blebs), is a major type of porosity (Figure 13h–13k; 13n–13r). The latter occurred after early dolomitization (dolomite cementation and replacement). These porosities were filled in the next stage by mildly–ferroan dolomite and anhydrite cements. The pore–filling mildly–ferroan cements show slightly wavy extinction in stained thin sections and are dull, zoned in CL. Late dissolution of midly–ferroan dolomite cement is common. The dissolution of anhydrite cement in these exposures is inferred from the presence rare pore–filling anhydrite cement.
Discussion
Dolomitisation and dissolution are the most important processes in diagenetic overprinting of the porosity in different microfacies of the studied sections of the Asmari Formation. The widespread development of dolomite in the Asmari Formation has been already reported (e.g., James and Wynd, 1965) and its origin has been investigated (Ehrenberg et al, 2007; Aqrawi et al., 2006; Al–Aasm et al., 2009). Dolomitization seems to be have been started early at shallow burial and after physical compaction and shell breakage and continued after stylolitization into deep burial. The zoned nature of the dolomite crystals indicates that the process of dolomitization was not a single event, but occurred in several stages through the burial of sediments. Aqrawi et al. (2006) suggested that the micritic matrix was the first component to be replaced by dolomite and that replacement took place syn–depositionally or during early stage of the diagenesis. This is not a general rule in the studies sections. However, dolomitization occurred through several phases as is evident by different zoned, replaced or cement crystals. The degree of dolomitization was not uniform through the different stratigraphic parts of these sections and diagenetic heterogeneity is extreme when the Izeh (Sections 1and 2) and High Zagros (Section 3) zones are compared. However, it is generally facies controlled as the extent of dolomitization is lowest is in outer ramp and the highest in inner ramp lithofacies.
The association of dolomites with anhydrite cement may suggest the circulation of brines, possibly associated with the evaporites of the Gachsaran Formation through fault conducts and along thrust belts in High Zagros. The evaporitic signature of the dolomites in the Asmari Formation has been also established by strontium isotope and other geochemical data (see Ehrenberg et al., 2007; Aqrawi et al, 2006; Al–Aasm et al, 2009). The general trend of degree of dolomitization in different studied sections and their relationship to porous intervals, suggest that there is a relationship between dolomitization and porosity. However, there are exceptions in some sections; the dolomite beds (dolomudstones) are not porous. The dominant porosity in most sections is intercrystalline and mouldic; the latter representing leaching of calcitic blebs and resulted during deeper burial, as discussed above (Figure 11). These porosities overprinted to vuggy pore types with non–uniform distribution in the studied sections.
The relationship between dolomitization and porosity enhancement has received much attention (e.g., Choquette et al., 1992; Purser et al, 1994; Ehrenberg et. al, 2006; Lucia, 2007). There are many factors controlling the porosity in Asmari Formation, among which dolomitization is very important. However, there are at least three important factors that determine if dolomitization has a positive or negative effect on porosity in the Asmari Formation. These are: degree of dolomitization as cement and replacement, dolomitization along stylolites, and post–dolomite dissolution. The porous nature of some of the mudstone with dolomicritic textures is different from what has been documented for other porous micritic carbonate reservoirs of the Middle East (e.g., Budd, 1989). The presence of calcitic blebs and relics of the micritic allochems indicate that postdolomite dissolution may have been very important in the Asmari Formation. Dissolution as leaching is a major control on the evolution of porosity and is a very important diagenetic process in the studied sections.
CONCLUSIONS
The outcrops of the Asmari Formation at Bagh–e Malek, Dehdez and Tufe–Sefid areas located in central and north–central Zagros are Chattian–Burdigalian, Rupelian–Burdigalian and Rupelian in age, respectively. In other words, while deposition of the Asmari Formation continued during Rupelian (Dehdez and Tufe–Sefid areas), the Bagh–e Malek area was still site of basinal deposition of the Pabdeh Formation. Lack of Chattian to Burdigalian deposits of the Asmari Formation in Tufe–Sefid area may be due to erosion or non–deposition.
During the Rupelian, the large benthic perforated and planktonic foraminifera associated to deeper and lower limit of the photic zone environment of outer ramp settings prevailed in Dehdez and Tufe–Sefid areas. During Chattian and Aquitanian the large benthic hyaline perforated and imperforated foraminifera associated to an oligophotic zone environment of middle ramp settings (restricted and semi–restricted lagoon) are characteristics of Bagh–e Malek and Dehdez areas. While Bagh–e Malek area was mostly site of euphotic restricted lagoon inner ramp deposition, Dehdez area was strongly site of outer ramp (slope) and sporadically bar sedimentation in Burdigalian.
Diagenetic history and evolution of the studied Asmari Formation waas affected by a series of processes including micritization, physical compaction, cementation, pressure dissolution, dolomitization, dissolution, late dolomitization and tectonic fracturing. Dolomitization and dissolution are the most important processes in diagenetic overprinting of the porosity in different microfacies of the studied sections of the Asmari Formation.
ACKNOWLEDGEMENTS
The authors wish to thank the National Iranian Oil Company Research and Development Management for their financial support and permission to publish this work. The authors appreciate the National Iranian Oil Company Exploration Division for providing surface geological maps.
We thank the Revista Mexicana de Ciencias Geológicas reviewer Dr. Juan Carlos García y Barragán and anonymus reviewers for their constructive comments. We appreciate Mrs. Nafarieh, Soltanian and Saleh for preparing the figures, plates and photography.
REFERENCES
Adams, T.D., Bourgeois F., 1967, Asmari biostratigraphy: Iranian Oil Operating Companies, Geological and Exploration Division, Report 1074. [ Links ]
Ahmadhadi, F., Daniel J.M., Azzizadeh, M., Lacombe, O., 2008, Evidence for pre–folding vein development in the Oligo–Miocene Asmari Formation in the Central Zagros Fold Belt, Iran: Tectonics, 27, 1–22. [ Links ]
Al–Aasm, I.S., Ghazban F., Ranjbaran, M., 2009, Dolomitization and related fluid evolution in the Oligocene–Miocene Asmari Formation, Gachsaran Area, SW Iran; petrographic and isotopic evidence: Journal of Petroleum Geology, 32(3), 287–304. [ Links ]
Alavi, M., 2007, Structures of the Zagros fold–thrust belt in Iran: American Journal of Sciences, 307, 1064–1095. [ Links ]
Amirshahkarami, M., 2008, Distribution of miogypsinoides in the Zagros Basin, in southwest Iran: Historical Biology, 20,175–184. [ Links ]
Amirshahkarami, M., Vaziri–Moghaddam, H., Taheri, A., 2007a, Paleoenvironmental model and sequence stratigraphy of the Asmari Formation in southwest Iran: Historical Biology, 19(2), 173–183. [ Links ]
Amirshahkarami, M., Vaziri–Moghaddam, H., Taheri, A., 2007b, Sedimentary facies and sequence stratigraphy of the Asmari Formation at Chaman Bolbol, Zagros Basin Iran: Journal of Asian Earth Sciences, 29, 947–959. [ Links ]
Aqrawi, A.A.M., Keramati, M., Ehrenberg, S.N., Pickard, A., Moallemi, A., Svana, T., Darke, G., Dickson, J.A.D., Oxtoby, N.H., 2006, The origin of dolomite in the Asmari Formation (Oligocene–Lower Miocene), Dezful embayment, SW Iran: Journal of Petroleum Geology, 29 (4), 381–402. [ Links ]
Bahroudi, A., Koyi, H.A., 2004, Tectono–sedimentary framework of the Gachsaran Formation in the Zagros foreland basin: Marine and Petroleum Geology, 21(10), 1295–1310. [ Links ]
Bassi, D., Hottinger, L., Nebelsick, J.H., 2007, Larger Foraminifera from the Upper Oligocene of the Venetian area, northeast Italy: Palaeontology, 50(4), 845–868. [ Links ]
Beavington–Penney, S.J., Racey, A., 2004, Ecology of extant nummulitids and other larger benthic foraminifera: applications in palaeoenvironmental analysis: Earth Science Review, 67(3–4), 219–265. [ Links ]
Betzler, C., Brachert, T., Nebelsick, J.H., 1997, The warm temperate carbonate province: facies, zonations and delimitations: Courier Forschungsinstitut Senckenberg, 201, 83–100. [ Links ]
Brandano, M., Frezza, V., Tomassetti, L., Cuffaro, M., 2009, Heterozoan carbonates in oligotrophic tropical waters: The Attard member of the Lower Coralline Limestone Formation (Upper Oligocene), Malta: Palaeogeography, Palaeoclimatology, Palaeoecology 274, 54–63. [ Links ]
Budd, D.A., 1989 Micro–rhombic calcite and microporosity in limestones: A geochemical study of the Lower Cretaceous Thamama Group U.A.E: Sedimentary Geology, 63, 293–311. [ Links ]
Busk, H.G., Mayo, H.T., 1918, Some notes on the geology of the Persian oilfields: Journal Institute Petroleum Technology, 5, 5–26. [ Links ]
Buxton M.W.N., Pedley, H.M., 1989, A standardized model for Thethyan Tertiary carbonate ramps: Journal of the Geological Society, London, 146(5), 746–748. [ Links ]
Cahuzac, B., Poignant, A., 1997, An attempt of biozonation of the European basin, by means of larger neritic foraminifera: Bulletin Society Geology. Farance, 168, 155–169. [ Links ]
Carannante, G., Esteban, M., Milliman, J.D., Simone, L., 1988, Carbonate lithofacies as paleolatitude indicators: problems and limitations: Sedimentary Geology, 60, 333–346. [ Links ]
Choquette, P.W., Cox, A., Meyer, W.J., 1992, Characteristics, distribution and origin of porosity in shelf dolostones, Burlington–Keokuk Formation (Mississippian), U.S. mid–continent: Journal of Sedimentary Petrology, 62, 167–189. [ Links ]
Daneshian, J., Ramezani Dana, L., 2007, Early Miocene benthic foraminifera and Biostratigraphy of the Qom Formation, Deh Namak, Central Iran: Jurnal of Asian Earth Sciences, 29, 844–858. [ Links ]
Dickson, J.A.D., 1965, A modified staining technique for carbonates in thin section: Nature, 205, 587. [ Links ]
Dunham R.J., 1962, Classification of carbonate rocks according to depositional texture. Memoirs – American Association of Petroleum Geologist, 1 (1962), pp. 108–121. [ Links ]
Ehrenberg, S.N., Eberli, G.P., Keramati, M., Moallemi, S.A., 2006, Porosity and permeability relationships in interlayered limestone–dolomite reservoir: American Association of Petroleum Geologists Bulletin, 90(1), 91–114. [ Links ]
Ehrenberg, S.N., Pickard, N.A.H., Laursen, G.V., Monibi, S., Mossadegh, Z.K., Svana, T.A., Aqrawi, A.A.M., McArthur, J.M., Thirlwall, M. F. , 2007, Strontium isotope stratigraphy of the Asmari Formation (Oligocene–Lower Miocene), SW Iran: Journal of Petroleum Geology, 30, 107–128. [ Links ]
Fakhari, M.D., Axen, G.J., Horton, B.K., Hassanzadeh, J., Amini, A., 2008, Revised age of proximal deposits in the Zagros foreland basin and implications for Cenozoic evolution of the High Zagros: Tectonophysics, 451, 170–185. [ Links ]
Flügel, E., 1982, Microfacies analysis of limestones. Berlin–Heidelberg, New York: Springer, 633 pp. [ Links ]
Flügel, E., 2004, Microfacies of carbonate rocks; analysis, interpretation and application: Berlin, Springer–Verlag, 976 pp. [ Links ]
Geel, T., 2000, Recognition of Stratigraphic sequence in carbonate platform and slope deposits: empirical models based on microfacies analyses of palaeogene deposits in southeastern Spain: Palaeogeography, Palaeoclimatology, Palaeoecology, 155(3), 211–238. [ Links ]
Hakimzadeh, S., Seyrafian, A., 2008, Late Oligocene–Early Miocene benthic foraminhfera and biostratigraphy of the Asmari Formation, south Yasuj, north–central Zagros basin, Iran: Carbonates and Evaporites, 23(1), 1–10. [ Links ]
Halfar, J., Godinez–Orta, L., Mutti, M., Valdez–Holguin, J.E., Borges, J.M., 2004, Nutrient and temperature controls on modern carbonate production: an example from the Gulf of California, Mexico: Geology, 32, 213–216. [ Links ]
Hallock, P., 1988, Diversification in algal symbiont–bearing foraminifera: a response to oligotrophy?: Revue de Paleobiologie, Special 2, 789–797. [ Links ]
Hallock, P., Glenn, E.C., 1986, Larger foraminifera: a tool for paleoenvironmental analysis of Cenozoic carbonate depositional facies: Palaios, 1(1), 55–64. [ Links ]
Heydari, E., 2008, Tectonics versus eustatic control on supersequences of the Zagros Mountains of Iran: Tectonophysics, 451, 56–70. [ Links ]
Hohenegger, J., 2000, Coenoclines of larger foraminifera: Micropaleontology, 4(1), 127–151. [ Links ]
Hohenegger, J., Yordanova, E., Hatta, A., 2000, Remarks on West Pacific Nummulitidae (Foraminifera): Journal of Foraminiferal Research, 30, 3–28. [ Links ]
Holzmann, M., Hohenegger, J., Hallock, P., Piller, W.E., Pawlowski, J., 2001, Molecular phylogeny of large miliolid foraminifera (Soritacea Ehrenberg 1839: Marine Micropaleontology, 43, 57–74. [ Links ]
International Commision on Stratigraphy (ICS), 2010, International Stratigraphic Chart: <https://engineering.purdue.edu/Stratigraphy/charts/chart.html> [ Links ].
Jalali, M.R., 1987, Stratigraphy of Zagros basin: National Iranian Oil Company, Exploration and Production Division, Report, 1249, 1072. [ Links ]
James, G.A., Wynd, J.G., 1965, Stratigraphic nomenclature of Iranian oil consortium, agreement area: American Association of Petroleum Geologists Bulletin, 49(12), 2182–2245. [ Links ]
James, N.P., 1997, The cool–water carbonate depositional realm, in James, N.P., Clarke, J., (eds.) Cool–water Carbonates: SEPM Special. Publications, 56, 1–20. [ Links ]
Kalantari, A., 1986, Microfacies of carbonate rocks of Iran: National Iranian Oil Company, Geological Laboratory Publication, Tehran, 520 pp. [ Links ]
Langer, M.R., Hottinger, L., 2000, Biogeography of selected "larger" foraminifera: Micropaleontology, 46(1), 57–86. [ Links ]
Laursen, G.V., Mobini, S., Allan, T.L., Pickard, N.A.H., Hosseiney, A., Vincent, B., Hamon, Y., Van–Buchem, F.S.P., Moallemi, A., Druillion, G., 2009, The Asmari Formation revisited: changed stratigraphic allocation and new biozonation: Shiraz, First International Petroleum Conference and Exhibition, European Association of Geoscientists and Engineers. [ Links ]
Lee, J.J., 1990, Fine structure of the rhodophycean Porphyridium purpureum in situ in Peneroplis pertusus (Forskal) and P. acicularis (Batsch) and in axenic culture: Journal of Foraminiferal Research, 20(2), 162–169. [ Links ]
Lees, A., 1975, Possible influence of salinity and temperature on modern shelf carbonate sedimentation: Marine Geology, 19, 159–198. [ Links ]
Lees, A., Buller, A.T., 1972, Modern temperate water and warm water shelf carbonate sediments contrasted: Marine Geology, 13, 1767–1773. [ Links ]
Lees, G.M., Richardson, F.D.F., 1940, The geology of the oil field belt of SW Iran and Iraq: Geology Magazine, 76(3), 227–252. [ Links ]
Leutenegger, S., 1984, Symbiosis in benthic foraminifera, specificity and host adaptations: Journal of Foraminiferal Research, 14(1), 16–35. [ Links ]
Lucia, F.J., 2007, Carbonate reservoir characterization: Springer, 334 pp. [ Links ]
Mossadegh, Z.K., Haig, D.R., Allan, T., Adabi, M.H., Sadeghi, A., 2009, Salinity changes during Late Oligocene to Early Miocene Asmari Formation deposition, Zagros Mountains, Iran: Palaeogeography, Palaeoclimatology, Palaeoecology 272, 17–36. [ Links ]
Motiei, H., 1993, Stratigraphy of Zagros: Geological Survey of Iran, 583 pp. [ Links ]
Murray, J.W., 1991, Ecology and palaeoecology of benthic foraminifera: New York, Wiley and Sons, 397 pp. [ Links ]
Pomar, L., Hallock, P., 2007, Changes in coral–reef structure through the Miocene in the Mediterranean province: Adaptive vs. environmental influence: Geology, 35, 899–902. [ Links ]
Purser, B.H., Tucker, M.E., Zenger, D.H.,1994, Problems, progress and future research concerning dolomites and dolomitization, in Purser, B.H., Tucker, M.E., Zenger, D.H., (eds.), Dolomites – A Volume in Honour of Dolomieu: Oxfrd, Blackwell Scientific Publications, Internationa Association of Sedimentologists, Special Publication, 21, 3–20. [ Links ]
Racey, A., 1995, Biostratigraphy and palaeobiogeographic significance of Tertiary nummulitids (foraminifera) from northern Oman: Micropaleotology, 41, 1–123. [ Links ]
Richardson, R.K., 1924, The geology and oil measures of southwest Persia: Jounal Institute Petroleum Technology, 10, 256–283. [ Links ]
Romero, J., Caus, E., Rossel, J., 2002, A model for the palaeoenvironmental distribution of larger foraminifera based on Late Middle Eocene deposits on the margin of the south Pyrenean basin: Palaeogeography, Palaeoclimatology, Palaeoecology, 179, 43–56. [ Links ]
Sadeghi, R., Vaziri–Moghaddam, H., Taheri, A., 2009, Biostratigraphy and Palaeoecology of the Oligo–Miocene succession in Fars and Khuzestan areas (Zagros Basin, SW Iran): Historical Biology, 21(1–2), 17–31. [ Links ]
Seyrafian, A., Hamedani, A., 2003, Microfacies and paleoenvironmental interpretations of the lower Asmari Formation (Oligocene), north–central Zagros basin, Iran: Neues Jahrbuch fur Geologie und Palaontologie, Monatshefte, 3,164–174. [ Links ]
Seyrafian, A., Mojikhalifeh, A.R., 2005, Biostratigraphy of the Late Paleogene–Early Neogene succession, north–cental border of the Persian Gulf: Carbonates and Evaporites, 20(1), 91–97. [ Links ]
Seyrafian, A., Vaziri–Moghaddam, H., Torabi, H., 1996, Biostratigraphy of the Asmari Formation, Burujen area, Iran: Journal of Sciences, Islamic Rebublic of Iran, 7(1), 31–48. [ Links ]
Sherkati, S., Letouzey, J., Frizon de Lamotte, D., 2006, The central Zagros fold–thrust belt (Iran): New insights from seismic data, field observation and sandbox modeling: Tectonics, 25, 1–27. [ Links ]
Sibley, D.F., Gregg, J.M., 1987, Classification of dolomite rock textures: Journal of Sedimentary Petrology, 57 (6), 967–975. [ Links ]
Thomas, A.N., 1948, The Asmari Limestone of southwest Iran: National Iranian Oil Company, Report 706, unpublished. [ Links ]
van Buchem, F.S.P., Allan, T.L., Laursen, G.V., Lotfpour, M., Moallemi, A., Monibi, S., Motiei, H., Pickard, N.A.H., Tahmasbi, A.R., Vedrenne , V., Vincent, B., 2010, Regional stratigraphic architecture and reservoir types of the Oligo–Miocene deposits in the Dezful Embayment (Asmari and Pabdeh Formations) SW Iran: Geological Society, London, Special Publications, 329, 219–263. [ Links ]
Vaziri–Moghaddam, H., Kimiagari, M., Taheri, A., 2006, Depositional environment and sequence stratigraphy of the Oligocene–Miocene Asmari Formation in SW Iran, Lali Area: Facies, 52(1), 41–51. [ Links ]
Wilson, J.L., 1975, Carbonate facies in geological history: New York, Springer, 471 pp. [ Links ]
Wynd, J.G., 1965, Biofacies of the Iranian consortium agreement area: Iranian Oil Offshore Company Report 1082, unpublished. [ Links ]

