










Services on Demand
Journal

Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
 Cited by SciELO
Cited by SciELO Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkRevista mexicana de ciencias geológicas
On-line version ISSN 2007-2902Print version ISSN 1026-8774
Rev. mex. cienc. geol vol.27 n.3 Ciudad de México Dec. 2010
Los Xenarthra (Mammalia) del Ensenadense (Pleistoceno inferior a medio) de la Región Pampeana (Argentina)
The Xenarthra (Mammalia) from the Ensenadean (lower to middle Pleistocene) of the Pampean region (Argentina)
Esteban Soibelzon1*, Angel Ramón Miño–Boilini2**, Alfredo Eduardo Zurita2 y Cecilia Mariana Krmpotic1
1 División Paleontología de Vertebrados, Museo de La Plata, Facultad de Ciencias Naturales y Museo, UNLP, Paseo del Bosque s/n, 1900 La Plata. CONICET. Argentina. * Correo electrónico: esoibelzon@fcnym.unlp.edu.ar
2 Centro de Ecología Aplicada del Litoral (CECOAL–CONICET), Ruta 5, Km. 2,5 (3400), Corrientes, Argentina. ** Correo electrónico: angelmioboilini@yahoo.com.ar
Manuscrito recibido: Diciembre 30, 2009.
Manuscrito corregido recibido: Mayo 17, 2010.
Manuscrito aceptado: Junio 7, 2010.
RESUMEN
Los Xenarthra (Mammalia) constituyen un grupo monofilético, caracterizado por una arquitectura esqueletaria particularmente distinta a la de los demás mamíferos placentarios. Su distribución actual es casi exclusivamente Neotropical; sin embargo, el registro fósil indica una distribución más amplia en el continente americano. En este trabajo se proporciona un listado actualizado de los Xenarthra registrados para el Pleistoceno inferior a medio (Piso Ensenadense) en la región pampeana (Argentina). Por otra parte, se describen brevemente las características anatómicas más conspicuas que permitieron la identificación taxonómica de los restos fósiles, en especial de aquéllos de interés bioestratigráfico. Asimismo se figuran los Cingulata presentes en el Ensenadense. Se estudiaron más de 700 ejemplares alojados en su gran mayoría en colecciones paleontológicas de la Argentina. El estudio de estos ejemplares permitió comprobar que durante el Pleistoceno existieron marcadas variaciones en la composición y distribución de la fauna pampeana en general y de los xenartros en particular, hecho íntimamente relacionado al intercambio biótico americano y a las variaciones climáticas producidas durante el Pleistoceno. De esta manera, en el Ensenadense se registran taxones exclusivos de este piso y otros que poseen su primer y/o último registro fósil durante este periodo. Se espera que este trabajo sirva de base para las futuras contribuciones respecto a la fauna pleistocénica de la región pampeana, así como del resto de la Argentina y América del Sur.
Palabras clave: bioestratigrafía, sistemática, Tardigrada, Cingulata, Cuaternario, América del Sur.
ABSTRACT
Xenarthra is a monophyletic group, characterized by a particular skeletal architecture, different to other placental mammals. Its current distribution is exclusively Neotropical; however, the fossil record indicates a greater distribution. The aim of this paper is to provide an updated list of the Xenarthra of the lower to middle Pleistocene (Ensenadan Stage) of the Pampean Region (Argentina). The most conspicuous anatomical features that allowed the taxonomic identification offossil remains, especially those of particular biostratigraphic interest, are also briefly described andfigured here. We studied more than 700fossil remains mostly housed in palaeontological collections of Argentina. The study of these specimens (from the Pampean Region), revealed that during the Pleistocene there were marked variations in the composition and distribution ofthe faunas in general and particularly in xenarthrans. This situation was closely related to the Great American Biotic Interchange and climatic oscillations occurring in the Pleistocene. Thus, the Ensenadan fossil record of xenarthrans consists on exclusive taxa and numerous cases of first and last records. Finally, we expect that this contribution provide a starting pointforfuture contributions in respect of the Pleistocene fauna, not only from the Pampean Region, but also from the rest of Argentina and South America.
Key words: biostratigraphy, systematic, Cingulata, Tardigrada, Quaternary, South America.
INTRODUCCIÓN
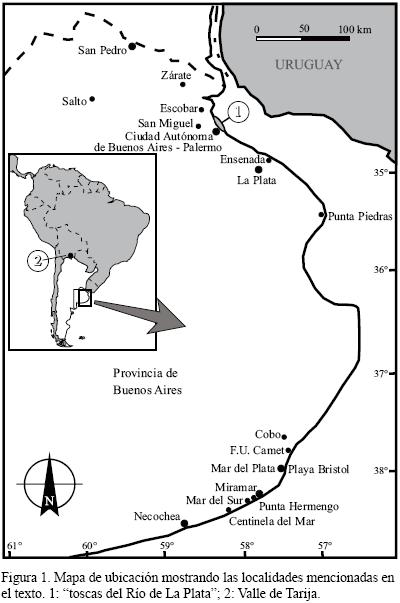
La presente contribución es parte de los trabajos de Tesis Doctoral de los autores (Zurita, 2007; Soibelzon, 2008; Krmpotic, 2009) y tiene como objetivo proporcionar un listado completo y actualizado de los Xenarthra (Mammalia) registrados durante el Ensenadense (Pleistoceno inferior a medio; Cione y Tonni, 2005; Soibelzon et al., 2008a) en la región Pampeana (Argentina, Figura 1). Asimismo, se brindan breves descripciones anatómicas, comentarios taxonómicos y sistemáticos de los taxones menos conocidos y/o de aquellos más problemáticos respecto a su procedencia geográfica y/o estratigráfica. Es importante remarcar que en este trabajo se figuran los osteodermos de todas las especies de Cingulata registrados en el Ensenadense, lo cual constituye por sí mismo un valioso aporte. Adicionalmente se brinda la distribución geográfica de los taxones (i. e., Dasypodidae) que se registran desde el Ensenadense a la actualidad.
El Ensenadense es una unidad cronoestratigráfica (Piso) definida para la región pampeana (Argentina) y basada en la biozona de Mesotherium cristatum de Cione y Tonni (2005). Según Soibelzon et al. (2008a), los sedimentos que incluyen fósiles característicos de esta biozona abarcan desde la parte inferior del Cron Brunhes (0.78 Ma) hasta más de 0.98 Ma (subcron Clrln). La base del Ensenadense, caracterizada bioestratigráficamente, es actualmente desconocida, pero se la ubica tentativamente en torno a los 2 Ma (ver Soibelzon et al., 2008a). Por otro lado, Verzi et al. (2004) ubican tentativamente el límite entre el Piso Ensenadense y Bonaerense en el comienzo del estadio isotópico 11 (ca. 0.40 Ma). El Piso Ensenadense, así definido, incluye los sedimentos depositados durante la Edad Ensenadense (entre ca. 1.95 Ma y más de 0.40 Ma).
El estudio de vertebrados fósiles en la Argentina tiene una larga tradición, conociéndose menciones aisladas provenientes de diversos sectores del territorio, tales como el hallazgo de una muela de un "gigante" en la ciudad de Córdoba (ver Lizarraga, 1916). En lo que concierne a los xenartros, su primera aparición en la bibliografía paleontológica corresponde a un esqueleto completo de Megatherium americanum Cuvier, 1796, procedente de las barrancas del río Luján (Buenos Aires, Argentina; ver Cuvier, 1796).
Los Xenarthra son considerados un grupo monofiléti–co (Gaudin, 2003, 2004). Los resultados obtenidos a través del empleo de metodologías moleculares son consistentes con esta interpretación (Delsuc et al., 2001, 2002; Delsuc y Douzery, 2009; Goloboff et al., 2009); sin embargo, su relación con los demás mamíferos placentarios es aún controversial (Delsuc y Douzery, 2008). En los Xenarthra se reconocen tres grupos (Delsuc et al., 2003; McDonald y Naples, 2007; Delsuc y Douzery, 2008): a) Cingulata (provistos de una coraza o armadura ósea); b) Vermilingua (marcadamente adaptados a la mirmecofagia); c) Tardigrada o Folivora (de hábitos terrestres y arborícolas, de los cuales en muchos casos se han preservado osteodermos subcutáneos). La distribución de los representantes vivientes del grupo es prácticamente neotropical, a excepción de Dasypus novencinctus (Linnaeus, 1758) o "armadillo de nueve bandas" que se registra hasta el centro del actual territorio de los Estados Unidos (Nowak, 1999).
Desde una perspectiva paleontológica, los Xenarthra son los mamíferos de registro más frecuente en la fauna pleistocena pampeana; entre ellos se encuentran taxones con importancia bioestratigráfica (e.g., Glyptodon munizi, Neosclerocalyptus pseudornatus) e indicadores paleoam–bientales (e.g., Tolypeutes matacus) (ver Soibelzon et al., 2006a; Soibelzon et al., 2008b).
Algunos listados taxonómicos de la fauna de xenartros ensenadenses se basaron, en parte, en materiales extremadamente fragmentarios y/o de dudosa procedencia estratigráfica (e.g., Gervais y Ameghino, 1880; Ameghino, 1889; Rusconi, 1936; Cione y Tonni, 1995, 1999; Carlini y Scillato–Yané, 1999), resultando así en una evidente sobreestimación del registro de xenartros pampeanos, situación que comienza a ser revertida a través de revisiones sistemáticas recientes (ver Zurita, et al., 2005, 2006, 2009a y 2009b; Soibelzon et al., 2006a y 2006b; Cruz et al., 2007; Krmpotic y Scillato–Yané, 2007; Brandoni et al., 2008; Krmpotic et al., 2009a; Miño–Boilini y Carlini, 2009; Miño–Boilini et al., 2009).
De acuerdo con las recientes revisiones realizadas por los autores mencionados en el párrafo anterior, gran parte de los taxones han sido sinonimizados. De esta manera, el registro ensenadense de Xenarthra se compone de un total de 22 géneros, con algunas especies con su primer registro fósil [e.g., Chaetophractus vellerosus (Gray, 1865)] y otras exclusivas de este piso (e.g., Glyptodon munizi Ameghino, 1881; Megatherium gallardoi Ameghino y Kraglievich, 1921).
MATERIALES Y MÉTODOS
Este trabajo se basó tanto en el estudio de ejemplares fósiles depositados en diversas colecciones paleontológicas de Argentina, España y Francia, como en trabajos de revisión sistemática y taxonómica publicados durante la década de 1990 (que constituyeron la primera revisión general de los mamíferos pampeanos desde tiempos de Ameghino; e.g., Scillato–Yané et al., 1995; Carlini y Scillato–Yané, 1999; Cione et al., 1999). Los reportes pertinentes fueron utilizados como punto de partida para la presente revisión. A esta información se agregó la proveniente de las recientes revisiones sistemáticas que serán citadas oportunamente. Cabe señalar que los listados faunísticos de las "toscas del Río de La Plata" (Buenos Aires, Argentina) son los presentados en Soibelzon et al. (2008b). Respecto a los ejemplares procedentes de la localidad de Centinela del Mar, es necesario aclarar que si bien no se han hallado fósiles característicos del Ensenadense, los niveles basales (A–B) de polaridad reversa (sensu Soibelzon et al., 2009) podrían ser referidos a esta edad. De esta manera, en el presente trabajo sólo se mencionarán los ejemplares procedentes de alguno de estos dos niveles.
En este trabajo seguimos el esquema sistemático de McKenna y Bell (1997) y el presentado como addenda en Alberdi et al. (1995). Debido a la cantidad de ejemplares implicados en este estudio (más de 700), sólo se mencionarán en detalle aquellos que presenten alguna particularidad, ya sea por su escasa frecuencia en el registro fósil o por su valor bioestratigráfico.
Abreviaturas. Col. Rusconi: Colección Rusconi (en parte), Museo J. C. Moyano (Mendoza, Argentina); IFG: Colección Henning, Museo Florentino y Carlos Ameghino (Santa Fé, Argentina); FML: Colección Rusconi (en parte), Instituto "Miguel Lillo" (Tucumán, Argentina); GCF y MSP: Grupo Conservacionista de Fósiles, Museo Paleontológico Municipal "Fray Manuel de Torres" (San Pedro, Buenos Aires, Argentina); MLP: División Paleontología Vertebrados, Museo de La Plata, (La Plata, Buenos Aires, Argentina); MACN: Sección Paleontología de Vertebrados, Museo Argentino de Ciencias Naturales "Bernardino Rivadavia" (Ciudad Autónoma de Buenos Aires, Argentina); MMP: Museo de Ciencias Naturales de Mar del Plata "Lorenzo Scaglia" (Mar del Plata, Buenos Aires, Argentina); MNHN PAM: Museum National d'Histoire Naturelle (Paris, Francia); MPV: Colección Botet, Museo Municipal de Valencia (Valencia, España).
TAXONES RECONOCIDOS PARA EL ENSENADENSE: TAXONOMÍA, PROCEDENCIA GEOGRÁFICA, DESCRIPCIÓN Y COMENTARIOS
Superorden Xenarthra Cope, 1889
Orden Cingulata Illiger, 1811
La característica anatómica más conspicua de este grupo es la presencia de una coraza dorsal que protege al cuerpo. Adicionalmente existe un escudo cefálico y un estuche caudal. También existen otras zonas en donde pueden hallarse osificaciones, como en el tegumento del rostro, en la región ventral del cuerpo y en las extremidades. Estas osificaciones no forman verdaderas corazas y han sido observadas y descriptas en algunos Cingulata fósiles (ver Burmeister, 1866; Lydekker, 1887; Tauber y Di Ronco, 2000; Soibelzon et al., 2006a; Zurita et al., 2010). Los osteodermos que conforman la coraza dorsal poseen en la cara expuesta una morfología particular y diferente en cada taxón, la cual ha sido la base para la sistemática del grupo. Como señalan Krmpotic et al. (2009b) los elementos más frecuentemente conservados en el registro fósil de los Cingulata corresponden a osteodermos sueltos, lo cual dificulta su asignación específica. De esta manera, el hallazgo de ejemplares completos o en buen estado de preservación (e.g., MPV 50–64 y MMP 3985 –ver apartado Eutatini y Glyptodontinae, respectivamente) permitió en muchos casos una nueva descripción y caracterización de numerosas especies.
Familia Dasypodidae Gray, 1821
Los dasipódidos (comúnmente denominados "armadillos") constituyen aproximadamente el 67% de los xenartros actuales. En la Argentina están representados por ocho géneros y 21 especies (Wetzel, 1982, 1985; Nowak, 1999; Parera, 2002), distribuidos en tres subfamilias (Euphractinae, Dasypodinae y Tolypeutinae; McKenna y Bell, 1997). A excepción del género Chlamyphorus Harlan, 1825, su coraza se divide en tres regiones claramente diferenciables: un escudo escapular, una región de bandas móviles (de número variable) y un escudo pélvico (Figura 2a). Cabe destacar que algunos representantes fósiles carecían de escudo escapular o éste se encontraba limitado a los flancos, quedando la región media de la coraza dorsal constituida en su totalidad por osteodermos móviles.
Subfamilia Euphractinae Winge, 1923
Tribu Euphractini Winge, 1923
Descripción y comentarios. Numerosos ejemplares componen el registro ensenadense de esta tribu. La mayor problemática radica en la asignación específica de estos ejemplares, ya que la ornamentación de los osteodermos es muy similar en los distintos taxones reconocidos. En este sentido resulta importante enumerar dos diferencias morfológicas en los osteodermos móviles de Chaetophractus y Zaedyus que permiten una separación clara de los dos géneros: 1) en Chaetophractus la zona de separación entre la tecla y la parte posterior del osteodermo está bien desarrollada y con superficie rugosa, mientras que es lisa en Zaedyus; 2) en Chaetophractus el borde posterior de los osteodermos es recto y presenta numerosos forámenes pilíferos en Chaetophractus, mientras que en Zaedyus este borde es delgado con escasos forámenes pilíferos y el foramen más notable coincide con la figura central (Vizcaíno y Bargo, 1993; ver Figura 2 en Soibelzon et al., 2006b).
Por último, el ejemplar MLP 69–VIII–9–5 (fémur derecho incompleto) asignado a Euphractus sexcinctus Linnaeus, 1758 e incluido en los listados faunísticos para el Ensenadense (ver Scillato–Yané, 1982; Carlini y Scillato–Yané, 1999; Cione et al., 1999) carece de datos fehacientes de procedencia.
Chaetophractus Fitzinger, 1871
Descripción y comentarios. Las dos especies registradas en el Ensenadense poseen un patrón similar de ornamentación en los osteodermos, siendo C. villosus (Desmarest 1804) aproximadamente un 13% mayor que C. vellerosus (Gray, 1865). Los osteodermos móviles de la coraza presentan una figura central y figuras laterales bien definidas (en número de 4 ó 5) siendo la más posterior de mayor tamaño (ver fig. 2 en Soibelzon et al., 2006b). La zona de separación entre la tecla y la parte ornamentada posee superficie rugosa.
Chaetophractus villosus (Desmarest 1804) Fitzinger 1871 (Figura 2b)
Materiales y procedencia. Las "toscas del Río de La Plata" (Col. Henning 163, MACN 8828), Necochea niveles PN9 a PN7 de Soibelzon et al. (2009) (MLP 04–VII–1–48, 04–VII–1–54, 04–VII–1–60).
Comentarios. Chaetophractus villosus se registra desde el Chapadmalalense (Plioceno medio; Scillato–Yané, 1982; Carlini y Scillato–Yané, 1999) a la actualidad. Se distribuye en Bolivia, Paraguay, Argentina y Chile (Gardner et al., 2005). En particular en la Argentina, se conoce con certeza desde la provincias de Salta y Chaco descendiendo hacia el sur por una franja central que se expande a la altura de la provincias de Santa Fe y Córdoba, llegando prácticamente hasta el contrafuerte andino en la provincia de Mendoza y el Atlántico en la provincia de Buenos Aires siguiendo hasta el extremo sur de la Patagonia argentina, excepto la cordillera (Parera, 2002). También se ha corroborado recientemente su presencia en Tierra del Fuego por introducción antrópica (Poljak et al., 2007).
Krmpotic et al. (2009b) indican que la superficie externa de los osteodermos fijos consiste en una figura central alargada y pequeñas figuras periféricas, al tiempo que la figura central ocupa los dos tercios posteriores del osteodermo. Las figuras periféricas son anteriores y laterales con respecto a la figura central y alrededor de la figura central hay pequeños forámenes. El borde posterior de los osteodermos posee pequeños forámenes pilíferos. Los osteodermos móviles presentan, en su porción caudal, una ornamentación similar a los osteodermos fijos pero en general carecen de figuras anteriores (ver figs. 1B, 1C, 2A, y 2B en Krmpotic et al., 2009b). Los osteodermos del escudo cefálico son esculpidos (Scillato–Yané, 1982).
Chaetophractus vellerosus (Gray, 1865) Yepes, 1928 (Figura 2b)
Materiales y procedencia. MLP 94–I–1–22 (cuatro osteodermos semimóviles de la primer hilera del escudo pélvico y un fragmento de otro osteodermo), Sector B del perfil expuesto en Punta Hermengo (ver Tonni y Fidalgo, 1982; Soibelzon et al., 2009), referido originalmente a Zaedyus pichiy (ver Soibelzon et al., 2006b).
Descripción y comentarios. Como se mencionó anteriormente, los osteodermos de la coraza dorsal de C. vellerosus son muy similares a los de C. villosus, pero de menor tamaño; los osteodermos del escudo cefálico son lisos (Scillato–Yané, 1982). Esta especie tiene su primer registro fósil en el Ensenadense de la región pampeana. La distribución actual es disyunta, con un área principal continua que abarca el Gran Chaco y centro de la Argentina hasta la provincia de Mendoza, y una secundaria en latitudes medias de la región costera de la provincia de Buenos Aires (ver fig. 1 en Soibelzon et al., 2006b), habitando sobre sedimentos arenosos bien drenados de la Bahía de Samborombón (Soibelzon et al., 2007). Esta distribución disyunta es el resultado de una paleodistribución más amplia que comprendía un sector mayor del actual territorio de la provincia de Buenos Aires, durante el Pleistoceno tardío y parte del Holoceno (Carlini y Vizcaíno, 1987; Soibelzon et al., 2007).
Zaedyus Ameghino, 1889
Descripción y comentarios. Osteodermos del escudete cefálico sin ornamentación. Osteodermos de la coraza dorsal con una figura central y figuras periféricas bien convexas. Entre la figura central y la periférica se disponen los forámenes de la cara expuesta muy poco notorios. En el extremo posterior de los osteodermos se disponen los forámenes pilíferos siendo el coincidente con la figura central, el que posee mayor tamaño.
Zaedyuspichiy (Desmarest, 1804) Osgood, 1919 (Figura 2b)
Materiales y procedencia. Norte de Mar del Plata (MMP 1209 y MLP 91–IV–5–160, posiblemente U6–U5 de Soibelzon et al., 2009), Mar del Sur (MLP 91–X–20–5, nivel B de Soibelzon et al., 2009), y Necochea (MLP 91–IV–25–70, 91–IV–25–71, 91–IV–25–74, 91–IV–25–47, 91–IV–25–187, 04–VII–1–40, 04–VII–1–50, 04–VII–1–57, 04–VII–1–71; niveles basales de Punta Negra y Las Grutas). El ejemplar 96–VIII–4–29 exhumado en Centinela del Mar proviene del nivel C, el cual podría ser de antigüedad ensenadense (según Soibelzon et al., 2009).
Comentarios. Z. pichiy tiene su primer registro fósil en el Ensenadense de la región pampeana; actualmente se distribuye en el centro y sur de Argentina (provincias de Mendoza, San Luis, sudoeste de Buenos Aires y la Patagonia argentina; ver Nowak, 1999).
En esta especie, las placas móviles de la coraza poseen una figura central y figuras laterales claramente diferenciables, la zona de separación entre la tecla y la porción ornamentada está menos desarrollada que en Chaetophractus y no posee superficie rugosa. Al igual que en Chaetophractus, existen pequeños forámenes en el sitio donde el vértice posterior de cada figura lateral intersecta el surco de separación de las figuras laterales con la central. El borde posterior de estos osteodermos es muy delgado y con escasos forámenes pilíferos; el foramen más notable coincide con la figura central (Vizcaíno y Bargo, 1993; ver fig. 2 en Soibelzon et al., 2006b).
Tribu Eutatini Bordas, 1933
Descripción y comentarios. Geográficamente ésta es una de las tribus de dasipódidos que está mejor representada en las unidades estratigráficas cenozoicas del Sur de América del Sur, esencialmente en la Argentina (Scillato–Yané, 1980; Scillato–Yané et al., 1995) y en menor proporción en el Uruguay (Mones y Francis, 1973; Perea, 1998; Bostelmann et al., 2008) y en Chile (Croft et al., 2007). La presencia de molariformes con tres capas de dentina bien diferencias y de grandes forámenes pilíferos (vinculados con una abundante pilosidad) se encuentran entre los caracteres morfológicos más conspicuos que caracterizan a los Eutatini. En el Ensenadense esta tribu está representada únicamente por el género Eutatus. Los materiales más completos que se conservan en las colecciones paleontológicas revisadas permiten reconocer una única especie para el Ensenadense, cuyo alcance geocronológico se extendería desde el Vorohuense hasta el Ensenadense (Krmpotic et al., 2009a). Lamentablemente, gran parte de los especímenes depositados en las colecciones paleontológicas corresponden a restos fragmentarios que imposibilitan su asignación específica.
Eutatus Gervais, 1867
Materiales y procedencia. Las "toscas del Río de La Plata" (ver tab. 1 en Soibelzon et al, 2008a), La Plata (MLP 82–IV–2–1), Punta Piedras (MACN 5675, MLP 82–IV–10–1, 86–IV–10–1), norte de Mar del Plata (MACN 5829, 10845, MMP 881, MLP 69–VIII–5–5), Miramar (MLP 40–VI–17–10, 52–IX–30–28, 69–VIII–1–8, 81–VII–20–6 y 06–II–2–6 provienen del sector B de Soibelzon et al., 2009), Mar del Sur (MLP 06–II–2–17, nivel A de Soibelzon et al., 2009) y Necochea (MLP 91–IV–25–66, 91–IV–25–81, 91–IV–25–86, 91–IV–25–90, 91–IV–25–149, 95–XII–1–26, 95–XII–1–27, 95–XII–1–28, 97–II–1–45; niveles basales de Punta Negra y Las Grutas). Los ejemplares MLP 91–IV–15–48, 04–VII–1–41, 04–VII–1–43, 04–VII–1–77 exhumados en Centinela del Mar provienen del nivel A, el cual podría ser de antigüedad ensenadense (según Soibelzon et al., 2009).
Comentarios. Eutatini de gran tamaño, similar al de Priodontes maximus. Su cráneo es muy alargado anteriormente y presenta gran desarrollo de los nasales y los premaxilares. La coraza dorsal se encuentra constituida por bandas nucales, un pseudoescudo escapular, bandas móviles y un escudo pélvico (Krmpotic, 2009). Tradicionalmente sólo Eutatus seguini Gervais, 1867 era conocido para el Ensenadense; sin embargo, recientes trabajos de revisión (Krmpotic y Scillato–Yané, 2007) indican que el material tipo de esta especie proviene del "pampeano superior" (sensu Ameghino, 1881), correspondiente al Lujanense del esquema pampeano (sensu Cione y Tonni, 2005).
Eutatus pascuali Krmpotic et al. 2009a (Figura 2b)
Materiales y procedencia. Playa Bristol, Mar del Plata (MMP 171–S), "toscas del Río de La Plata" (IFG 73, 131; MACN 5238, 8823, 10425, 10845, 11073; MMP 1212; MLP 52–X–2–27, 62–VII–27–3, 69–IX–5–3, 70–III–10–2; MPV 5064 (Krmpotic et al., 2009a).
Comentarios. Los osteodermos de la región central anterior del escudo pélvico presentan una figura central ancha rodeada de figuras periféricas anteriores y laterales. Los forámenes de la cara expuesta se disponen entre la figura central y las periféricas como finas puntuaciones. Generalmente presentan de 5 a 6 forámenes pilíferos.
Subfamilia Tolypeutinae Gray, 1865
Tolypeutes Illiger, 1811
Descripción y comentarios. Coraza dorsal sumamente convexa, con dos o tres bandas móviles entre el escudo escapular y el pélvico. Los osteodermos móviles presentan una figura central y dos laterales, constituidas por pequeños tubérculos. Los osteodermos fijos presentan una figura central rodeada adelante a los lados y atrás por figuritas periféricas, constituidas por pequeños tubérculos (Scillato–Yané, 1982).
Tolypeutes matacus (Desmarest, 1804) (Figura 2b)
Materiales y procedencia. Las "toscas del Río de La Plata" [MACN 980, 981, 5515, FML 904 o 268 de la numeración original brindada por Rusconi (1937), 905 o 361 de la numeración original brindada por Rusconi (1937) y 1428], ciudad de La Plata (MACN A 1479 y 2339), Miramar (MACN 13016, MLP 52–IX–28–49, 81–VII–20–2, 06–II–2–8 del nivel B de Soibelzon et al., 2009), Mar del Sur (MLP 06–II–2–18, nivel A de Soibelzon et al., 2009) y Necochea (91–IV–25–185, 91–IV–25–42, 97–II–1–42, niveles basales de Punta Negra y Las Grutas).
Comentarios. Frenguelli (1921) definió y caracterizó una especie como exclusiva del Ensenadense, Tolypeutes pampaeus, caracterizada por la presencia de tubérculos pequeños, numerosos y de vértice más agudo que los de la especie viviente T. matacus. Asimismo, los osteodermos de las bandas móviles presentan los tubérculos de mayor tamaño en la parte central y disminuyen de tamaño hacia los laterales. Kraglievich (1934) indica que los caracteres propuestos por Frenguelli (1921) entran dentro del rango de variación de la especie T. matacus. Más tarde Scillato–Yané (1980) incluye a T. pampaeus en el elenco de Dasypodidae de la Argentina y, posteriormente, esta especie sirvió de base a Cione y Tonni (1995) para definir la Biozona de Tolypeutes pampaeus, base bioestratigráfica del Piso Ensenadense. Según nuestros estudios, el material tipo de T. pampaeus (MLP 52–IX–28–49) y los materiales referidos a esta especie no muestran evidencia suficiente que permita separarlo de la especie actual, T. matacus. Consecuentemente, T. pampaeus sería sinónimo de T. matacus. No obstante, un trabajo de descripción (en preparación) de la coraza de Tolypeutes brindará un panorama más claro sobre la taxonomía de este grupo.
En la actualidad, T. matacus se distribuye en Bolivia, Sur de Brasil, el Chaco de Paraguay y Argentina (Nowak, 1999). En la Argentina en particular y siguiendo la clasificación de Cabrera y Willink (1980) se encuentra limitada biogeográficamente a la Provincia Chaqueña, del Espinal del Monte y Pampeana, todas ellas incluidas en el Dominio Chaqueño.
Subfamilia Dasypodinae Gray, 1821
Descripción y comentarios. Esta subfamilia está muy pobremente representada en el Ensenadense de la Región Pampeana e incluye sólo a Propraopus grandis Ameghino, 1881. Como carácter distintivo, los osteodermos fijos de la coraza dorsal poseen forma hexagonal a pentagonal, con una figura central grande, que ocupa la casi totalidad de la superficie del osteodermo, y pequeñas figuras periféricas desarrolladas sobre el borde craneal de la misma; los osteodermos móviles presentan una figura central lageniforme.
Scillato–Yané (1982) indica la presencia de Dasypus en sedimentos de la Formación Ensenada de la provincia de Buenos Aires; sin embargo éstos no fueron hallados en ninguna colección revisada, con lo cual no se incluye este taxón en el elenco ensenadense.
Propraopus Ameghino, 1881
Descripción y comentarios. Este género se registra en el Pleistoceno de Argentina, Bolivia, Brasil, Ecuador, Uruguay y Venezuela (Scillato–Yané, 1980; Marshall et al., 1984). Los osteodermos de la coraza dorsal son de gran tamaño y prácticamente duplican a los de Dasypus. Los ubicados en el escudo escapular y pélvico son isométricos y de forma hexagonal, la figura central es subcircular y se encuentra rodeada por dos o tres figuras periféricas. En la cara expuesta existen tres o cuatro forámenes pilíferos, localizados sobre el surco central y entre los surcos periféricos. Los osteodermos de las bandas móviles son alongados antero–posteriomente, la figura central se encuentra delimitada por dos surcos divergentes de forma triangular cuya base está dirigida hacia adelante (otorgándole el típico aspecto lageniforme observable en los osteodermos móviles de las especies incluidas en la tribu Dasypodini Gray, 1821).
Propraopus grandis Ameghino, 1881 (Figura 3a)
Materiales y procedencia. Las "toscas del Río de La Plata" (MACN 8826, 8830), ciudad de La Plata (MACN 1170, MLP 52–X–1–43), norte de Mar del Plata (MACN 13017, Tipo de Propraopus minor) y de Necochea (MLP 91–IV–25–183).
Comentarios. Armadillo de gran tamaño, de talla comparable a la de Priodontes maximus y Eutatus seguini. Coraza dorsal compuesta por un escudo escapular, seis bandas móviles y un escudo pélvico. Tanto los osteodermos fijos como los móviles poseen forámenes de la cara expuesta en los surcos que separan la figura central de las periféricas (Scillato–Yané, 1982).
Familia Pampatheriidae Paula Couto, 1954
Pampatherium Gervais y Ameghino, 1880
Descripción y comentarios. Este género es de gran tamaño, presenta los osteodermos de la coraza dorsal con la figura central apenas esbozada o absolutamente indistinguible (Scillato–Yané, 1982; Scillato–Yané et al., 2005). La superficie expuesta de los osteodermos es muy uniforme, y la zona central es plana y carente de una figura central.
Pampatherium typum (Gervais y Ameghino, 1880) (Figura 3b)
Materiales y procedencia. El registro ensenadense de esta especie se limita a las "toscas del Río de La Plata" (MACN 5519, 6211, MLP 69–VIII–1–1, Col. Rusconi 51), ciudad de La Plata (MLP 54–III–16–1), Punta Piedras (MLP MACN 5679) y Mar del Plata (MMP 216, posiblemente U6 de Soibelzon et al., 2009).
Comentarios. Los grandes osteodermos de la coraza dorsal de P. typum poseen la superficie lisa y finamente punteada, sin el desarrollo de una figura central (ver Cattoi, 1966; Scillato–Yané et al., 2005). El registro de la familia Pampatheriidae es escaso en el Cuaternario de la región Pampeana, no así durante el Mioceno–Plioceno de la Mesopotamia y la región subandina, posiblemente relacionado a una conexión de estas regiones con áreas subtropicales (Carlini y Scillato–Yané, 1999). Entre los pampatéridos cuaternarios de América del Sur, Scillato–Yané et al. (2005) reconocen los géneros Holmesina Simpson, 1930 (con tres especies) y Pampatherium Ameghino, 1891 (con dos especies). Según estos autores, sólo dos especies se registran en la Argentina, Holmesina paulacoutoi (Cartelle y Bohorquez, 1985) en el Pleistoceno superior de las provincias de Formosa, Corrientes, Buenos Aires y Entre Ríos, y Pampatherium typum (Gervais y Ameghino, 1880) en el Pleistoceno Inferior a Superior de Corrientes, Formosa y Buenos Aires. Esta última especie es la forma más pequeña del grupo.
Familia Glyptodontidae Gray, 1869
Durante el Pleistoceno se registran por primera vez un total de 17 especies nominales de gliptodontes, agrupadas en tres subfamilias: Glyptodontinae Gray, 1869, Hoplophorinae Huxley, 1864 y Doedicurinae Ameghino, 1889; de éstas, 13 serían exclusivas del Ensenadense, entre las que se cuenta una de las de mayor tamaño, Panochthus intermedius Lydekker, 1894 (Scillato–Yané y Carlini, 1998a; Carlini y Scillato–Yané, 1999).
Subfamilia Glyptodontinae Gray, 1869
Glyptodon Owen, 1839
Materiales y procedencia. Glyptodon Owen, 1839 constituye uno de los taxones más conspicuos entre el conjunto de gliptodontes pleistocénicos de América del Sur (Paula Couto, 1979; Hoffstetter, 1958; Soibelzon et al., 2006a), con una amplia distribución latitudinal, desde el sur de la provincia de Buenos Aires hasta Venezuela probablemente (Carlini et al., 2008a, 2008b; Zurita et al., 2009b). Registros ensenadenses de Glyptodon incluyen las localidades de San Miguel (MLP 07–IX–2–1, nivel E3 de Soibelzon et al., 2008c), "toscas del Río de La Plata" (MACN 1771, 1856, 1889, 1897, 1899, 1956, 1990, 2012, 2078, 2079, 2110, 2184, 2193, 2269, 5419, 7061, MLP 54–VI–19–9), La Plata (54–III–16–2 , 71–VII–25–1 , 74–IV–26–4 , 80–IX–7–1), Punta Piedras (MACN 5670, 5680, 5692, 8689, MLP 16–150), norte de Mar del Plata (MACN 10841, MLP 06–II–2–15 de la U6 de Soibelzon et al., 2009, MMP 22–S, 122–S, 138–S, 139–S, 142–S, 230–S, 279–S, 289–S, 294–S, 333–S, 335–S, 772–S), Miramar (MLP 91–IV–30–24, 91–IV–30–33, 06–II–2–14, niveles A y B de Soibelzon et al., 2009), Mar del Sur (MLP 06–II–2–27, nivel A de Soibelzon et al., 2009) y Necochea (MLP 91–IV–25–30, 91–IV–25–67, 91–IV–25–69, niveles basales de Punta Negra y Las Grutas).
Descripción y comentarios. Glyptodon posee caracteres altamente especializados (Ameghino, 1889; Cattoi, 1966), aunque con una persistencia notable de ciertos rasgos primitivos, como por ejemplo la presencia de anillos caudales y de cinco dígitos en las extremidades (Hoffstetter, 1958). Los molariformes son trilobulados (a excepción del molariforme 1) (Soibelzon et al., 2006a) y la coraza está formada por osteodermos gruesos, con una figura central rodeada de una hilera de 5 a 7 figuritas periféricas conformando el característico diseño en "roseta" (Ameghino, 1889). Glyptodon incluye especies de gran tamaño, con una dieta principalmente pastadora (Fidalgo y Tonni, 1983; Pérez et. al., 2000; Vizcaíno et al., 2006).
Varios autores (e.g., Owen, 1839, 1845; Burmeister, 1866; Ameghino, 1881, 1883, 1889) han reconocido un gran número de especies incluidas en el género Glyptodon, fundamentalmente con base en fragmentos de la coraza dorsal, lo cual ha producido una sobreestimación de la diversidad del género (Duarte, 1997; Soibelzon et al., 2006a), fenómeno que ya había sido percibido por Ameghino (1889). Al igual que ha sucedido con otros grupos de Glyptodontidae, esta concepción netamente tipológica de especie es propia del siglo XIX (ver Mayr, 1996; Giraudo, 1997). Si bien son abundantes los restos atribuibles a Glyptodon sp. procedentes de niveles ensenadenses, su estado fragmentario (e.g., osteodermos aislados o asociados en pequeños grupos, huesos largos, costillas, entre otros) limita la posibilidad de una asignación específica correcta, teniendo en cuenta, además, la evidente variación morfológica observada en los osteodermos de las distintas regiones de la coraza dorsal. Por el momento no se han llevado a cabo revisiones sistemáticas modernas del género, aunque cabe resaltar los aportes de Duarte (1997) y Tonni y Berman (1988) sobre la diferenciación entre Glyptodon reticulatus Owen, 1845 y Glyptodon clavipesOwen, 1839; por su parte Rinderknecht (2000a) evaluó posibles variaciones ontogenéticas de los osteodermos de la coraza dorsal de Glyptodon y Soibelzon et al. (2006a) caracterizaron y describieron Glyptodon munizi.
Dentro de este complejo panorama taxonómico, para el Ensenadense se reconocieron históricamente cuatro especies: G. principalis Gervais y Ameghino, 1880 (posteriormente referida como Sclerocalyptus principalis por Ameghino, 1889), G. laevis Burmeister, 1866, G. gemmatum Nodot, 1857 y G. munizi (Ameghino, 1889; Carlini y Scillato–Yané, 1999; Cione et al., 1999; Cione y Tonni, 1999; Soibelzon et al., 2006a).
Glyptodon munizi Ameghino, 1889 (Figura 4a)
Materiales y procedencia. Restos atribuibles a esta especie han sido hallados en San Pedro (GCF 10, nivel E de la cantera Bigliochi–Iglesias, ver Soibelzon et al., 2006a y Soibelzon et al., 2008c), en las "toscas del Río de La Plata", frente a Vicente López (MACN 8706), en las proximidades de Mar del Plata (MMP 3985, ruta 2, kilómetro 389, próximo a la localidad de Cobo, 37°50'43.10"S y 57°36'57.13"W; ver más acerca de esta localidad en Soibelzon et al., 2001) y frente al Golf Club de Miramar (MLP 07–V–3–2; 200 m al sur de la "baliza Chica" de Miramar; 38°15'24.1''S y 57°47'55.5''W; ver Zurita et al., 2010). Por último el ejemplar MLP 07–IX–2–2 procedente de la localidad de San Miguel (Buenos Aires 34°31'58.8''S y 58°37'16.2'W) podría corresponder a esta especie. Este espécimen fue hallado en el nivel E1 de polaridad reversa, asignable a la parte final del Cron Matuyama (>0.78 Ma; ver fig. 2 en Soibelzon et al., 2008c).
Comentarios. Los hallazgos de una coraza dorsal (MMP 3985) y de un esqueleto casi completo (GCF 10) han brindado la oportunidad de completar significativamente, a nivel craneal y de la coraza dorsal, la descripción y caracterización morfológica de Glyptodon munizi (ver Soibelzon et al., 2006a). Adicionalmente, uno de los ejemplares (GCF 10) conserva restos de un plastrón ventral (estructura previamente citada por Burmeister, 1866, Lydekker, 1887 y Tauber y Di Ronco, 2000 para G. reticulatus) y un número significativo de osteodermos faciales (figs. 1C y 1D en Soibelzon et al., 2006a). Estructuras de este tipo sólo habían sido reportadas por Burmeister (1874) y Rinderknecht (2000b) en las extremidades posteriores de Glyptodon clavipes.
Subfamilia Doedicurinae Ameghino, 1889
Descripción y comentarios. Los Doedicurinae son uno de los Glyptodontidae de mayor tamaño, caracterizados por la presencia de osteodermos de la coraza dorsal que no forman figuras discretas y con perforaciones que atraviesan todo el espesor de la coraza; el tubo caudal es aplanado y notablemente ensanchado distalmente (Pascual et al., 1966).
El registro ensenadense de esta subfamilia incluye, además de Doedicurus (ver más adelante), dos especies: Daedicuroides eguiai (Ameghino, 1891) y Plaxhaplus ensenadensis (Ameghino, 1904). Castellanos (1940) incluyó en Daedicuroides unos ejemplares publicados por Ameghino (1891) procedentes del "pampeano inferior" de Mar del Plata. Ameghino (1904) funda Doedicurus ensenadensis Ameghino, 1904 con base en restos de un tubo caudal y de los osteodermos de la coraza dorsal, procedentes de las excavaciones del Puerto de La Plata. Posteriormente, Castellanos (1940) sinonimizó esta especie a Plaxhaplus ensenadensis. Cabe destacar que este último autor, "por causas ya conocidas" (Castellanos, 1940: 416) no pudo estudiar los materiales de Ameghino, sino que se basó en las descripciones brindadas por este investigador. No obstante, al momento de la elaboración de este trabajo, los restos atribuidos a estas especies y depositados en el MACN, no fueron hallados, con lo cual no se pudo aclarar esta situación.
Por último, cabe mencionar que el nombre genérico Daedicurus (Lydekker, 1894; Castellanos, 1940) fue mal escrito, debido a que existen suficientes evidencias de que se debió a un error de imprenta.
Doedicurus Burmeister, 1874 (Figura 4b)
Materiales y procedencia. Ciudad de La Plata (MLP 76–VI–26–1, 91–II–10–1, 99–VIII–1–2), Punta Piedras (MLP 55–V–3–2), Mar del Plata (MACN 6046; MMP 4–S, 87–S, 704–S; MLP 62–VII–27–103, 74–IV–22–1). El ejemplar MLP 96–III–11–6, procedente de niveles ensenadenses de la Cantera de Gorina no fue localizado al momento de la realización de este trabajo; por último, los ejemplares MACN 2274 (fémur incompleto) y 5084 (astrágalo) procedentes de las "toscas del Río de La Plata" podrían ser referidos a Doedicurus.
Descripción y comentarios. Osteodermos de la coraza de gran espesor y con tres a cinco forámenes muy notorios. Burmeister (1874) erigió este género sobre un ejemplar de gran tamaño (Glyptodon giganteus Serres, 1865 =Panochthus giganteus Burmeister, 1874) procedente de Salto (provincia de Buenos Aires), compuesto por varios elementos postcraneales (incluyendo el tubo caudal) y numerosos osteodermos que interpretó como pertenecientes a una coraza ventral (Gervais y Ameghino,1880, indican que estos osteodermos corresponden a la coraza dorsal). Asimismo, Burmeister (1866) destacó la presencia, en el cráneo, de algunos osteodermos pequeños correspondientes a osículos. Años más tarde, Burmeister (1879) fundó la nueva especie, Doedicurus giganteus (Serres, 1865) y después Gervais y Ameghino (1880) reconocieron tres nuevas especies: D. clavicaudatus, D. uruguayensis, D. poucheti con base en fragmentos de tubos caudales, a la vez que señalan que los ejemplares asignados por Burmeister (1879) no difieren claramente de las especies anteriormente mencionadas. Posteriormente, Ameghino (1889) colectó nuevos ejemplares en los alrededores de Luján (provincia de Buenos Aires) y analizó nuevamente los ejemplares sobre los que Burmeister (1879) propuso a la especie D. giganteus, concluyendo que el ejemplar procedente de Salto (provincia de Buenos Aires), más los nuevos materiales revisados correspondían a una nueva especie, Doedicurus kokenianus, registrada en los "pisos" "ensenadense, belgranense y base del bonaerense". Sin embargo, las procedencias geográficas de estos ejemplares no permiten asegurar con total certeza que dicha especie pudiera registrarse en el Ensenadense.
Burmeister (1866) atribuyó a Doedicurus clavi–caudatus (especie registrada en el Lujanense) una pelvis colectada en la provincia de Buenos Aires y depositada en el MNHN, sobre la que previamente se había erigido Glyptodon giganteus Serrés, 1865. Por su parte, Lydekker (1894) sinonimizó a D. kokenianus con D. clavicaudatus, mientras que Castellanos (1940) considera como válida a D. kokenianus.
Por lo expresado anteriormente, se desprende la necesidad de una minuciosa revisión taxonómica de las especies consideradas en el género Doedicurus y, por lo tanto, en este trabajo se optará por mantener la asignación de los ejemplares ensenadenses a nivel genérico.
Subfamilia "Hoplophorinae" Huxley, 1864
Tribu Lomaphorini Hoffstetter, 1958
Lomaphorus Ameghino, 1889 (Figura 4c)
Materiales y procedencia. El registro del género se limita a las "toscas del Río de La Plata" (MACN 1003, 1374, 5701, 5656, 13548; estos ejemplares fueron erróneamente atribuidos a Doedicurus por Soibelzon et al., 2008b), La Plata (MACN A 503, 1253, 1374, 1375) y Miramar (MACN 1023, MLP 07–V–3–19, nivel B de Soibelzon et al., 2009).
Descripciones y comentarios. El género Lomaphorus reúne algunas de las especies previamente incluidas en Hoplophorus Lund, 1839 (e.g., H. imperfectus Gervais y Ameghino, 1880, H. elevatus Ameghino, 1883 y H. compressus Ameghino, 1883). Este taxón posee osteodermos hexagonales y pentagonales, con una figura central circular, rodeada por una serie de figuritas periféricas poco notorias de superficie estriada. Para su "ensenadense", Ameghino (1889) reconoce Lomaphorus imperfectus (Gervais y Ameghino, 1880) y L. compressus (Ameghino, 1883), con base en osteodermos de la coraza dorsal; asimismo, describió un tubo caudal (posiblemente MACN 1003) procedente de las "toscas del Río de La Plata" y lo refierió a L. imperfectus. Ambas especies se diferencian, según Ameghino (1889), por su tamaño y forma general de los osteodermos y de la figura central; sin embargo, las características brindadas, así como los materiales figurados, no permiten una asignación específica para los restos ensenadenses.
Tribu Panochthini Simpson, 1945
Panochthus Burmeister, 1866
Materiales y procedencia. Los restos de Panochthus ensenadenses son escasos y se limitan a las "toscas del Río de La Plata" (MACN 1958 y 1959), La Plata (MACN A 403, MLP 84–IX–2–11 y 96–III–10–7).), alrededores de la CABA (MACN 10897), norte de Mar del Plata (MMP 141–S y 662–S) y Mar del Sur (MACN 7097).
Descripción y comentarios. El cráneo de Panochthus presenta un marcado descenso de la región fronto–nasal (ver Lydekker, 1894, lám. XXII), en tanto que la bóveda craneal es alta. Los osteodermos de la coraza dorsal presentan un gran número de tubérculos o figuritas pequeñas y en algunos casos es posible diferenciar una figura central. La coraza caudal está compuesta por seis o siete hileras de anillos y culmina en un tubo terminal deprimido, ensanchado en el extremo distal y con verrugas de diferentes tamaños en los laterales (Ameghino, 1889; Lydekker 1894). En particular, los osteodermos de los extremos de la coraza de P. intermedius Lydekker, 1894 presentan una figura central rodeada por varias hileras de pequeñas figuritas periféricas poligonales de tamaño semejante (mostrando cierta similitud a lo observado en los "Hoplophorinae" Plohophorini), mientras que a nivel de la región dorsal de la coraza ya no es posible diferenciar las figuras centrales y se observa en consecuencia un reticulado general (ver Lydekker, 1894: lám. XIX). Por su parte, en la especie Lujanense P. tuberculatus (Owen, 1845), las figuras centrales de los osteodermos ya desaparecen por completo (Scillato–Yané et al., 1995). Rusconi (1936) cita P. frenzelianus para el ensenadense de la provincia de Buenos Aires, pero este registro es dudoso; a su vez el material al que hace referencia se encuentra perdido con lo cual no se lo incluye en este trabajo.
Panochthus intermedius Lydekker, 1894 (Figura 4d)
Materiales y procedencia. Alrededores de la CABA (MLP 16–36 ejemplar tipo de la especie).
Comentarios. Respecto al ejemplar tipo de P. intermedius que proviene del "Pampeano del Puerto de Buenos Aires, conservado actualmente en el Museo de San Fernando" (Lydekker, 1894: 34), no ha sido hallado en dicha institución, sino que se encuentra en el Museo de La Plata (MLP16–36).
Panochthus subintermedius Castellanos, 1933
Materiales y procedencia. MACN 5130, Ensenadense de Palermo (Ciudad de Buenos Aires).
Comentarios. Si bien esta especie no es considerada en los listados generales de fauna pampeana (e.g., Cione et al., 1999; Carlini y Scillato–Yané, 1999), Cruz et al. (2007) indican que el material tipo (MACN 5130) no presenta diferencias con las especies mejor descriptas para el Bonaerense–Lujanense: P. tuberculatus (Owen) y P. frenzelianus Ameghino, 1889, en lo que respecta a la coraza dorsal, pero sí a nivel del tubo caudal, lo que sugiere que se trata de una especie válida.
Tribu Neuryurini Hoffstetter, 1958
Comentarios. La tribu Neuryurini fue reconocida y caracterizada por Hoffstetter (1958) y constituye dentro de los Glyptodontidae Hoplophorinae, uno de los grupos más pobremente conocidos (ver Hoffstetter, 1958; Cattoi, 1966; Paula Couto, 1979; Zurita et al., 2009c).
Neuryurus Ameghino, 1889
Descripción y comentarios. Este género se caracteriza por la presencia de osteodermos gruesos, mayormente rectangulares, y articuladas de modo laxo. La superficie expuesta de cada osteodermo está uniformemente perforada, de aspecto áspero, y con ausencia de ornamentación. En los osteodermos de mayor espesor puede observarse, en su cara ventral, la presencia de tres o cuatro evidentes forámenes de ubicación central (Pascual et al., 1966; Zurita y Ferrero, 2009).
Neuryurus rudis (Gervais, 1878) Ameghino 1889 (Figura 4e)
Materiales y procedencia. el registro de N. rudis es sumamente escaso en el Ensenadense y limitado al material tipo procedente de Santa Fe (MNHN s/n). El material tipo corresponde al ejemplar figurado en Ameghino (1889, lám. 61, fig. 10) y en la actualidad se encuentra reducido a menos de 20 osteodermos muy mal conservados (Vincent Pernègre –MNHN– comunicación personal, 2007); asimismo, existe un ejemplar procedente de Punta Hermengo referible a Neuryurus cf. N. rudis (MLP 91–IV–30–34; nivel A de de Soibelzon et al., 2009).
Descripción y comentarios. Hasta el momento constituye, junto con N. trabeculatus (ver Zurita y Ferrero, 2009), la única especie bien caracterizada de la tribu, limitada al Ensenadense de la región Pampeana (ver Ameghino, 1889: 314; Simpson, 1945: 74; Mones, 1986: 235; McKenna y Bell, 1997: 90; Zurita et al., 2006, 2009c). Cabe destacar que, según Castellanos (1940), en el tramo del río Carcarañá de donde provienen los restos del material tipo, no aflora el Ensenadense, sino que este material provendría del "belgranense", en consecuencia, de confirmarse este dato, N. rudis no sería exclusivo del Ensenadense.
Esta especie es de gran tamaño (comparable al de Panochthus Burmeister, 1866), tiene una coraza dorsal integrada por grandes osteodermos de superficie dorsal uniformemente perforada y sin clara evidencia de división en figuras y figuritas (ver Zurita et al., 2006; 2009c). Los osteodermos adyacentes a la escotadura caudal poseen una evidente convexidad (ver Ameghino, 1889; Hoffstetter, 1958; Cattoi, 1966; Paula Couto, 1979). El tubo caudal está formado por osteodermos similares a los de la coraza dorsal, unidos entre sí de modo laxo y presentando a los laterales "verrugas" elípticas con una elevación cónica en su parte central, mostrando cierta analogía con Panochthus y Hoplophorus Lund, 1839 (Ameghino, 1889, 1895; Hoffsetter, 1958; Paula Couto, 1979).
Tribu Neothoracophorini Castellanos, 1951
Comentarios. Los Neothoracophorini incluyen dos géneros muy mal caracterizados (ver Castellanos, 1951): Pseudothoracophorus Ameghino, 1881 y Neothoracophorus Ameghino, 1889. Este último género se registra en el Pleistoceno e incluye a N. elevatus (Nodot, 1857) en el "Ensenadense".
Lamentablemente no se han encontrado restos asignables a dicha especie en ninguna colección revisada, pese a que Ameghino (1889: 232) indica haber colectado restos en las "toscas del Río de La Plata". Sin embargo el material tipo de esta especie, según comentarios del propio Ameghino (1889: 231) se encontraría en el MNHN. Es importante remarcar que Zurita et al. (2009b) indican que en el Valle de Tarija, Bolivia (ver fig. 1 en Zurita et al., 2009b), los registros asignados a Neothoracophorus corresponden en realidad a ejemplares juveniles de Glyptodon.
Tribu Hoplophorini Huxley, 1864
Neosclerocalyptus Paula Couto, 1957 (Figura 4f)
Materiales y procedencia. Las "toscas del Río de La Plata" (FML 601, 1416, 559, IFG 1151), alrededores de la ciudad de Buenos Aires (FML 561 o 496 de la numeración original brindada por Rusconi, 1937) y de La Plata (MACN A 826, MLP 96–III–10–1), norte de Mar del Plata (MMP 143–S, 223–S, 233–S, 295–S) y Punta Hermengo (MLP 06–II–2–4, 06–II–2–11, nivel B de Soibelzon et al., 2009).
Descripción y comentarios. Las especies incluidas en este género (=Sclerocalyptus Ameghino, 1891; ver Zurita, 2007) son los Glyptodontidae pleistocénicos de menor tamaño (Carlini y Tonni, 2000). Se caracterizan por poseer una coraza dorsal alargada, subcilíndrica y de perfil dorsal casi totalmente recto, con los extremos anteriores desplegados a modo de "aletas". Los osteodermos son delgados y poseen un patrón de ornamentación en "roseta" de bajo relieve. La coraza caudal es larga y se compone de una serie de anillos caudales y finaliza en un tubo cílindrico–cónico (Zurita et al., 2005; Zurita, 2007). El cráneo presenta un notable desarrollo en la neumatización y expansión lateral de los senos fronto–nasales, un carácter único entre los Glyptodontidae (Zurita et al., 2009a). El carácter fragmentario de muchos de los ejemplares ensenadenses de Neosclerocalyptus no permite su asignación específica.
Neosclerocalyptus pseudornatus (Ameghino, 1889)
Materiales y procedencia. Las "toscas del Río de La Plata" (ver Zurita, 2007; Soibelzon et al., 2008b) y Mar del Plata (MMP 234).
Comentarios. Como señalan Zurita et al. (2005) y Zurita (2007), el cráneo de N. pseudornatus es de constitución primitiva dentro del clado Glyptodontidae, dado que los senos fronto–nasales están muy poco neumatizados. De modo sugerente, la aparición de este carácter coincide con la Gran Glaciación Patagónica (OIS 30–34) (ver Zurita et al., 2009a). En un contexto adaptativo, este carácter es interpretado como una posible respuesta a los ambientes predominantemente fríos y áridos del Pleistoceno (Zurita et al., 2005, 2009a)
Neosclerocalyptus ornatus (Owen, 1845)
Materiales y procedencia. San Pedro (MSP 12) y Mar del Plata (MACN 8091, MLP 16–28, MMP 4300) y en un sector de las barrancas del Río Paraná, Granadero Baigorria, en la provincia de Santa Fe (IFG 656). Cabe destacar que los numerosos restos atribuidos por Rusconi (1937) a N. ornatus y procedentes de excavaciones realizadas en la ciudad de Buenos Aires no fueron hallados en las colecciones revisadas.
Comentarios. Desde un punto de vista estratigráfico, este taxón parece tener un registro más tardío que N. pseudornatus, limitado a la parte tardía del Ensenadense (ca. 0.98–0.40 Ma; Soibelzon et al., 2008b; Zurita et al., 2009a). Morfológicamente, los senos fronto–nasales muestran un mayor grado de desarrollo y neumatización (Zurita et al., 2009a)
Orden Tardigrada Latham y Davis, 1795
Comentarios. Los Tardigrada, Phyllophaga o Folivora (ver Delsuc et al., 2001) representan uno de los grupos de mamíferos característicos del Cenozoico de América del Sur, pero que también se encuentra representado en América Central y América del Norte (Mathew y Paula Couto, 1959; McDonald, 2006). Comprenden dos grandes linajes: Megatherioidea y Mylodontoidea, los cuales fueron notablemente diversos durante el Neógeno (Mioceno–Plioceno; ver Scillato–Yané y Carlini, 1998b; Brandoni, 2009). Los Megatherioidea tradicionalmente comprenden tres familias: Megatheriidae Owen, 1842, Megalonychidae Ameghino, 1889 y Nothrotheriidae Gaudin, 2004. El único Megalonychidae que se conoce en Argentina durante el Ensenadense es Megalonychops carlesi Kraglievich, 1930 (MLP 95); sin embargo, su procedencia estratigráfica es dudosa, por lo cual no será incluido en el registro considerado. Asimismo, se conoce un Megalonychidae indet. (MLP 63–X–18–1), procedente del Ensenadense del norte de Mar del Plata; sin embargo, no fue hallado en esa institución (MLP), por lo cual no se pudo confirmar si se trata de M. carlesi (o de algún otro Megalonychidae). De esta forma, no se tiene registro certero de la presencia de esta familia en el Ensenadense.
Por otro lado, según las referencias bibliográficas (Carlini y Scillato–Yané, 1999; Cione et al., 1999) los Nothrotheriinae se hallan representados en el Ensenadense por Nothropus carcaranensis (procedente de Santa Fé) y Diheterocnus holmbergi Kraglievich, 1930. Este último autor describe y figura un resto asignado a Diheterocnus holmbergi procedente "al parecer del Pampeano inferior de la provincia de Buenos Aires" (Kraglievich, 1930: 16); sin embargo, este espécimen no ha sido hallado en las colecciones del MLP; el carácter dudoso de su procedencia estratigráfica pone en duda su presencia en el Ensenadense, por lo cual no será incluido en el registro de Xenarthra.
Familia Megatheriidae Owen, 1842
Megatherium Cuvier, 1796
Materiales y procedencia. El registro ensenadense de Megatherium es llamativamente escaso (en relación al Bonaerense–Lujanense) y corresponde en su mayoría a elementos fragmentarios procedentes de las "toscas del Río de La Plata" (MACN 855,1838, 1839, 2176, 2177,2899, 8843, 10451, 10654 y 10858).
Descripción y comentarios. Las especies incluidas en Megatherium poseen tamaño mediano a grande, molariformes prismáticos de lados subparalelos y sección cuadrangular; mediano a alto grado de hipsodoncia (ver Brandoni et al., 2008); margen anterior del proceso coronoide anterior al molariforme 4, extremos posterior de la sínfisis mandibular es posterior al molariforme 1; premaxilares alargados y en forma de "Y", porción predentaria del maxilar corta; facetas de articulación atlas–axis separadas; miembro anterior con dos a cinco dígitos funcionales, los dígitos dos a cuatro poseen garras; epífisis distal de la tibia comprimida anteroposteriormente; astrágalo con la faceta discoide y ectal separadas (en vista lateral) (ver además Pujos y Salas, 2004).
Megatherium gallardoi Ameghino y Kraglievich, 1921
Materiales y procedencia. El ejemplar MACN 5002 constituye el único espécimen claramente referible a esta especie y fue hallado en una excavación en la Ciudad Autónoma de Buenos Aires (34°33'23.53''S y 58°25'46.66''O).
Descripción y comentarios. El ejemplar MACN 5002 (tipo de M. gallardoi) fue descripto originalmente por Ameghino y Kraglievich (1921) y posteriormente ampliada su caracterización por Brandoni et al. (2008). Cabe destacar que dentro de Megatherium se han incluido numerosas especies cuya validez ha sido y sigue siendo discutida (ver Brandoni et al., 2008). En este sentido, varios autores consideran que la única especie de Megatheriinae válida para el Pleistoceno de la región Pampeana es M. americanum (De Iuliis 1996, Pujos y Salas, 2004; Pujos, 2006). No obstante, otros autores indican que la diversidad fue mayor (Carlini y Scillato–Yané, 1999; Cione et al., 1999; Saint–André y De Iuliis, 2001; Carlini et al., 2006) e incluyen a Megatherium gallardoi Ameghino y Kraglievich, 1921, Megatherium tarijense Gervais y Ameghino, 1880 y Megatherium silenum (Ameghino, 1898) en el registro de megaterinos del Ensenadense de la región mencionada. Según Brandoni et al. (2008), Megatherium silenum es un nomen dubium ya que el material tipo (MLP 2–61) corresponde a un ejemplar juvenil y Megatherium tarijense está restringido al Valle de Tarija, Bolivia.
Familia Mylodontidae Gill, 1872
Subfamilia Mylodontinae Gill, 1872
Comentarios. Los milodóntidos Mylodontinae son el grupo menos estudiado de la familia, con excepción de los trabajos de Kraglievich (1930), Cabrera (1936) y Esteban (1996).
Glossotherium Owen, 1839
Materiales y procedencia. Las "toscas del Río de La Plata" (IFG 72, 103), Miramar (MACN 6856) y Necochea (MLP 40–X–29–2). Los ejemplares MACN 5235 y MACN 5980 procedente de "las toscas del Río de La Plata" y de Miramar respectivamente, no fueron hallados en la colección de esa institución.
Comentarios. Esteban (1996) considera a todos los ejemplares pleistocénicos incluidos en su estudio como pertenecientes a Glossotherium robustum (Owen, 1842); no obstante, Carlini y Scillato–Yané (1999) consideran a esta especie como Lujanense y reconocen que los ejemplares ensenadenses podrían corresponder a una posible nueva especie, aún no determinada.
Lestodon Gervais, 1855
Materiales y procedencia. Este taxón fue hallado en numerosas localidades, sin embargo el estado de conocimiento actual no permite la asignación específica de todos los ejemplares. En este sentido se lo ha hallado en el norte de Mar del Plata (MMP 47 S, 235 S, MACN 5214, 14117, MLP 62–VII–27–98 y, salvo algunas excepciones que se detallan más adelante, MACN 10699 a 10835) y Punta Hermengo (MACN 2324, 8639, 10628, 17252, MLP 52–X–2–47, 52–X–4–11, 52–X–4–12).
Descripción y comentarios. Pascual et al. ( 1966) y Esteban (1996) definen Lestodon con base en características craneales. En este sentido, el cráneo de Lestodon presenta la sutura naso–frontal en forma de "U", a excepción de lo que sucede en L. armatus Gervais, 1855 donde es en forma de "V". Presenta una constricción prezigomática, el techo del hocico está formado casi exclusivamente por los nasales, posee un largo diastema, el primer molariforme superior está bien desarrollado y es de sección triangular.
Lestodon armatus Gervais, 1855
Materiales y procedencia. Según Esteban (1996), su registro comprende las localidades de las "toscas del Río de La Plata" (MACN 8846), Punta Piedras (MACN 8684), Miramar (MACN 2323, 10048) y norte de Mar del Plata (MACN 2323, 10048, 10826, 10827, 10828, 10829, 10830, 10833 y 10834).
Descripción y comentarios. Pleistoceno de Argentina, Uruguay, Paraguay, Bolivia y Brasil. Para el Ensenadense sólo se conoce L. armatus (Ameghino, 1889; Rusconi, 1931; Carlini y Scillato–Yané, 1999). Lestodon armatus es la especie tipo de género. De acuerdo con Esteban (1996), esta especie se caracteriza por presentar el borde anterior y posterior de los nasales del mismo ancho. El caniniforme superior muy proyectado hacia adelante, si se lo compara con la otra especie L. australis.
Mylodon Owen, 1839
Materiales y procedencia. Los escasos restos atribuibles a Mylodon sp. provienen de las "toscas del Río de La Plata" (MACN 854, ejemplar juvenil), ciudad de La Plata (MLP 96–III–10–6) y de Miramar (MACN 5209). Si bien Carlini y Scillato–Yané (1999) mencionan la presencia de una nueva especie exclusiva de este piso, nunca fue formalmente nominada.
Descripción y comentarios. Según Esteban (1996), Mylodon se caracteriza por presentar a nivel del premaxilar una prolongación dorsal para formar un arco prenasal que se une a los nasales. Además los nasales presentan una marcada convexidad. A nivel de la serie dentaria, su fórmula dentaria es 4/4. De acuerdo con la revisión realizada por Esteban (1996), este género sólo está representado por M. darwini Owen, 1839.
Subfamilia Scelidotheriinae Ameghino, 1904
Comentarios. Comprende una subfamilia monofilética, sustentada por más de 20 sinapomorfías (Gaudin, 2004). Existe gran controversia entre los autores en cuanto a los géneros y especies válidas, que ha complicado el panorama taxonómico del grupo (ver Pujos, 2000; McDonald y Perea, 2002; Miño–Boilini y Carlini, 2009; Miño–Boilini et al., 2009). Se reconocen, con mayor o menor grado de aceptación, tres géneros pleistocénicos: Scelidotherium Owen, 1839, Scelidodon Ameghino, 1881 y Catonyx Ameghino, 1891 (Pujos, 2000, Guérin y Faure, 2004). No obstante, McDonald (1987) y McDonald y Perea (2002) consideran que Scelidodon es un sinónimo junior de Scelidotherium.
En este trabajo se seguirá el ordenamiento propuesto por Kraglievich (1923), Pujos (2000), Miño–Boilini y Carlini (2009), con lo cual Scelidodon y Catonyx deben ser considerados géneros diferentes, estando sólo el primero registrado en la Argentina, Bolivia, Uruguay, Chile, Perú y Ecuador (McDonald, 1987; Pujos, 2000; Miño–Boilini y Carlini, 2009); mientras que Catonyx debe ser considerado endémico de Brasil (Dantas y Zucon, 2007; Cartelle et al., 2009; Miño–Boilini y Carlini, 2009).
Scelidotherium Owen, 1839
Comentarios. Los primeros restos atribuidos a Scelidotherium fueron colectados por Darwin en Punta Alta (provincia de Buenos Aires) y referidos a Scelidotherium leptocephalum Owen, 1839. Lydekker (1886) erigió Scelidotherium bravardi; el material tipo procede de las "toscas del Río de La Plata", provincia de Buenos (Argentina).
Para el ensenadense de la región pampeana, Carlini y Scillato–Yané (1999) citan una nueva especie, caracterizada por talla pequeña; no obstante y de acuerdo con Miño–Boilini y Carlini (2009) hasta el momento la única especie registrada para el ensenadense es S. bravardi. La otra especie citada para la región pampeana es Scelidotherium leptocephalum proveniente del Bonaerense–Lujanense de la provincia de Buenos Aires (Miño–Boilini y Carlini, 2009).
Scelidotherium bravardi Lydekker, 1886
Materiales y procedencia. Las "toscas del Río de La Plata" (más de 160 ejemplares, ver Soibelzon et al., 2008b)
Descripción y comentarios. Scelidotherium bravardi se registra en el Ensenadense de Buenos Aires y en el Pleistoceno de San Luis (Miño–Boilini y Carlini, 2009). Lamentablemente la mayoría de los especímenes procedentes de las "toscas del Río de La Plata" corresponden a elementos aislados del post–cráneo (e.g., tibias, calcáneos, metacarpianos) y deben ser referidos a Scelidotheriinae indet. hasta contar con más evidencia disponible. Desde un punto de vista anatómico, S. bravardi posee una talla menor que S. leptocephalum (Miño–Boilini y Carlini, 2009), a su vez el cráneo de S. bravardi es relativamente más angosto con la cresta fronto–parietal más estrecha y posee el occipital menos arqueado que el de S. leptocephalum; la mandíbula de S. bravardi posee la sínfisis de mayor longitud y es más angosta y fuertemente inclinada hacia arriba (Lydekker, 1886) que la de S. leptocephalum. McDonald (1987) y McDonald y Perea (2002) consideran que Scelidotherium bravardi es un sinónimo junior de Scelidotherium leptocephalum; sin embargo Miño–Boilini y Carlini (2009) no concuerdan con esta propuesta.
Scelidodon Ameghino, 1881
Materiales y procedencia. Las "toscas del Río de La Plata" (MACN 994 y 995) y alrededores de la ciudad de Buenos Aires (MACN 10159).
Descripción y comentarios. Ameghino (1881) erigió Scelidodon con base en un maxilar que asignó a su nueva especie Scelidodon copei (MACN A–1158, ejemplar tipo). Según Carlini y Scillato–Yané (1999) esta especie estaría registrada en el Ensenadense de la provincia de Buenos Aires. Ameghino (1889) y Kraglievich (1923) asignan los ejemplares MACN 994 y 995 a Scelidodon capellini (Gervais y Ameghino, 1880), sin embargo, ambos autores coinciden en la similitud morfológica de esta especie con Scelidodon tarijensis (Gervais y Ameghino, 1880), no obstante el carácter fragmentario de ambos ejemplares no permite asignarlos a nivel específico. Según Carlini y Scillato–Yané (1999) y Cione y Tonni (2005), S. capellini es exclusiva del Ensenadense, sin embargo nunca se mencionó sobre cuáles especímenes se basó esta asignación (ver Miño–Boilini y Carlini, 2009). De acuerdo con Pujos (2000) y McDonald y Perea (2002) esta especie es un sinónimo posterior de Scelidodon tarijensis (Gervais y Ameghinio, 1880).
Scelidodon cf. tarijensis
Materiales y procedencia. MMP 3989 (porción posterior de cráneo) procedente de los alrededores de la ciudad de Mar del Plata.
Descripción y comentarios. Scelidodon tarijensis (=Catonyx tarijensis de McDonald y Perea, 2002) (Gervais y Ameghino, 1880) se caracteriza por presentar a nivel del cráneo la cresta sagital muy marcada y la sutura que separa el escamoso del parietal y del frontal de orientación sub–horizontal; a nivel mandibular el molariforme 4 posee un lóbulo accesorio (ver McDonald, 1987, McDonald y Perea, 2002, Miño–Boilini y Carlini, 2009). Esta especie fue citada para el Ensenadense de Bolivia por Pujos (2000) y McDonald y Perea (2002). Esta especie fue mencionada por Miño–Boilini y Carlini (2009) en sedimentos del Ensenadense y Bonaerense de la provincia de Buenos Aires.
RESULTADOS
Con la información disponible se elaboró un listado de los xenartros presentes en el Piso Ensenadense de la región pampeana (ver Tabla 1), tomando como base los últimos listados de Xenarthra ensenadenses (e.g., Carlini y Scillato–Yané, 1999; Cione y Tonni, 2005).

En este trabajo se reconocen tanto géneros y especies nuevas (i.e., Chaetophractus vellerosus, Eutatus pascuali, Tolypeutes matacus, Panochthus subintermedius), como otras exclusivas de este piso (i.e., Neosclerocalyptus ornatus, Neosclerocalyptus pseudornatus, Neuryurus rudis, Panochthus intermedius, Panochthus subinter–medius, Glyptodon munizi y Megatherium gallardoi). Las especies Chaetophractus vellerosus, Zaedyus pichiy, Propraopus grandis, Tolypeutes matacus, Pampatherium typum, Doedicurus, Mylodon, Scelidotherium bravardi y Scelidodon tarijensis se distribuyen desde el Ensenadense hasta Pisos más modernos.
Ciertos taxones pobremente definidos o con dudas en cuanto a su procedencia estratigráfica y/o geográfica se excluyeron del listado de fauna ensenadense. De esta manera, " Tolypeutes pampaeus", "Sclerocalyptus perfectus", "Sclerocalyptus scrobiculatus", Neothoracophorus elevatus, Lomaphorus compressus, Lomaphorus imperfectus, Daedicuroides eguiae, Doedicurus kokenianus, Plaxhaplous ensenadensis, Glyptodon principalis, Glyptodon gemmatum, Glyptodon laevis, Glyptodon perforatus, Diheterochnus holmbergi, Nothropus carcaranensis, Megatherium sile–num, Megatherium tarijense, Megalonychops carlesi y Scelidodon capellini son descartadas del elenco de xenartros del ensenadense, por diferentes motivos expuestos anteriormente; mientras que la presencia de Neothoracophorus elevatus, Lomaphorus compressus, Lomaphorus imperfectus, Daedicuroides eguiae, Plaxhaplous ensenadensis, Glyptodon principalis, Glyptodon gemmatum, Glyptodon laevis, Glyptodon perforatus no pudo ser corroborada.
CONCLUSIONES
Los resultados obtenidos en este trabajo evidencian que durante el Ensenadense se registran 18 géneros y 19 especies de Xenarthra, siete de ellas exclusivas de este Piso. Algunos de ellos incluyen más de una especie (i.e., Chaetophractus, Panochthus y Neosclerocalyptus), mientras que la gran mayoría son monoespecíficos. Asimismo, en algunos casos, la escasez de restos, su carácter fragmentario y/o la ausencia de estudios anatómicos actualizados, impidió la asignación específica de alguno de los especimenes ensenadenses (i.e., Doedicurus, Lomaphorus). De esta manera, se observa que los listados precedentes (e.g., Carlini y Scillato–Yané, 1999) sobredimensionaron el elenco pampeano, reconociendo unas 36 especies de Xenarthra ensenadenses.
Desde una perspectiva paleoecológica, es interesante mencionar que durante el Ensenadense se produce una gran diversificación faunística y, particularmente entre los Xenarthra, se encuentran las formas de mayor tamaño del Pleistoceno (Scillato–Yané y Carlini, 1998a; Soibelzon, 2008). Zurita et al. (2010) hallaron y describieron una serie de osteodermos cónicos, ubicados en hileras sobre los flancos de la coraza de Glyptodon munizi (MLP 07–V–3–2) y de Glyptodon reticulatus Owen, 1845 (dos ejemplares provenientes del Lujanense de la provincia de Buenos Aires). Dichos osteodermos sirvieron de protección y defensa del cuello y del vientre, las zonas más desprotegidas en estos grandes mamíferos acorazados. La presencia de estas estructuras accesorias de protección (sin registro en las especies preensenadenses) y la alta masa corporal registrada en numerosos taxones ensenadenses (e.g., Megatherium gallardoi), permite conjeturar que estas características se podrían haber desarrollado en los mamíferos autóctonos de América del Sur en respuesta al arribo de los Carnívora placentarios (desde América del Norte), durante el "Gran Intercambio Biótico Americano" (más detalles sobre esto en Zurita et al., 2010). Por último, es importante mencionar que de los Glyptodontidae registrados en el Pleistoceno de la región pampeana, Glyptodon es claramente el más frecuente, tanto en las secuencias ensenadenses como pos–ensenadenses, en concordancia con lo observado por Zurita et al. (2009b) para el Valle de Tarija (Bolivia).
Desde una perspectiva paleoclimática, durante el último millón de años se han observado al menos 15 ciclos glaciales–interglaciales, especialmente a partir de la Gran Glaciación Patagónica (GPG, ca. 1168–1016 Ma; ver Rabassa et al., 2005; Soibelzon y Tonni, 2009). En este sentido, la mayoría de las asociaciones faunísticas ensenadenses reflejan condiciones áridas y más frías que las actuales, con el desarrollo de ambientes abiertos o semiabiertos (e.g., Zaedyus pichiy, Tolypeutes matacus, Chaetophractus vellerosus, Eutatus sp., Glyptodontidae y Mylodontidae; ver Scillato–Yané et al., 1995); a la vez que ciertos taxones como Tapirus indican la ocurrencia de pulsos cálidos (ver Soibelzon y Tonni, 2009). Es interesante remarcar que entre los Xenarthra, hay un claro ejemplo de la presión que ejercieron estos cambios climáticos sobre la fauna. De esta manera, en Neosclerocalyptus se observa una progresiva neumatización y expansión lateral de los senos fronto–nasales. La aparición de este carácter coincide temporalmente con la GPG y su desarrollo aumenta desde las especies ensenadenses a las lujanenses, claramente en respuesta a temperaturas gradualmente más frías.
Es importante remarcar que los Xenarthra ensenadenses exhumados en la región pampeana son sin dudas los más diversos de América del Sur, tanto morfológica como taxonómicamente. Fuera de Argentina, los yacimientos con fauna ensenadense son particularmente escasos y se los conoce únicamente en Uruguay y Bolivia (ver Marshall et al., 1984). Es importante remarcar que la asignación al Ensenadense pampeano de los fósiles provenientes de Uruguay (Formación Raigón) resulta dudosa, ya que el único taxón exhumado allí es Scelidodon tarijensis (ver Ubilla et al., 2008). Nuestros estudios indican que este taxón se distribuye desde el Ensenadense hasta pisos más recientes. Por otro lado, numerosos investigadores indicaron una antigüedad ensenadense para los fósiles provenientes del Valle de Tarija (Bolivia; ver Tonni et al., 2009 y la bibliografía allí citada). Bioestratigráficamente sólo tres especies allí presentes (i.e., Glyptodon munizi, Arctotherium angustidens y Hippidion devillei) son exclusivos del Ensenadense pampeano, mientras que el grueso de la fauna es bioestratigráficamente referible al Bonaerense–Lujanense (Pleistoceno medio–Holoceno temprano; ver Tonni et al., 2009). Por lo tanto, si toda la secuencia de Tarija se correlaciona con la Edad Lujanense pampeana, entonces la supervivencia de ciertos taxones exclusivos del Ensenadense pampeano podría deberse a un efecto de existencia continuada y supervivencia local.
AGRADECIMIENTOS
Este manuscrito es parte de los trabajos de Tesis Doctoral de los autores. En este sentido, queremos expresar nuestro más sincero agradecimiento a E. P. Tonni, G. J. Scillato–Yané y A. A. Carlini, quienes nos han dirigido a lo largo de estos años de trabajo. Asimismo, nuestro más sincero agradecimiento a todas aquellas personas que aportaron información e ideas: M. Bond, D. Brandoni, M. M. Cenizo, M. R. Ciancio, A. L. Cione, M. de los Reyes, M. Zamorano. A los encargados de las diferentes colecciones visitadas: A. Kramarz y Laura Cruz (MACN), S. Bargo, M. Reguero y L. H. Pomi (MLP), E. Cerdeño (Col. Rusconi), A. Dondas (MMP), J. E. Powell (FML), S. Cornero y F. R. Solomita (IFG), J. L. Aguilar y J. I. Verdón (MSP), C. de Muizon, C. Sagne y P. Tassy (MNHN, París). A los revisores Federico Agnolín y Victor Manuel Bravo Cuevas por sus comentarios y sugerencias. A Francisco J. Vega (Editor Científico) y Ma. Teresa Orozco–Esquivel (Editora Técnica) de la RMCG por su apoyo y sugerencias durante las etapas de envío y publicación del manuscrito. Este trabajo fue parcialmente subsidiado por la ANPCyT, CONICET, CIC.
REFERENCIAS
Alberdi, M.T., Leone, G., Tonni, E.P. (eds.), 1995, Evolución biológica y climática de la región Pampeana durante los últimos cinco millones de años. Un ensayo de correlación con el Mediterráneo occidental: Madrid, Museo de Ciencias Naturales, Consejo de Investigaciones, Monografías CSIC, 12, 423 pp. [ Links ]
Ameghino, F., 1881, La antigüedad del hombre en el Plata: París y Buenos Aires, Masson–Igon, 557pp. [ Links ]
Ameghino, F., 1883, Sobre la necesidad de borrar el género Schistopleurum y sobre la clasificación y sinonimia de los Gliptodontes en general: Boletín de la Academia Nacional de Ciencias, 5, 1–34. [ Links ]
Ameghino, F., 1889, Contribución al conocimiento de los mamíferos fósiles de la República Argentina: Actas de la Academia Nacional de Ciencias de Córdoba, 6, 1–1027. [ Links ]
Ameghino, F., 1891, Mamíferos y aves fósiles argentinas. Especies nuevas, adiciones y correcciones: Revista Argentina de Historia Natural, 1(4), 240–259. [ Links ]
Ameghino, F., 1895, Sur les édentés fossiles de l' Argentine (Examen critique, révision et correction de l* ouvrage de M. R. Lydekker "The extinct Edentates of Argentina"): Revista del Jardín Zoológico de Buenos Aires, 3, 97–192. [ Links ]
Ameghino, F., 1898, Sinopsis geológico–paleontológica, en II Censo de la República Argentina: Buenos Aires, Argentina, 111–255. [ Links ]
Ameghino, F., 1904, Nuevas especies de mamíferos cretáceos y terciarios de la República Argentina (continuación): Anales de la Sociedad Científica Argentina, 58, 1–188. [ Links ]
Ameghino, C., Kraglievich, L., 1921, Descripción del Megatherium gallardoi C. Ameg. descubierto en el Pampeano Inferior de la Ciudad de Buenos Aires: Anales del Museo Nacional de Historia Natural de Buenos Aires, 31, 135–156. [ Links ]
Bostelman, E.T., Rinderknecht, A., Lecuona, G., 2008, Los Eutatini (Mammalia, Dasypodidae) del Uruguay, en III Congreso Latinoamericano de Paleontología de Vertebrados, Neuquén, Argentina. resumen 28. [ Links ]
Bordas, A.F., 1933, Notas sobre los Eutatinae nueva subfamilia extinguida de Dasypodidae: Anales del Museo Argentino de Ciencias Naturales, 37, 583–614. [ Links ]
Brandoni, D., 2009, Descripción anatómica del fémur de Protomegalonyx chasicoensis S cillato–Yané (Mammalia, Xenarthra, Megalonychidae) procedente la Formación Arroyo Chasicó (Mioceno tardío), de la provincia de Buenos Aires, Argentina: Ameghiniana, 46(3), 513–521. [ Links ]
Brandoni, D., Soibelzon, E., Scarano, A., 2008, On the status of Megatherium gallardoi Ameghino y Kraglievich (Xenarthra, Tardigrada) and its biostratigraphic implications: Geodiversitas, 30(4), 793–804. [ Links ]
Burmeister, H., 1866, Einige Bemerkungen über die im Museum zu Buenos Aires befindlichen Glyptodonarte: Zeitschrift für die gesammten Naturwissenschaften, 28, 138–142. [ Links ]
Burmeister, H., 1874, Monografía de los Glyptodontes en el Museo Público de Buenos Aires: Anales del Museo Público de Buenos Aires, 2, 412 pp. [ Links ]
Burmeister, H., 1879, Description physique de la Repúblique Argentine d aprés des observations personelles et ètrangéres: animaux vertébrés, 1, Mammifères vivants et éteints, 1–155. [ Links ]
Cabrera, A.L., 1936, Las especies del género Glossotherium: Notas del Museo de La Plata, 1, 193–206. [ Links ]
Cabrera, A.L., Willink, A., 1980, Biogeografía de América Latina: Washington, Organización de Estados Americanos, serie Biología, 13, 122 pp. [ Links ]
Carlini, A.A., Scillato–Yané, G.J., 1999, Evolution of Quaternary Xenarthrans (Mammalia) of Argentina, in Tonni, E.P, Cione A.L. (eds.), Quaternary of South America and Antarctic Peninsula: Rotterdam, Holanda, A.A. Balkema, 12, 149–175. [ Links ]
Carlini, A.A., Tonni, E.P., 2000, Mamíferos Fósiles del Paraguay: Buenos Aires, Cooperación Técnica Paraguayo–Alemana, Proyecto Sistema Ambiental del Chaco–Proyecto Sistema Ambiental Región Oriental, 108 pp. [ Links ]
Carlini, A.A., Vizcaino, S.F., 1987, Chaetophractus vellerosus (Gray, 1865) (Mammalia, Dasypodidae) in Buenos Aires Province, Argentina. Its disjunctive distribution: possible cause: Studies on Neotropical Fauna and Environment, 22 (1), 53–56. [ Links ]
Carlini, A.A., Brandoni, D., Sánchez, R., 2006, First megatheriines (Xenarthra, Phyllophaga, Megatheriidae) from the Urumaco (Late Miocene) and Codore (Pliocene) Formations, estado Falcón, Venezuela: Journal of Systematic Paleontology, 4(3), 269–278. [ Links ]
Carlini, A.A., Zurita, A.E., Aguilera, O.A., 2008a, North American Glyptodontines (Xenarthra, Mammalia) in the upper Pleistocene of northern South America: Palaeontologische Zeitschrift, 82, 125–138. [ Links ]
Carlini, A.A., Zurita, A.E., Scillato–Yané, G.J., Sánchez, R., Aguilera, O.A., 2008b, New glyptodont from the Codore Formation (Pliocene), Falcón State, Venezuela, its relationship with the Asterostemma problem, and the paleobiogeography of the Glyptodontina: Palaeontologische Zeitschrift, 82, 139–152. [ Links ]
Cartelle, C., Bohorquez, G.A., 1985, Pampatheriumpaulacoutoi, uma nova espécie de tatu gigante da Bahia, Brasil (Edentata, Dasypodidae): Revista Brasileira de Zoologia, 2(4), 229–254. [ Links ]
Cartelle, C., De Iuliis, G., Lopes–Ferreira, R., 2009, Systematic revision of tropical Brazilian Scelidotheriine sloths (Xenarthra, Mylodontoidea): Journal of Vertebrate Paleontology, 29(2), 555–566. [ Links ]
Castellanos, A., 1933, Las arenas del antiguo Paraná y la perforación de Villa Ballester (F.C.C.A.) ¿Quid Novi?: Revista de las Asociaciones de Exalumnas y Padres de la Escuela Normal, 2(5), 1–11. [ Links ]
Castellanos, A., 1940, A propósito de los géneros Plohophorus, Nopachthus y Panochthus: Publicaciones del Instituto de Fisiografía y Geología, 1(6), 1–279. [ Links ]
Castellanos, A., 1951, Acotaciones al género Neothoracophorus Ameghino: Revista de la Asociación Geológica Argentina, 6(1), 63–82. [ Links ]
Cattoi, N., 1966, Orden Edentata, en Borrelo, A.V. (ed.), Paleontografía Bonaerense. Fascículo IV: Vertebrata: Buenos Aires, Comisión de Investigaciones Científicas, 59–99. [ Links ]
Cione, A.L., Tonni, E.P., 1995, Bioestratigrafía y cronología del Cenozoico de la región Pampeana, en Alberdi, M.T., Leone, G., Tonni, E.P. (eds.), Evolución biológica y climática de la región Pampeana durante los últimos cinco millones de años. Un ensayo de correlación con el Mediterráneo occidental: Madrid, Museo Nacional de Ciencias Naturales, Consejo Superior de Investigaciones Científicas, 47–74. [ Links ]
Cione, A.L., Tonni, E.P., 1999, Biostratigraphy and chronological scale of upper–most Cenozoic in the Pampean Area, Argentina, in Tonni, E.P, Cione A.L. (eds.), Quaternary of South America and Antarctic Peninsula: Rotterdam, Holanda, A.A. Balkema, 12, 23–51. [ Links ]
Cione, A.L., Tonni, E.P., 2005, Bioestratigrafía basada en mamíferos del Cenozoico Superior de la provincia de Buenos Aires, Argentina, en XVI Congreso Geológico Argentino, La Plata, Argentina, Relatorio, 183–200. [ Links ]
Cione, L.A., Tonni E.P., Bond, M., Carlini A.A., Pardiñas, U.F., Scillato–Yané, G.J., Verzi, D., Vucetich, M.G., 1999, Occurrence charts of Pleistocene mammals in the Pampean area, eastern Argentina, in Tonni, E.P, Cione A.L. (eds.), Quaternary of South America and Antarctic Peninsula: Rotterdam, Holanda, A.A. Balkema, 12, 53–59. [ Links ]
Cope, E.D., 1889, The Edentata of North America: American Naturalist, 23, 657–664. [ Links ]
Croft, D.A., Flynn, J.J., Wyss, A.R, 2007, A new basal Glytodontid and the other taxa Xenarthra of the early Miocene Chucal fauna, Northern Chile: Journal of Vertebrate Paleontology, 27(4), 781–197. [ Links ]
Cruz, L.E., Zamorano, M., Scillato–Yané, G.J., 2007, Panochthus subintermedius Castellanos (Xenarthra, Glyptodontidae): redescripción e implicancia biocronológica: Ameghiniana, 44(4), 13R. [ Links ]
Cuvier, G., 1796, Notice sur le squelette d'une très grande espèce de quadrupède inconnue jusqu'à présent, trouvé au Paraquay, et déposé au cabinet d'histoire naturelle de Madrid: Magasin encyclopédique, ou Journal des Sciences, des Lettres et des Arts, 1, 303–310. [ Links ]
Dantas, M.A.T., Zucon, M.H., 2007, Occurrence of Catonyx cuvieri (Lund, 1839) (Tardigrada, Scelidotheriinae) in late Pleistocene–Holocene of Brazil: Revista Brasileira de Paleontologia, 10, 129–232. [ Links ]
De Iuliis, G., 1996, A Systematic Review of the Megatheriinae (Mammalia: Xenarthra: Megatheriidae): Canada, University of Toronto, tesis doctoral, 781 pp. [ Links ]
Delsuc, F., Douzery, E.J.P., 2008, Recent advances and future prospects in xenarthran molecular phylogenetics, in Vizcaíno, S.F., Loughry, W.J. (eds.), The Biology of the Xenarthra: Gainesville, University Press of Florida, 11–23. [ Links ]
Delsuc, F., Douzery, E.J.P., 2009, Armadillos, anteaters, and sloths (Xenarthra), in Hegges, S.B., Kumer, S. (eds.), The Timetree of life: London, Oxford University Press, 475–478. [ Links ]
Delsuc, F., Catzeflis, F.M., Stanhope, M.J., Douzery. E.J.P., 2001, The evolution of armadillos, anteaters and sloths depicted by nuclear and mitochondrial phylogenies: implications for the status of the enigmatic fossil Eurotamandua: Proceedings of the Royal Society of London B, 268, 1605–1615. [ Links ]
Delsuc, F., Scally, M., Madsen, O., Stanhope, M.J., de Jong, W.W., Catzeflis, F.M., Springer, M.S., Douzery, E.J.P., 2002, Molecular Phylogeny of Living Xenarthrans and the Impact of Character and Taxon Sampling on the Placental Tree Rooting: Molecular Biology and Evolution, 19(10), 1656–1671. [ Links ]
Delsuc, F., Stanhope, M.J., Douzery, E.J.P., 2003, Molecular systematics of armadillos (Xenarthra, Dasypodidae): contribution of maximum likelihood and Bayesian analyses of mitochondrial and nuclear genes: Molecular Phylogenetics and Evolution, 28(2), 261–275. [ Links ]
Desmarest, 1804, Nouveau dictionnaire d'Histoire naturelle: Paris, Chez Deterville, 24 , 1–577. [ Links ]
Duarte, R.G., 1997, Gliptodontes del Pleistoceno tardío de Aguas de las Palomas, Campo de Pucará, Catamarca, Argentina. Variaciones morfológicas del caparazón de Glyptodon reticulatus Owen, 1845: Ameghiniana, 34(3), 345–355. [ Links ]
Esteban, G.I., 1996, Revisión de los Mylodontinae Cuaternarios (Edentata–Tardigrada) e Argentina, Bolivia y Uruguay. Sistemática, Filogenia, Paleobiología, Paleozoogeografía y Paleoecología, Tucumán, Argentina: Universidad Nacional de Tucumán, tesis doctoral, 235 pp. [ Links ]
Fidalgo, F., Tonni, E.P., 1983, Geología y paleontología de los sedimentos encauzados del Pleistoceno tardío y Holoceno en Punta Hermengo y Arroyo Las Brusquitas (Partido de General Alvarado y General Pueyrredón, Provincia de Buenos Aires: Ameghiniana, 20 (3–4), 281–296. [ Links ]
Fitzinger, L.J., 1871, Die natürliche Familie der Gürteltiere (Dasypodes): Sitzungsberichte Mathematisch–Naturwissenschaftliche Klasse der Akademie der Wissenschaften, Wien, 64, 209–276. [ Links ]
Frenguelli, J., 1921, Los Terrenos de la Costa Atlántica en los alrededores de Miramar (Prov. de Buenos Aires) y sus correlaciones: Boletín de la Academia Nacional de Ciencias de Córdoba, 24(3–4), 325–485. [ Links ]
Gardner, A., Wilson, D.E., Reeder, D.M. (eds), 2005, Mammal Species of the World. A Taxonomic and Geographic Reference: Baltimore, Johns Hopkins University Press, 142 pp. [ Links ]
Gaudin, T.J., 2003, Phylogeny of the Xenarthra, in Fariña, R.A., Vizcaíno, S.F., Storch, G. (eds.), Morphological Studies in Fossil and Extant Xenarthra (Mammalia): Frankfurt am Main, Seckenbergiana Biologica, 27–40. [ Links ]
Gaudin, T.J., 2004, Phylogenetic relationships among sloths (Mammalia, Xenarthra, Tardigrada): the craniodental evidence: Zoological Journal of the Linnean Society, 140, 255–305. [ Links ]
Gervais, P., 1855, Recherches sur les mammifères fossiles propres à l*Amérique méridionale: Comptes rendus des séances de l' Académie des Sciences, 40(20), 1112–1114. [ Links ]
Gervais, P., 1867, Sur une nouvelle colection d'ossementes fossiles de Mammifères recueillé par M. Fr. Seguin dans la Conféderation Argentine: Comptes rendus des séances de l' Académie des Sciences, 65, 279–282. [ Links ]
Gervais, P., 1878, Nouvelles recherches sur les mammifères fossiles propres à l*Amérique méridionale: Comptes rendus des séances de l' Académie des Sciences, , 86(22), 1359–1362. [ Links ]
Gervais, H., Ameghino, F., 1880, Los mamíferos fósiles de la América del Sur: Paris– Buenos Aires, Sabih e Igon, 225 pp. [ Links ]
Gill, T., 1842, Arrangement of the families of mammals with analytical tables: Smihsonian Miscellaneous Collections, 11(1), 1–98. [ Links ]
Guérin, C., Faure, M., 2004, Scelidodon piauiense nov. sp., nouveau Mylodontidae, Scelidotheriinae (Mammalia, Xenarthra) du Quaternaire de la region du parc national Serra da Capivara (Piauí, Brésil): Comptes Rendus Palevol, 3, 3–42. [ Links ]
Giraudo, A., 1997, El Concepto de especie. Parte I: Natura Neotropicalis, 28(2), 161–169. [ Links ]
Gray, J.E., 1821, On the natural arrangement of vertebrose animal: London Medical Repository, 15(1), 296–310. [ Links ]
Gray, J.E., 1865, Revision on the genera and species of entomophagous, Edentata, founded on examination of the specimens in British Museum: Proceedings of the Zoological Society of London, 33(1), 359–386. [ Links ]
Gray, J.E., 1869, Catalogue of carnivorous, pachydermatous and edentate Mammalia in the British Museum: London, British Museum of Natural History, 398 pp. [ Links ]
Goloboff, P.A., Catalano, S.A., Mirande, J.M., Szumik, C.A., Arias, J.S., Källersjö, M., Farris, J.S., 2009, Phylogenetic analysis of 73 060 taxa corroborates major eukaryotic groups: Cladistics, 25(3), 211–230. [ Links ]
Harlan, R., 1825, Description of a new genus of mammmiferous quadrupeds of the Order Edentata: Annals of the New York Lyceum of the Natural History, 1, part 2, 235–246. [ Links ]
Hoffstetter, R., 1958, Xenarthra, in Piveteau, J. (ed.): Paris, Traité de Paléontologie, 6(2), 535–636. [ Links ]
Huxley, T.H., 1864, On the osteology of the genus Glyptodon: Philosophical transactions in the Royal Society of London, 155, 31–70 [ Links ]
Illiger, C., 1811, Prodromus systematis mammalium et avium; additis terminis zoographicis utriusque classis, eorumque versione germanica: Berlin, C. Salfeld, 301 pp. [ Links ]
Kraglievich, L., 1923, Descripción comparada de los cráneos de Scelidodon rothi Ameghino y Scelidotherium parodi n. sp. procedentes del horizonte "chapadmalense": Anales del Museo Nacional de Historia Natural "Bernardino Rivadavia", 33, 57–103. [ Links ]
Kraglievich, L., 1930, Nuevos megalonícidos gigantescos de los géneros Megalonychops Kraglievich y Diheterocnus Kraglievich: Revista del Museo de La Plata, 32, 9–21. [ Links ]
Kraglievich, L., 1934, La antigüedad Pliocena de las faunas de Monte Hermoso y Chapadmalal, deducidas de su comparación con las que le precedieron y sucedieron: El Siglo Ilustrado, 17–133. [ Links ]
Krmpotic, C.M., 2009, "Los Eutatini (Xenarthra, Dasypodidae) del Neógeno Tardío del Cono Sur de América del Sur (Argentina Y Uruguay). Filogenia, diversidad e historia biogeográfica": La Plata, Argentina, Universidad Nacional de La Plata, tesis doctoral, 427 pp. [ Links ]
Krmpotic, C.M., Scillato–Yané, G.J., 2007, Rectificación de la procedencia estratigráfica de Eutatus seguini Gervais, 1867 (Xenarthra, Dasypodidae): Ameghiniana, 43(4), 637–638. [ Links ]
Krmpotic, C.M., Carlini, A.A., Scillato–Yané, G.J., 2009a, The species of Eutatus (Mammalia, Xenarthra): Assessment morphology and climate: Quaternary International, 210(1–2), 66–75. [ Links ]
Krmpotic, C.M., Ciancio, M.R., Barbeito, C., Mario, R.C., Carlini, A.A., 2009b, Osteoderm Morphology in Recent and Fossil Euphractinae Xenarthrans: Acta Zoologica, 90(4), 339–351. [ Links ]
Latham J., Davies, H., 1795, Faunula indica, in Forster, J.R. (ed.), Zoologia indica: Halle, Joannis Jacobi Gebaueri, 38 pp. [ Links ]
Linnaeus, C., 1758, Systema naturae per regna tria naturae, secundum classes, ordines, genera, species, cum characteribus, differentiis, synonymis, locis: Stockholm, Laurentii Salvii, Editio Decima, reformata, t. I, 824 pp. [ Links ]
Lizarraga, de, R., 1916, Descripción colonial (título original "Descripción breve de toda la tierra del Perú, Tucumán, Río de la Plata y Chile", 1605): Buenos Aires, Colección Biblioteca Argentina, 239 pp. [ Links ]
Lund, P.W., 1839, Coup da oeil sur les espéces éteintes de mammifères du Bresil, extrait de quelques mémoires presentés a la Acadamie Royal des Sciences de Copenhaghe: Annales des Sciences Naturelles (Zoologie), 2(11), 214–234. [ Links ]
Lydekker, R., 1886, Description of three species of Scelidotherium: Proceedings of the Zoological Society of London, 496–537. [ Links ]
Lydekker, R., 1887, Catalogue of the fossil Mammalia in the British Museum (Natural History): London, British Museum, 345pp. [ Links ]
Lydekker, R., 1894, Contribution to a knowledge of the fossil vertebrates of Argentina. 2. The extinct Edentates of Argentina: Anales del Museo de La Plata, (Paleontología), 3, 1–118. [ Links ]
Marshall, L.G., Berta, A., Hoffstetter, R., Pascual, R., Reig, O.A., Bombin, M., Mones, A., 1984, Mammals and stratigraphy: geochronology of the continental mammal–bearing quaternary of South America: Palaeovertebrata, 1–76. [ Links ]
Mathew, W.D., Couto, C de P., 1959, The Cuban Edentates: Bulletin of the American Museum of Natural History, 117, 1–56. [ Links ]
Mayr, E., 1996, What is species, and what is not?: Philosophy of Science, 63, 262–277. [ Links ]
McDonald, H.G., 1987, A Systematic Review of the Plio–Pleistocene Scelidotherinae Group Sloth (Mammalia: Xenarthra: Mylodontidae): Toronto, Canada, University of Toronto, tesis doctoral, 478 pp. [ Links ]
McDonald, H.G., 2006, Sexual dimorphism in the skull of Harlan's ground sloth. Contributions in Science: Natural History Museum of Los Angeles County, 510, 1–9. [ Links ]
McDonald, H.G., Naples, V.L., 2007, Xenarthra, in Janis, C.M., Gunnell, G.F., Uhen, M.D. (eds.), Evolution of Tertiary Mammals of North America: Cambridge University Press, 147–160. [ Links ]
McDonald, H.G., Perea, D., 2002, The large scelidothere Catonyx tarijensis (Xenarthra, Mylodontidae) from the Pleistocene of Uruguay: Journal of Vertebrate Paleontology, 22(3), 677–683. [ Links ]
McKenna, M.C., Bell, S.K., 1997, Classification of Mammals above the Species Level: New York, Columbia University Press, 631, pp. [ Links ]
Miño–Boilini, A.R., Carlini, A.A., 2009, The Scelidotheriinae Ameghino, 1904 (Phyllophaga, Xenarthra) from the Ensenadan–Lujanian Stage/Ages (Early Pleistocene to Early–Middle Pleistocene–Early Holocene) of Argentina: Quaternary International, 210(1–2), 93–101. [ Links ]
Miño–Boilini, A.R., Carlini, A.A., Chiesa, J.O., Lucero, N.P., Zurita, A.E., 2009, First record of Scelidodon chiliense (Lydekker) (Phyllophaga, Scelidotheriinae) from the Lujanian stage (late Pleistocene– early Holocene) of Argentina: Neues Jahrbuch für Geologie und Palãontologie–Abhandlungen, 253(2–3), 373–381. [ Links ]
Mones, A., 1986, Paleovertebrata Sudamericana. Catálogo sistemático de los vertebrados fósiles de América del Sur. Parte I. Lista preliminar y Bibliografía: Courier Forschungsinstitut Senckenberg, 82, 1–625 [ Links ]
Mones, A., Francis, J.C., 1973. Lista de vertebrados fósiles del Uruguay, II.: Comunicaciones Paleontologicas del Museo de Historia Natural de Montevideo, 4(1), 39–97. [ Links ]
Nodot, L., 1857, Description d un nouveaux genre d.édenté fossile renfermant plusieurs espèces voisines du Glyptodon, etc. Mémoires de L.Académie Impérial des Sciences, Arts et Belles–Lettres de Dijon, 2(5), 1–170. [ Links ]
Nowak, R.M., 1999, Order Xenarthra, in Johns Hopkins University Press. (ed.), Walker's Mammals of the world: Baltimore, Hopkins Fulfillment Service, 149–168. [ Links ]
Osgood, W.H., 1919, Names of some South American mammals: Journal of Mammalogy, 1, 33–36. [ Links ]
Owen, R., 1839, Description of a tooth and part of the skeleton of the Glyptodon, a large quadruped of the edentate order, to which belongs the tessellated bony armour figured by Mr. CLIFT in his memoir on the remains of the Megatherium, brought to England by Sir Woodbine PARISH, F.G.S.: Proceedings of the Geological Society of London, 3, 108–113. [ Links ]
Owen, R., 1842, Description of the skeleton of an extinct gigantic sloth, Mylodon robustus, Owen, with observations on the osteology, natural affinities, and probable habits of the Megatherioids quadrupeds in general: Londres, R and J. Taylor, 176 pp. [ Links ]
Owen, R., 1845, Descriptive and illustrated catalogue ofthe fossil organic remains of Mammalia and Aves contained in the Museum of the Royal College of Surgeons of London: England, Richard and John E. Taylor, 391 pp. [ Links ]
Parera, A., 2002, Los mamíferos de la Argentina y la región austral de Sudamérica: Buenos Aires, El Ateneo, 453 pp. [ Links ]
Pascual, R., Ortega Hinojosa, E.J., Gondar, D.G., Tonni E.P., 1966, Vertebrata, en Borrello, A.V. (ed.), Paleontografía Bonaerense. Vertebrata I: Buenos Aires, Comisión de Investigaciones Científicas de la provincia de Buenos Aires, 202 pp. [ Links ]
Paula Couto, C. de, 1954, Sôbre um gliptodonte do Uruguai e um tatú fóssil do Brasil, Divisão de Geologia e Mineralogia, Notas preliminares e Estudos, 80, 1–10. [ Links ]
Paula Couto, C. de, 1957, Sôbre um gliptodonte do Brasil: Boletim Divisão de Geología e Mineralogía, 165, 1–37. [ Links ]
Paula Couto, C. de 1979, Tratado de Paleomastozoología: Rio de Janeiro, Academia Brasileira de Ciencias, 590 pp. [ Links ]
Perea, D., 1998, Xenarthra fósiles del Uruguay: Distribución estratigráfica; caracterización osteológica y sistemática de algunos Tardigrada: Montevideo, Uruguay, Universidad de la República, Facultad de Ciencias, Departamento de Paleontología, Tesis doctoral, 107 pp. [ Links ]
Pérez, L.M., Scillato–Yané, G.J., Vizcaíno, S.F., 2000, Estudio morfofuncional del aparato hiodeo de Glyptodon cf. clavipes (Cingulata: Glyptodontidae): Ameghiniana, 37(3), 293–299. [ Links ]
Poljak, S., Escobar, J., Deferrari, G., Lizarralde, M., 2007, Un nuevo mamífero introducido en la Tierra del Fuego: el "peludo" Chaetophractus villosus (Mammalia, Dasypodidae) en Isla Grande: Revista Chilena de Historia Natural, 80, 285–294. [ Links ]
Pujos, F., 2000, Scelidodon chiliensis (Xenarthra, Mammalia) du Pléistocène terminal de "Pampa de los Fósiles": Quaternaire, 11(3–4), 197–206. [ Links ]
Pujos, F., 2006, Megatherium celendinense sp. nov. from the Pleistocene of Peruvian Andes and the megatheriine phylogenetic relationship: Palaeontology, 49(1), 285–306. [ Links ]
Pujos, F., Salas, R., 2004, A new species of Megatherium (Mammalia: Xenarthra: Megatheriidae) from the Pleistocene of Sacaco and Tres Ventanas, Peru: Palaeontology, 47(3), 579–604. [ Links ]
Rabassa, J., Coronato, A.M., Salemme, M., 2005, Chronology of the Late Cenozoic Patagonian glaciations and their correlation with biostratigraphic units of the Pampean region (Argentina): Journal of South American Earth Sciences, 20(1–2), 81–103. [ Links ]
Rinderknecht, A., 2000a, Estudios sobre la familia Glyptdontidae Gray, 1869. II. Variación morfológica en la coraza de Glyptodon sp. juvenil (Mammalia, Cingulata): Revista de la Sociedad Uruguaya de Geología, 3(7), 32–35. [ Links ]
Rinderknecht, A., 2000b, La presencia de osteodermos en las extremidades posteriores de Glyptodon clavipes Owen, 1839 (Mammalia: Cingulata): Ameghiniana, 37(3), 369–373. [ Links ]
Rusconi, C., 1931, Lista de los Vertebrados fósiles del Plioceno superior de Buenos Aires: La Semana Médica, 38, 1 –19. [ Links ]
Rusconi, C., 1936, Distribución de los vertebrados fósiles del Piso Ensenadense: Boletín de la Academia Nacional de Ciencias de Córdoba, 33, 183–215. [ Links ]
Rusconi, C., 1937, Contribución al conocimiento de la Geología de la Ciudad de Buenos Aires y sus alrededores y referencia a su Fauna: Actas de la Academia Nacional de Ciencias de la República Argentina, 10, 177–384. [ Links ]
Saint–André, P.A., De Iuliis, G., 2001, The smallest and most ancient representative ofthe genus Megatherium Cuvier, 1796 (Xenarthra, Tardigrada, Megatheriidae), from the Pliocene of the Bolivian Altiplano: Geodiversitas, 23(4), 625–645. [ Links ]
Scillato–Yané, G.J., 1980, Catálogo de los Dasypodidae fósiles (Mammalia, Edentata) de la República Argentina, en Actas del II Congreso Argentino de Paleontología y Bioestratigrafía y I Congreso Latinoamericano de Paleontología, Buenos Aires 1978: Buenos Aires, 3, 7–36. [ Links ]
Scillato–Yané, G.J., 1982, Los Dasypodidae (Mammalia, Edentata) del Plioceno y Pleistoceno de Argentina: La Plata, Argentina, Universidad Nacional de La Plata, tesis doctoral, 159pp. [ Links ]
Scillato–Yané, G.J., Carlini, A.A., 1998a, Nuevos Xenarthra del Friasense (Mioceno medio) de Argentina: Studia geologica salmanticensia, 34, 43–67. [ Links ]
Scillato–Yané, G.J. Carlini, A.A., 1998b, Un gigantesco gliptodonte en los alrededores de la ciudad de La Plata: Revista Museo 2, 11, 45–48. [ Links ]
Scillato–Yané, G.J., Carlini, A.A., Vizcaíno, S.F. Ortiz Jaureguizar, E., 1995, Los Xenarthros, en Alberdi, M.T., Leone, G., Tonni, E.P. (eds.), Evolución biológica y climática de la región Pampeana durante los últimos cinco millones de años. Un ensayo de correlación con el Mediterráneo occidental: Madrid, Museo de Ciencias Naturales, Consejo de Investigaciones, 12, 183–209. [ Links ]
Scillato–Yané, G.J., Carlini, A.A., Tonni, E.P. Noriega, J.I., 2005, Paleobiogeography of the late Pleistocene pampatheres of South America: Journal of South American Earth Sciences, 20(1–2), 131–138. [ Links ]
Serres, M., 1865, Deuxième note surle squelette du Glyptodon clavipes: Comptes rendus des séances de l' Académie des Sciences, 61(12), 457–466. [ Links ]
Simpson, G. G., 1930, Holmesina septentrionalis, extinct giant armadillo of Florida: American Museum Novitates, 442, 1–10. [ Links ]
Simpson, G.G., 1945, The Principles of Classification and a Classification of Mammals: Bulletin of the American Museum of Natural History, 85, 350 pp. [ Links ]
Soibelzon, E., 2008, Los Mamíferos del Ensenadense (Pleistoceno Inferior –Medio) del Este de la Región Pampeana, con énfasis en los Xenarthra. Bioestratigrafía, Diversidad y Correlaciones Biogeográficas: La Plata, Argentina, Universidad Nacional de La Plata, tesis doctoral, 304 pp. [ Links ]
Soibelzon, E., Tonni, E.P., 2009, Early–Pleistocene Glaciations in Argentina (South America) and the Response of Mammals: The Case of the Pampean Region: Current Research in the Pleistocene, 26, 175–177. [ Links ]
Soibelzon, E., Zurita, A.E., Carlini, A.A., 2006a, Glyptodon munizi Ameghino (Mammalia, Cingulata, Glyptodontidae): redescripción y anatomía: Ameghiniana, 43(2), 377–384. [ Links ]
Soibelzon, E., Carlini, A. A., Tonni, E.P., Soibelzon, L.H., 2006b, Chaetophractus vellerosus (Mammalia: Dasypodidae) in the Ensenadan (Early–Middle Pleistocene) of the southeastern Pampean region (Argentina). Paleozoogeographical and paleoclimatic aspects: Neues Jahrbuch fur Geologie und Palaontologie–Monatshefte, 12, 734–748. [ Links ]
Soibelzon, E., Danielle, G., Negrete, J., Carlini, A.A., Plischuk, S., 2007, Annual diet of little hairy armadillo Chaetophractus vellerosus Gray, 1865 (Mammalia, Dasypodidae) in Buenos Aires province, Argentina: Journal of Mammalogy, 88(5), 1319–1324. [ Links ]
Soibelzon, E. , Tonni, E. P, Bidegain, J. C. , 2008a, Cronología, magnetoestratigrafía y caracterización bioestratigráfica del Ensenadense (Pleistoceno inferior–medio) en la ciudad de Buenos Aire: Revista de la Asociación Geológica Argentina, 63(3), 421–429. [ Links ]
Soibelzon, E., Gasparini, G.M., Zurita, A.E., Soibelzon, L.H., 2008b, Las "toscas del Río de La Plata" (Buenos Aires, Argentina). Análisis paleofaunístico de un yacimiento paleontológico en desaparición: Revista del Museo Argentino de Ciencias Naturales, 10, 291–308. [ Links ]
Soibelzon, E., Carlini, A.A., Zurita, A.E., Tonni, E.P., Bidegain, J.C., Rico, Y., 2008c, Magneto y Bioestratigrafía del Sector Noreste de Buenos Aires (resúmen), en XVII Congreso Geológico Argentino: Jujuy, Argentina, 1049–1050. [ Links ]
Soibelzon, E., Prevosti, F.J., Bidegain, J.C., Rico, Y., Verzi, D.H., Tonni, E.P., 2009, Correlation of late Cenozoic sequences of southeastern Buenos Aires province: Biostratigraphy and magnetostratigraphy: Quaternary International, 210(1–2), 51–56. [ Links ]
Soibelzon, L.H., Soibelzon, E., Nedoszitko, R., Rodriguez, J., Medina, M., 2001, Arctotherium latidens (Ursidae: Tremarctinae) en el Ensenadense de la República Argentina: Ameghiniana, 38(4), 41R. [ Links ]
Tauber, A.A., Di Ronco, J., 2000, Nuevo hallazgo de placas ventrales de Glyptodon Owen, 1839 (Mammalia, Cingulata, Glyptodontidae) en la provincia de Córdoba, República Argentina: Boletín de la Academia Nacional de Ciencias, 64, 336–347. [ Links ]
Tonni, E. P. , Berman, W., 1988, Sobre dos especies de Glyptodon (Mammalia, Cingulata) del Pleistoceno tardío de la provincia de Buenos Aires. Su importancia bioestratigráfica en 2° Jornadas Geológicas Bonaerenses: Bahía Blanca, Actas 109–116. [ Links ]
Tonni, E.P., Fidalgo, F., 1982, Geología y Paleontología de los Sedimentos del Pleistoceno en el Área de Punta Hermengo (Miramar, Prov. de Buenos Aires, Rep. Argentina): Aspectos Paleoclimáticos: Ameghiniana 19(1–2), 79–108. [ Links ]
Tonni, E.P., Soibelzon, E., Cione, A.L., Carlini A.A., Scillato–Yané, G.J, Zurita, A.E., Paredes–Rios, F., 2009, Mammals from the Pleistocene of the Tarija Valley (Bolivia). Correlation with the Pampean chronological Standard: Quaternary International, 210, 57–65. [ Links ]
Ubilla, M., Perea, D., Lorenzo, N., Gutierrez, M., Rinderknecht, A., 2008, Fauna cuaternaria continental, en Perea, D. (ed.), Fósiles de Uruguay: Montevideo, DIRAC–Facultad de Ciencias, 283–314. [ Links ]
Verzi, D.H., Deschamps, C.M., Tonni, E.P., 2004., Biostratigraphic and palaeoclimatic meaning of the Middle Pleistocene South American rodent Ctenomys kraglievichi (Caviomorpha, Octodontidae): Palaeogeography, Palaeoclimatology, Palaeoecology, 212(3–4), 315–329. [ Links ]
Vizcaíno, S.F., Bargo, M.S., 1993, Los armadillos (Mammalia, Dasypodidae) de La Toma (Partido de Coronel. Pringles) y otros sitios arqueológicos de la Provincia de Buenos Aires. Consideraciones paleoambientales: Ameghiniana, 30(4), 435–442. [ Links ]
Vizcaíno, S.F., Bargo, M.S., Cassini, G.H., 2006, Dental occlusal surface area in relation to food habits and other biologic features in fossil Xenarthrans: Ameghiniana, 43(1), 11–26. [ Links ]
Wetzel, R.M., 1982, Systematics, distribution, ecology and conservation of South American Edentates: Special Publication Pymatuning Laboratory of Ecology, 6, 345–375. [ Links ]
Wetzel, R.M., 1985, Taxonomy and distribution of armadillos, Dasypodidae, in Montgomery, G.G. (eds.), The Evolution and Ecology of armadillos, sloths and vermiliguas: Washington y Londres, Smithsonian Institution Press, 23–50. [ Links ]
Winge, H., 1923, Pattedyr–slaegter, v. 1: Copenhague, H. Hagerups, 360 pp. [ Links ]
Yepes, 1928, Los "Edentata" argentinos, sistemática y distribución: Revista de la Universidad de Buenos Aires, 2a serie, 5(1): 461–515. [ Links ]
Zurita, A.E., 2007, Sistemática y evolución de los Hoplophorini (Xenarthra, Glyptodontidae, Hoplophorinae. Mioceno tardío–Holoceno temprano). Importancia bioestratigráfica, paleobiogeográfica y paleoambiental: La Plata, Argentina, Universidad Nacional de La Plata, tesis doctoral, 367 pp. [ Links ]
Zurita, A.E., Ferrero, B., 2009, Una nueva especie de Neuryurus Ameghino (Mammalia, Glyptodontidae) en el Pleistoceno tardío de la Mesopotamia de Argentina. El registro más completo del género fuera de la región Pampeana: Geobios, 42(5), 663–673. [ Links ]
Zurita, A.E., Scillato–Yané, G.J., Carlini, A.A., 2005, Palaeozoogeographic, biostratigraphic and systematic aspects of the genus Sclerocalyptus Ameghino 1891 (Xenarthra, Glyptodontidae) of Argentina: Journal of South American Earth Sciences, 20, 120–129. [ Links ]
Zurita, A.E., Soibelzon, E., Carlini, A.A. 2006, Neuryurus (Xenarthra, Glyptodontidae) in the Lujanian (late Pleistocene– early Holocene) of the Pampean region: Neues Jahrbuch für Geologie und Palaontologie–Monatshefte, 2, 78–88. [ Links ]
Zurita, A.E., Carlini, A.A, Scillato–Yané, G.J., 2009a, Paleobiogeography, biostratigraphy and systematics of the Hoplophorini (Xenarthra, Glyptodontoidea, Hoplophorinae) from the Ensenadan Stage (early Pleistocene to early–middle Pleistocene): Quaternary International, 210(1–2), 82–92. [ Links ]
Zurita, A.E., Miño–Boilini, A.R., Soibelzon, E., Carlini, A.A., Paredes–Ríos, F., 2009b, The diversity of Glyptodontidae (Xenarthra, Cingulata) in the Tarija Valley (Bolivia): systematic, biostratigraphic and paleobiogeographic aspects of a particular assemblage: Neues Jahrbuch für Geologie und Palaontologie–Monatshefte, 251–252, 225–237. [ Links ]
Zurita, A.E., Soibelzon, E., Scillato–Yané, G.J., Cenizo, M.M., 2009c, The earliest record of Neuryurus Ameghino (Mammalia, Glyptodontidae, Hoplophorinae): Alcheringa, 33(1), 49–57. [ Links ]
Zurita, A.E., Soibelzon L.H., Soibelzon, E., Gasparini, G.M., Cenizo, M.M., Arzani, H., 2010, Accessory protection structures in Glyptodon Owen (Xenarthra, Cingulata, Glyptodontidae): Annales de Paleontologie, 96(1), 1–11. [ Links ]

