










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkRevista mexicana de ciencias geológicas
On-line version ISSN 2007-2902Print version ISSN 1026-8774
Rev. mex. cienc. geol vol.27 n.2 Ciudad de México Aug. 2010
New camelid (Artiodactyla: Camelidae) record from the late Pleistocene of Calama (Second Region, Chile): a morphological and morphometric discussion
Nuevo registro de camélido (Artiodactyla: Camelidae) del Pleistoceno final de Calama (Segunda Región, Chile): una discusión morfológica y morfométrica
Isabel Cartajena1,*, Patricio López2, and Ismael Martínez3
1 Departamento de Antropología, Facultad de Ciencias Social, Universidad de Chile, Ignacio Carrera Pinto 1045, 7800284 Ñuñoa, Santiago, Chile. * icartaje@uchile.cl
2 Universidad Católica del Norte, Instituto de Investigaciones Arqueológicas y Museo Gustavo Le Paige (IIAM), Calle Gustavo Le Paige 380, 1410000, San Pedro de Atacama, Chile.
3 Av. Irarrázaval 2061, Depto. 408, 7750041 Ñuñoa, Santiago, Chile.
Manuscript received: September 2, 2009.
Corrected manuscript received: January 7, 2010.
Manuscript accepted: January 21, 2010.
ABSTRACT
A description of fossil remains belonging to camelids from the Kamac Mayu site (Second Region, Chile) is presented. This record was recovered from deposits of gravel and sand on karstic formations eroded by rivers during the late Pleistocene. In addition to the remains of camelids, diverse fossil material composed ofHippidion saldiasi (Mammalia, Equidae), Macraucheniapatachonica (Mammalia, Litopterna), undeterminedxenarthrans (Mammalia, Xenarhra) and birds (Aves, Rheidae), were recovered, along with specimens of Planorbidae, Hydrobiidaee and Sphaeriidae (Mollusca) from deposits formed by the drainage of the Chiu Chiu–Calama basin. The comparative anatomical study of the fossil material enabled the taxonomic assignment of the camelid remains to Lama gracilis (Gervais and Ameghino 1880), constituting an unprecedented finding for this region of the country.
Key words: South American camelids, Lama gracilis, late Pleistocene, Northern Chile.
RESUMEN
En el presente trabajo se describe el material correspondiente a restos fósiles de camélidos del sitio Kamac Mayu (II Región, Chile). Este registro, fue recobrado a partir de excavaciones sistemáticas en depósitos de grava arenosa y arena dispuestos sobre una formación kárstica, erosionadas por cursos fluviales durante el Pleistoceno tardío. Además de los restos de camélidos se recobró material fósil diverso compuesto por Hippidion saldiasi (Mammalia, Equidae), Macrauchenia patachonica (Mammalia, Litopterna), xenarthros indeterminados (Mammalia, Xenarhra) y aves (Aves, Rheidae), junto a especímenes de Planorbidae, Hydrobiidaee and Sphaeriidae (Mollusca) dentro de depósitos formados por el desagüe de la antigua cuenca Chiu Chiu–Calama. El estudio anatómico comparado de los restos fósiles permitió la asignación taxonómica a Lama gracilis (Gervais y Ameghino 1880), constituyéndose en un hallazgo inédito para esta región del país.
Palabras clave: camélidos sudamericanos, Lama gracilis, Pleistoceno tardío, Norte de Chile.
INTRODUCTION
Studies undertaken on the present–day species of camelids, both wild and domesticated, in the Southern Cone of America, distributed in various regions of Peru, Bolivia, Argentina and Chile, have emphasised diverse aspects related to their taxonomy, geographical distribution, ecology conservation and economic use (Cardozo, 1974; Franklin, 1983; Torres, 1992; Lichteinstein and Vilá, 2002; Marín et al., 2007; Bonacic, 2008; among others). It should be noted that a large number of these issues have been addressed on archaeological reports of camelid remains (Wheeler, 1984; Mengoni–Goñalons and Yacobaccio, 2006; Cartajena et al., 2007). Also, fossil records of Camelidae have been widely treated in the South American literature (Ferrero, 2005, 2006; Menegaz, 2000; Menegaz et al., 1989, Menegaz and Ortiz Jaureguizar, 1995, Noriega y Ferrero, 2007; Ribeiro et al., 2007; Scherer et al., 2007; Ubilla, 2004; Ubilla et al., 2004, 2007). However, they have been relatively scarce in the case of Chile (López et al., 2005).
The first fossil remains belonging to South American Camelidae correspond to findings of deposits from the Late Pliocene (Marplatan Stage/Age, Barrancalobian Subage). Once the isthmus of Panama was formed, some Lamini forms (llamas) migrated to South America, where they radiated (Cione and Tonni, 1995; Menegaz and Ortiz Jaureguizar, 1995; Webb, 2006; Woodburne et al., 2006). The greatest diversity of South American genera was produced in the late Pleistocene, including taxa with different morphological traits, distribution, and marked differences in size, from large species of Hemiauchenia Gervais and Ameghino, 1880 and Palaeolama Gervais, 1867, to smaller species within the genus Lama such as Lama gracilis (Gervais and Ameghino, 1880). Recent paleontological studies based on material from the northeast of Brazil (Guérin y Faure, 1999), from Córdoba Province (Tauber, 1999), from Entre Ríos Province (Ferrero, 2005; Scherer et al., 2007), and from the of Pampa Bonaerense (Menegaz and Ortiz–Jaureguizar, 1995; Menegaz, 2000) have aimed to systematize an extensive list of camelid findings, in order to amend former classifications and identify new species. Menegaz' work (Menegaz, 2000; Menegaz et al., 1989; Menegaz and Ortiz–Jaureguizar, 1995) is particularly interesting as he subordinates the genus Vicugna Lesson, 1842 as a subgenus of the Lama Cuvier, 1800, which would be represented by the Lama (Vicugna) vicugna (Molina, 1782) and the Lama (Vicugna) gracilis (Gervais and Ameghino, 1880). The latter corresponds to a species whose size and cranial characters (incisors) locate it between the vicuña and the guanaco (Menegaz and Ortiz–Jaureguizar, 1995; Guérin and Faure, 1999). The gracile camelid remains were recovered from Bolivia to the Tierra del Fuego deposits of middle to late Pleistocene and early Holocene age, and the majority of the material was collected from different archaeological sites (Menegaz and Ortiz–Jaureguizar, 1995; Menegaz, 2000). Recent DNA molecular analysis of dental pieces from chilean patagonian remains show however that these remains previously classified as Lama gracilis corresponds to Vicugna vicugna. Therefore, this evidence suggests that these remains do not belong to an extinct species but rather to a large vicuña with a wider geographical distribution as known before (Weinstock et al., 2009).
In the present study we describe material corresponding to fossil remains of camelids from the Kamac Mayu site (Second Region, Chile). The record was obtained by systematic excavations in deposits of gravel and sand set out on a karst formation eroded by river courses during the late Pleistocene (Figures 1a, 1b, 1c, 1d). As well as the Camelidae record, diverse fossil material was recovered, including Hippidion saldiasi Roth, 1899 (Mammalia, Equidae), Macrauchenia patachonica Owen, 1838 (Mammalia, Litopterna), undeterminated xenarhrans (Xenarhra, Mammal), and birds (Aves, Rheidae) (Alberdi et al., 2007; Gelfo et al., 2008), as well as specimens of Planorbidae, Hydrobiidaee and Sphaeriidae (Mollusca) within deposits formed during the draining of the former basin in Chiu Chiu, Calama (May et al., 2005). The morphological and osteometric analyses of cranial and post–cranial elements enabled the taxonomic assignment of the remains to Lama gracilis, not seen before in this region of the country.
GEOLOGICAL SETTING
The paleontological site of Kamac Mayu (68° 54' 40'' W, 22° 26' 30'' S) is located in the city of Calama (2250 m a. s. l.), in a fluvial basin containing sandy gravel and sand deposits, 100–300 m away from the Loa river. According to Chong–Díaz and Jensen (2004), four stages of depositation can be distinguished at the Kamac Mayu site:
Lacustrine stage. Represented by a stratified unit of disintegrated marl with diatomite lenses from the Pliocene–Pleistocene age. Due to its lithology, this lacustrine unit can be assigned to the Chiu Chiu Formation from the Pliocene–Pleistocene age, defined by Naranjo and Paskoff (1981).
Karstic stage. Represented by an eroded surface formed by dissolution cavities. In the area under study, the walls of these cavities have a hard covering that is 1 to 3 mm thick and whose origin stems from calcium carbonate precipitation. This contrasts with the yellow marl, whose aspect is pulverulant and brittle. A fluvial system, characterized by clastic sediments, occurred after this event.
Fluvial stage. A third stage of fluvial origin represented by sandy gravel and sand deposits that fill multiple channels formed above the karstic horizon. The stratigraphic distribution comprises fine sequences of centimetric magnitude with crossed concave stratification. The fluvial stage was formed prior to the formation of the Loa River canyon and corresponds to the interval in which the fossil remains were deposited. The deposits with fossil remains reach a maximum depth of approximately 1.80 m. (Figure 1)
Calcareous cementation stage. After the gravel sedimentation, the meteoric waters, saturated in calcium carbonate, percolated through the gravel that fills the channels. This process enabled the precipitation of calcite in intergranular spaces leading to the cementation of clasts.
The presence of fossil remains belonging to Macrauchenia patachonica and Hippidion saldiasi suggests a late Pleistocene age for the deposits (Alberdi and Prado, 2004; Alberdi et al., 2007). Two bone samples from the gracile camelids were sent to be dated. Unfortunately, they did not contain enough organic material. However, the remains of Hippidion saldiasi from the neighboring site Betecsa–1 (68° 54' 44'' W, 22° 26' 30'' S) were dated; the dating carried out on the right M3 layer showed a 14C age of 21,070 ± 100 B. P. (not calibrated. GrA–29389, Centrum voor Isotopen Onderzoek, Holland). In addition, the dating of a basicranium fragment belonging to the same Hippidion saldiasi showed an age of 21,380 ± 100 B. P. (not calibrated. GrA–29388, Centrum voor Isotopen Onderzoek, Holland) (Alberdi et al., 2007). The faunistic and sedimentolgical affinities between the Betecsa–1 and Kamac Mayu suggest a similar age for both sites, equivalent to Lujanian stage of the South American Land Mammal Ages (SALMA).
MATERIAL AND METHODS
Morphometric analysis was performed in order to study the change and variation of the fossil sample. Size was studied by using osteometric methods and comparing measurements with fossil and modern camelid samples. Measures were taken following criteria presented in von den Driesch (1999) and Menegaz (2000). Multivariate analysis is commonly employed for morphometric studies, specially Principal Components (Menegaz et al., 1989). However, in this case the number of observations is too small for the use of parametric statistics. Instead, we used bivariate scaterplots in order to compare the samples. In addition, a comparative anatomical study focused on the identification and description of diagnostic landmarks or characters was performed. For the morphological description of the material we use the criteria defined by Menegaz (2000), and Scherer et al. (2007) for the dental nomenclature.
For the identification and comparison of the sample considered in this study, we used camelid fossil remains from the Colección División Paleontológica de Vertebrados housed at the Museo de La Plata (Argentina) and the Collection of Mammiféres fossils, Muséum National d'Histoire Naturelle (Francia). In the same way, we also used as reference present–day collections and measurements of Lama guanicoe and Vicugna vicugna from Staatssammlung für Anthropologie und Palaoanatomie, Munich (Germany), Zoologische Staatssammlung, Munich (Germany), Zoologisches Museum, Berlin (Germany), Natural History Museum, London (England) and Departamento de Antropología Universidad de Chile, Santiago (Cartajena 2003).
Abbreviations
Institutional abreviations.
KM, Kamac Mayu; MLP, Museo de La Plata; MNHNP–TAR, Musee d'Histoire Naturelle Paris,fossil remains recovered from Tarija locality (Bolivia).
Anatomical abbreviations
DP, Deciduous premolar teeth; M, Molars; I, Incisors; DI, Deciduous incisors.
SYSTEMATIC PALEONTOLOGY
Order Artiodactyla Owen, 1848
Family Camelidae Gray, 1821
Subfamily Camelinae Gray, 1821
Genus Lama Cuvier, 1800
Lama gracilis (Gervais and Ameghino, 1880)

Auchenia gracilis Gervais y Ameghino, 1880, p. 115.
Auchenia frontosa Gervais y Ameghino, 1880, p. 117.
Auchenia lujanensis Ameghino, 1889, p. 586, fig. 8.
Hemiaucheniaprístina Ameghino, 1891, p. 138, fig. 30
Palaeolama crequii var. provicugna Boule and Thevenin, 1920, p. 180. plate 20, fig. 1.
Lama gracilis López–Aranguren, 1930, p. 113, fig. 16, left.
Vicugna vicugna Cabrera, 1932, p. 116.
Lama (Vicugna) gracilis Menegaz, Goin y Ortiz–Jaureguizar 1989, p. 161, plate 1, figs. 1–4.
Diagnosis. The size of metapodials is smaller than the Lama guanicoe and slightly larger than the Vicugna vicugna, and the metapodials are comparatively more gracile than in these species. The lower incisors are slightly imbricated; with a thick layer of enamel on the labial face which becomes much thinner towards the lingual face of the tooth layer, especially in the I1. The incisors show bicelated wear facets at their distal extremes. In vicuñas this feature is similar but much developed since the wear facet covers a wider surface at the lingual face of the tooth. On the other hand, in guanacos wear facets are much less developed and over the incisors borders, the dentine is exposed as a fine band on the lingual and labial enamel (Menegaz et al. 1989, p. 160). The wearing down of the inferior incisors is located in the sub quadrangular section, unlike the Vicugna vicugna where this section is enlarged, with a sub rectangular and sub elliptical section.
Materials. KM/SC/SA/1, right metatarsal; KM/SC/SA/2, right metacarpal; KM/SC/SA/3, incomplete metapodial; KM/SC/SA/4, left metacarpal; KM/SC/SA/5, incomplete metapodial; KM/SC/SA/6, incomplete metapodial; KM/ SC/SA/7, incomplete left tibia; KM/SC/SA/8, incomplete metapodial; KM/SC/SA/13, incomplete metapodial, KM/SC/SA/17, right metacarpal; KM/SC/SA/62, fourth tarsal; KM/SC/SA/22, incomplete right femur; KM/SC/SA/23, incomplete radius; KM/SC/SA/24, incomplete right femur; KM/SC/SA/25, incomplete right radius; KM/SC/SA/26, incomplete right humerus; KM/SC/SA/27, incomplete right humerus; KM/SC/SA/28, incomplete left humerus; KM/SC/SA/30, incomplete cervical vertebrae; KM/SC/SA/31, incomplete left humerus; KM/SC/SA/33, incomplete right calcaneus; KM/SC/SA/35, incomplete cervical vertebrae; KM/SC/SA/36, caudal vertebrae; KM/SC/SA/37, incomplete first phalanx; KM/SC/SA/38, incomplete rib; KM/SC/SA/39, incomplete rib; KM/SC/SA/40, incomplete rib; KM/SC/SA/41, incomplete rib; KM/SC/SA/42, incomplete rib; KM/SC/SA/43, incomplete rib; KM/SC/SA/44, incomplete left humerus; KM/SC/SA/45, incomplete right tibia; KM/SC/SA/46, caudal vertebrae; KM/SC/SA/47, thoracic vertebrae; KM/SC/SA/48, incomplete right metatarsal; KM/SC/SA/53, incomplete right humerus; KM/SC/SA/56, sacrum; KM/SC/SA/62, scapula; KM/SC/SA/63, incomplete first phalanx; KM/SC/SA/64, incomplete first phalanx; KM/SC/SA/65, second phalanx; KM/SC/SA/66, incomplete rib; KM/SC/SA/67, second phalanx; KM/SC/SA/68, incomplete rib; KM/SC/SA/69, incomplete cervical vertebrae; KM/SC/SA/70, incomplete thoracic vertebrae; KM/SC/SA/71, incomplete thoracic vertebrae; KM/SC/SA/72, incomplete thoracic vertebrae; KM/SC/SA/73, skull; KM/SC/SA/74, incomplete right mandible; KM/SC/SA/75, incomplete left mandible; KM/SC/SA/76, incomplete left femur; KM/SC/SA/77; incomplete left maxilla; KM/SC/SA/78, incomplete left maxilla; KM/SC/SA/79, incomplete metapodial; KM/SC/SA/80, incomplete rib; KM/SC/SA/81, incomplete rib; KM/SC/SA/82, incomplete rib; KM/SC/SA/83, incomplete cervical vertebrae; KM/SC/SA/84, incomplete lumbar vertebrae; KM/SC/SA/85, incomplete lumbar vertebrae; KM/SC/SA/86, incomplete thoracic vertebrae; KM/SC/SA/87, incomplete thoracic vertebrae; KM/SC/SA/88, incomplete second phalanx; KM/SC/SA/89, incomplete rib; KM/SC/SA/90, incomplete rib; KM/SC/SA/91, incomplete rib; KM/SC/SA/92, incomplete rib; KM/SC/SA/93, incomplete mandible; KM/B3/C3/RP15, complete left femur; KM/B3/C3/RP16, complete right radius–ulna; KM/C3/B4/RP29, complete left metatarsal; KM/A4/C3/RP32, incomplete thoracic vertebrae; KM/A3/C1/SRP, second phalanx; KM/B3/C3/SRP, patella; KM/ B4/C3/SRP, incomplete right maxilla.
Preservation. The fossil record pertaining to camelids is composed of 37 specimens, including axial and appendicular elements, which showed a regular state of conservation due to fissures produced by the pressure of sediments and adhesion of salt crystals typical of this zone (López and Labarca, 2005). Taphonomic alterations such as weathering or damage produced by other natural agents such as carnivores were not observed.
DESCRIPTION AND DISCUSSION
Skull, mandible, and dentition
Skull
Four fragments of craniums have been recovered. Three of these specimens correspond to fragments of maxillaries, whereas one of the samples is a relatively complete cranium of a young individual. In the case of the maxillaries, the specimen KM/SC/SA/77 shows the DP3, DP4 and M1 which suggests an age of 4 to 7 months (de la Torre, 1981). In the specimen KM/SC/SA/78 only the M1 and M2 are conserved, indicating an age of between 14 to 20 months. On the other hand, the fragment of maxillary KM/B4/C3/SRP is covered with an extremely hard calcareous sediment, nevertheless, the presence of DP3, DP4 and M1 suggests a similar age to that proposed for the specimen KM/SC/SA/77.
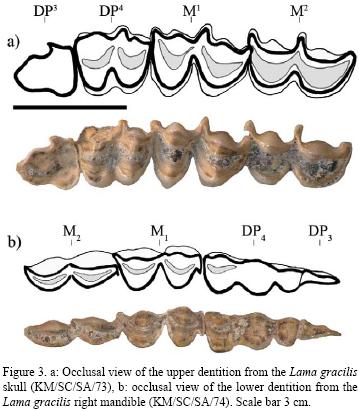
The cranium (KM/SC/SA/73) consists of both maxillaries, a part of the two frontals, a fragment of the right parietal (with a total absence of left one), both lacrimals, a part of the nasals, the temporal, the right zygomatic and part of the left one, as well as both supermaxillaries and the palatine, although there is a total absence of the premaxillaries. Both dental series are in a good state of conservation and are represented by the DP3 whose occlusal surface exhibits an advanced tooth wear, the DP4 shows intermediate wear, the M1 does not show much wear. As the M2 is still in the process of eruption, the age of this individual has been defined around 14–20 months (de la Torre, 1981). The relation between the molariform series is relatively convergent and there is a deep, well defined palatine notch (Table 1, Figure 2a).

One remarkable trait of the cranium (KM/SC/SA/73) recovered from the Kamac Mayu site, is the great development of the Foramen ethmoidale. Herre (1952) proposes that this feature is evident in guanacos, whereas it is poorly developed or absent in vicuñas. However, this feature cannot be considered as diagnostic, given that it has been observed in some recent camelids (Cartajena, 2003).
Mandible
The sample includes two fragments of mandibles (left and right) of the same individual and a mandibular symphysis with a complete dental series of another individual. The right mandible (KM/SC/SA/74, Figures 2b and 3b) shows a better state of preservation; the molar series DP3, DP4, M1 and M2 have been conserved, which suggests an age between 14 to 20 months (de la Torre, 1981); in this specimen the coronoid process and the mandibular symphysis are broken. The masseteric fossa is marked and deep and the mandibular condyle is low, with a triangular section and shows ample facets with well marked boarders (Menegaz, 2000). In the fragment of the left mandible (KM/SC/SA/75) part of the molar series DP4, M1 and M2 can be observed (the DP3 is broken at the root) the mandibular angular, the mandibular condyle, the coronoid process and the mandibular symphysis are broken.
In the case of mandibular symphysis (KM/SC/SA/93, Figure 2c), the development and damage of the lower deciduous incisors indicate an individual age of less than 15 months (de la Torre 1981). Measurements in Table 2.

Dentition
Upper deciduous dentition. The DP3 and DP4 from the specimens KM/SC/SA/77, KM/B4/C3/SRP, and KM/SC/ SA/73 are moraliform, with two distinct, clearly defined U–shaped lobes. The mesostyle is projected towards the labial surface more than the parastyle and metastyle. In the DP3 of the maxillary KM/SC/SA/77 the occlusal surface can be observed with greater clarity, whereas in KM/SC/SA/73 it is completely worn down and in the KM/B4/C3/SRP it is covered by calcareous sediment. In the DP3 the trigon fossa is elongated and less curved than the talon fossa; at the same time the protocone is not touching the hypocone; the latter is barely visible; it disappears completely into the cranium due to the advanced state of tooth wear. On the other hand, the DP4 is characterized by the even more marked prolongation of the mesostyle towards the labial surface. The trigon fossa and the talon fossa are also curved and unfolded; both lobes are U–shaped and subequal in size. The protocone and hypocone are united across the narrow valley. In the piece KM/SC/SA/73, the metastyle is practically absent due the damage to the DP4, whereas the parastyle takes on a quadrangular form.
Upper permanent dentition. Various stages of wear can be observed in the four samples studied. In the cranium (KM/SC/SA/73) M1 has also almost totally erupted, whereas the M2 is in the initial stage of eruption (Figure 3a). In the fragment of the maxillary KM/SC/SA/78 a similar situation can be seen, although the M2 is located further into the alveolar cavity. Lastly, in the specimens KM/SC/SA/77 and KM/BB5/C3/SRP the M1 is in the process of eruption, as is the M2, although the latter is more contained. In the M1 the medial lobe is larger than the distal lobe. Like the deciduous premolars, the mesostyle is extended towards the lingual surface, surpassing the parastyle and the metastyle. In the specimen KM/SC/SA/77 both lobes tend to be V–shaped, although this condition is due to the slight wear of this molar as this is not observed in the rest of the material in which U–shaped lobes predominates; this feature is comparable to that observed in a fragment of a left maxillary (MLP. 9. 473) referable to the Lama gracilis from the Vitel Lagoon (Pampean Formation), Argentina. In the M1 of the KM/SC/ SA/77, the trigon fossa is narrow and curved, unlike the talon fossa, which does not show such a sharp convexity. In the same way, the protocone shows a greater labial–lingual length than the hypocone. In the rest of the M1 these differences cannot be seen as both the fossas and the cones are very similar. On the other hand, all the M2 studied are in the process of eruption and significant differences between them are not present.
Lower deciduous dentition. The sample of the lower deciduous dentition is scarce and comprises of pieces of KM/SC/SA/74, i.e. the DP3 and the right DP4 and the DP4 from the left mandible from the same individual (KM/SC/ SA/75) (Figure 3b). The DP3 consists of only one lobe that has been transversely compressed and shows significant wear on the occlusal surface. The molariform DP4 shows two lobes with intermediate wear; the talonid fossa is absent, whereas the trigonid fossa is hardly visible, unfolded and curved. Due to the wear, the metastylid and the entostylid are absent and the parastylid can hardly be seen. The protoconid and the hypoconid are united, whereas the size of the hypoconulid is equivalent to 2/3 of both main lobes.
In the mandibular symphysis (KM/SC/SA/93) the deciduous incisors show a slight imbrication, with signs of wear in the case of the DI2 in the elliptical section and in the case of the DI3 of a sub rectangular type. Based on the analyses carried out through the use of X–rays taken from these samples, it was possible to ascertain that the enamel tended to be thicker on the labial face, reducing to a thin layer on the lingual face (Figure 4a and 4b).
Lower permanent dentition. As in the last case, the only samples of lower molars correspond to the two mandibles recovered (KM/SC/SA74 and KM/SC/SA/75). In both cases the M1 shows slight tooth wear whereas the M2 is in the process of eruption. The M1 has two lobes similar in size, with the metastylid and the parastylid barely visible, unlike the entostylid which sticks out towards the labial surface. The trigonid fossa and the talonid fossa show similar characteristics due to their size and to the slightly pronounced curve. In the labial surface both lobes are U–shaped and have a well defined and projected protostylid. In the case of the M2 the very scarce degree of the eruption of the tooth does not allow for a detailed description. The mesial lobe is more developed than the distal lobe; the parastylid and the entostylid are scarcely developed; only the metastylid can be observed.
Axial skeleton
Scapula
The only scapula recovered from Kamac Mayu is very fragmented and only the neck of scapula and part of the spine of scapula can be seen; the acromion process is absent. On the other hand, in the caudal border in the area of the neck of scapula, a ridged surface can be observed, situated on the border of a slightly marked tuberosity. Due to the state of conservation, only one measurement was obtained (Table 3).

Humerus
All the humeri from the Kamac Mayu site correspond to young individuals from which metric data was not obtained. From the five samples recovered, three correspond to distal diaphyses (KM/SC/SA/44; KM/SC/SA/53; KM/SC/SA/27) all of them not presenting the humerus condyle although the olecranon fossa is deep and marked. The piece KM/SC/SA/27 possesses the majority of the diaphyses and here the helical torsion is less pronounced than in comparison with adult camelidae humerus (Cartajena, 2003). The proximal fragments (KM/SC/SA/26; KM/SC/SA/31) do not have the greater tubercle or the lesser tubercle fusion and in the best conserved piece KM/SC/SA/26, the deltoid tuberosity is hardly visible.
Radius–ulna
Among the bones that correspond to individual adults there is a radius–ulna (KM/B3/C3/RP16) in a very bad state of conservation due to fractures caused by sediment pressure (Figures 5a and 5b). Likewise, within the sample, two unfused radial diaphyses from young individuals were counted (KM/SC/SA/23; KM/SC/SA/25). Regarding the complete piece, the olecranon is long and tall, but compressed in the lateral–medial section. Both proximal articular facies are wide and deep with a shallow depression on the extremity of the head of the radius, whereas the lateral tuberosity and the lateral coronoid process are extremely pronounced. The distal articular surface is made up of three facies of the carpi: radial, intermediate, and ulnar; they have similar dimensions.
The width of the radius–ulna (KM/B3/C3/RP16) is comparable to that observed in specimens belonging to vicuña. However, the specimens from the Kamac Mayu site and also from the MLP are classified in the 'extremely high' range of the established size of present–day vicuñas (Table 4). This would suggest that the differences between the gracile camelid (Lama gracilis) and the vicuña radius–ulna are associated with the length of the bones. Nevertheless, the muscular insertion zone in the specimens from the Kamac Mayu site are much more pronounced in relation to present–day vicuñas.

Metacarpals
Three metacarpals were recovered: a proximal fragment of a right metacarpal (KM/SC/SA/2), a diaphysis and a distal epiphysis from a left metacarpal from an adult animal (KM/SC/SA/4) and a right proximal fragment (KM/SC/SA/17). The specimen KM/SC/SA/2 shows a smooth articular facet for the fourth carpal, with rounded edges that show that the bone is not well developed and therefore corresponds to a young individual. Furthermore, the absence of a division between the articular facets for the second carpal and third carpal can be seen. On the other hand, the articular facets in the specimen KM/SC/SA/17 shows well marked borders with concave surfaces; the result of development and which correspond to an adult animal. As in the case of the radius–ulna, in terms of size the measurements from the proximal diaphysis of the adult individual show a similarity with the largest vicuñas (Table 5).

First phalanges
The first phalanges correspond to a proximal fragment (KM/SC/SA/63) and to a distal extreme (KM/SC/SA/64), both from the anterior member of adult animals, as well as a proximal fragment with an unfused articular surface (Table 6, Figure 5c). In the extreme proximal of the adult individual (KM/SC/SA/63), specifically on the palmar surface, the flexor tuberosity is long and it is distally extended. Furthermore, the articular surface on this specimen is quadrangular, whereas the diaphysis has a pronounced triangular section and tends to be more elongated and gracile compared to the vicuña and guanaco. In the extreme distal the axial distal trochlear lobe and the abaxial lobe are absent, due to fracture, therefore it is not possible to carry out a detailed description of this specimen.

Sacrum
The only recovered sacrum (KM/SC/SA/56) has a complete fusion of the first, second and third vertebra; due to a postdepositional fracture the fourth and fifth vertebra are absent (Table 7, Figure 5d). This is also the case for a large part of the sagital crest, the articular processes and part of the articular facies of the ilium which are poster–oventrally extended. Between this articular facies and the cranial articular process there is a notch that is less marked in present–day wild Camelidae (Cartajena, 2003). This trait was observed in the specimen MLP. 71 .XI. 1–1 kept in the MLP, although in this case it is much more pronounced. All the sacral foramen are ovoidal with a distribution that tends to be more horizontal, whereas the opening of the sacrum is triangular, although the vertebral arch is less elevated than in the case of the vicuña and the guanaco.

Femur
The femur sample is one of the most complete and informative as it comprises a complete piece (KM/B3/C3/ RP15) and a distal diaphysis (KM/SC/SA/76) from two adult individuals, two unfused distal diaphysis (KM/SC/SA/24 and KM/SC/SA/28), and a diaphysis with the proximal epiphysis (KM/SC/SA/22) from a young animal.
Based on discreet morphological characteristics, some differences among wild camelids have been discussed (Cartajena, 2003). In the case of specimen KM/B3/C3/RP15 the fovea of the femoral head is deep and well marked, similar to the guanaco but different to the majority of the present–day vicuñas used as a comparison, in which the fovea of the femoral head is not pronounced. The trochanteric fossa is more similar to the guanaco than the vicuña as it is smaller and deeper. The rough line of the Lama gracilis shows a smooth, regular shape which is very similar to that of the vicuña, given that in the case of guanacos it is much more irregular and has a rigid surface. The Linea aspera in the Kamac Mayu specimen is distinct and has quite a prominent edge, similar to that of the vicuña.
In the distal diaphysis (KM/B3/C3/RP15), the edges of the trochlea are high and concave and extend towards the suprapatellar fossa. The fossa poplitea is deep, like in the case of the guanaco. The metric data taken from the Kamac Mayu sample show that the specimen is of medium size, between the guanaco and the vicuña, although it is more similar to the latter species (Table 8, Figures 5e and 5f).

Tibia
The remains of the tibias are scarce and in both cases they correspond to remains from young animals; one corresponds to an unfused distal diaphysis from a right tibia (KM/SC/SA/45) and the other corresponds to a reassembled distal epiphysis and diaphysis in a bad state of conservation from a left tibia (KM/SC/SA/7). In the surface diaphysis of piece KM/SC/SA/45 the notches for astragalus show long, deep, oval, facets parallel to the anteroposterior axis.
Patella
The anterior and posterior surfaces of the patella are oval shaped. Furthermore, on this surface the development of a convexity that appears in alpacas, llamas and in some vicuñas and guanacos is not transversally observed (Cartajena, 2003). Measurements in Table 9.

Calcaneus
Only one calcaneus was recovered from Kamac Mayu site. This piece corresponds to a right calcaneus from a young individual (KM/SC/SA/33) whose distal extreme is absent due to a postdepositional fracture; the calcaneal tuberosity is unfused.
Metatarsals
The sample from Kamac Mayu comprises two almost complete metatarsals (KM/SC/SA/1; KM/C3/B4/RP29) with the distal epiphyses unfused. In addition, there is a proximal fragment (KM/SC/SA/48) from a right metatarsal that is a slightly eroded, thus it was not possible to collect metric data from it (Table 10). In all specimens the proximal surface is characterised by two articular facets, one medial for the articulation with the third tarsal (triangle shaped) and a second lateral facet for the fourth tarsal (oval shaped). In the plantar surface there is quite a deep, open groove that extends along almost the entire bone extension, with the exception of the distal quarter. As in the case of the metacarpals, the metatarsals are diagnostic anatomical units for the identification of the Lama gracilis (Figure 6a, 6b and 6c). This is due to their characteristically gracile metapodials and their intermediate length, which is between the guanaco and the vicuña. One interesting aspect is that the angle formed in the union between both facets tends to be more open than in vicuñas and guanacos and in the bones analysed it protrudes between 85° and 95° (Figure 7).
Despite the fact that all the specimens correspond to young individuals, some metric traits have been compared in order to obtain a general panorama of the dimensions of the metatarsals from Kamac Mayu with the samples from Tarija (Bolivia), Argentina (Table 11) and from the present–day vicuñas and guanacos (Figures 8a and 8b).
The specimens from Kamac Mayu are of different size in comparison to the specimens of present–day vicuñas in terms of length, despite the fact that they are unfused, which brings them closer to the Tarija specimens. The difference in the breadth of the diaphysis and the proximal epiphysis is less than in the present–day vicuñas.
Considering the ratio between the GL (Greatest length) and Bp (Greatest breadth of the proximal end), the metatarsals from Kamac Mayu are characteristically longer than those of the vicuña (Figure 8). In this respect, Menegaz (2000) states that the gracility index (Bp:GL) is 0,14 for the Lama gracilis. However, according to the IG obtained for the specimens from Kamac Mayu (IG 0.13), it can be seen that these are within the range of gracile individuals (IG 0.12–IG 0,14. See Table 12). Nevertheless, it should be noted that the specimens from Kamac Mayu are not fused; therefore the index is closer to the values obtained for the Lama graclis by Menegaz (2000, p. 45). We also compared the value of a non fused guanaco metatarsal in a similar ontogenic stage (IG 0.16), which differs considerably from those of Kamac Mayu, Lama gracilis and present day vicuñas suggesting that that fossil remains do not belong to adult nor juvenile L. guanicoe (see Table 12).

Fourth tarsal
The only tarsal bone corresponds to a very well preserved fourth tarsal (KM/SC/SA/62). Measurements in Table 13.

Second phalanges
Among the second phalanges four specimens were recovered, three of which correspond to adult animals and one belongs to a young individual due to the absence of fusion of the proximal articular surface. The specimen KM/SC/SA/88 is extremely eroded therefore it was not considered for obtaining metric data (Table 14). In the case of the phalanges KM/SC/SA/65 and KM/SC/SA/67, these correspond to the posterior member. In both specimens the distal articular surface shows an inclination towards the axial and a very marked line of separation between the abaxial and the axial portion of the articular surface can be observed. As is shown in the graph of the Figure 8e, one of the specimens is within the lower range established for present–day guanacos, and as with the metatarsals, there is an extremely elongated tendency in the case of the inferior extremities.
CONCLUSIONS
Through a comparative anatomical study we managed to identify some diagnostic traits that enabled us to assign the studied fossil material to Lama gracilis; one of these traits is the morphology of the incisors, specifically the distribution of the enamel and the imbrication of the incisors. In addition, by means of osteometric comparison is it possible to observe that the size of the remains from the Kamac Mayu site is above the established size–range for present–day vicuñas. They are closer to samples of camelid fossils belonging to the Lama gracilis (MLP) and the Palaeolama crequii provicugna (NMNHP) and much lower than the collection of present–day guanacos (Cartajena, 2003).
One of the diagnostic traits of the Lama gracilis is associated with the length and gracility of the metapodials. As mentioned in above, although the metapodials registered belong to young individuals whose distal epiphyses are not fused, their length is greater than the specimens of present–day adult vicuñas and also juvenile (non fused) modern guanacos used as reference. However, with the non fussed guanaco the most notable differences are related to the breadth. Despite the fact that we measure young animals, we used them as comparison parameters. They show the gracile character of the specimens; the gracility index is similar to that of the vicuña. In the case of the long bones that were recovered belonging to adult animals, they are also intermediate sized, between the present–day vicuña and guanaco. However, it is noteworthy that the muscular insertions are marked in both the long and flat bones.
The remains of the Lama gracilis from the Kamac Mayu site are similar morphologically to the remains located in strata from the middle and late Pleistocene from the Province of Buenos Aires (Menegaz et al., 1989). In Chile, the records of this species are scarce and are limited to a few findings in sites in continental Patagonia dated from around the late Pleistocene (Prieto and Canto, 1997; Massone and Prieto, 2004), that have not been described in detail. According to Menegaz and Ortiz–Jaureguizar (1995, p. 323), the Lama gracilis was a herbivorous form with grazing habits, found linked to peneplain pasturelands. The presence of this species in deposits from the late Pleistocene associated with the Hippidion saldiasi and Macrauchenia patachonica suggests open environments with abundant pasture land and foraging material. In this respect, isotopic analysis carried out on the bone and teeth of an individual of Hippidion saldiasi from the Betecsa 1 site show typical values of animals that only feed on C3 grass (Alberdi et al., 2007), which could be due to the altitude where the site is located (2250 m a. s. l.).
During the Finipleistocene and the Early Holocene the distribution of the Lama gracilis covered the Pampean Region, Patagonia Argentina, and the north of Uruguay (Ubilla, 2004). Nevertheless, considering that their distribution probably also reached the north of Chile we can suggest that the specimens from Tarija (TAR 165, 176, 177 and 206 used as a comparison in this work), could correspond to a more septentrional distribution of L. gracilis. Although these latter remains should be studied in detail, the metric ranges of the specimens assigned to Paleolama crequii provicugna (Mones, 1986) are closer to those observed for the L. gracilis. In this context, the present study suggests a wider geographic spatial distribution of gracile llamas during the Pleistocene, and the use of not only the low plains but also the high plains, as in the case of Kamac Mayu.
These results contribute to the discussion of new molecular evidence regarding the assignation of fossil records from Patagonia to Vicugna vicugna instead of Lama gracilis (Weinstock et al., 2009). In our study, morphological and osteometrical comparisons of cranial and postcranial anatomical units allow us to identify the presence of a gracil camelid with similar characteristics of a Lama gracilis (Menegaz et al., 1989). The morphological and size differences observed between the Lama gracilis with present day vicuñas, raises the question of whether it is related to an extinct specie or they correspond to a late Pleistocene and early Holocene vicuña.
ACKNOWLEDGMENTS
Research on the Kamac Mayu site was funded by Nawel Consultores (Chile). The authors thank Nuriluz Hermosilla (Departamento de Antropología, Universidad de Chile), Dr. M. Reguero and Dr. Javier Gelfo (Museo de La Plata, Argentina) and also to the Museo de La Plata (Argentina), Musee d'Histoire Naturelle Paris (France), Staatssammlung für Anthropologie und Palaoanatomie, Munich (Germany), Zoologische Staatssammlung, Munich (Germany), Zoologisches Museum, Berlin (Germany) and Natural History Museum, London (England) for the permission to access the collections. In the same way we thank Elisa Parraguéz and the Facultad de Odontología of the Universidad Mayor for their collaboration with the radiographic analysis. We also thank to Dr. Victor Manuel Bravo Cuevas and Dr. Brenda S. Ferrero for their critical revisions that helped to improve this manuscript.
REFERENCES
Alberdi, M.T., Prado J.L., 2004, Caballos fósiles de América del Sur. Una historia de tres millones de años: Olavarría, Universidad Nacional del Centro de la Provincia de Buenos Aires, Facultad de Ciencias Sociales, Investigaciones Arqueológicas y Paleontológicas del Cuaternario Pampeano (INCUAPA), Serie Monográfica, 3, 269 pp. [ Links ]
Alberdi, M.T., Prado, J.L., López, P., Labarca, R., Martínez, I., 2007, Hippidion saldiasi Roth, 1899 (Mammalia, Perissodactyla) en el Pleistoceno tardío de Calama, norte de Chile: Revista Chilena de Historia Natural, 80, 157–171. [ Links ]
Ameghino, F., 1889, Contribución al conocimiento de los mamíferos fósiles de la Republica Argentina: Actas Academia Nacional de Ciencias Córdoba, 6, 1–1027. [ Links ]
Ameghino, F., 1891, Mamíferos y aves fósiles argentinas. Especies nuevas, adiciones y correcciones: Revista Argentina de Historia Natural, 1, 240–259. [ Links ]
Bonacic, C., 2008, Ecología de la vicuña y su ordenación (online): Santiago, Ecología Info 27, actualización: 04 de agosto de 2008, <http://www.ecologia.info/vicugna.htm>, consulta: 01 de septiembre de 2009. [ Links ]
Boule, M., Thevenin, A., 1920, Mammiferes fossiles de Tarija. Mission scientifique G. de Créqui–Montfort et E. Sénéchal de la Grange: Paris, Imprimerie Nationale Édit, 256 pp. [ Links ]
Cabrera, A., 1932, Sobre los Camélidos fósiles y actuales de la América austral: Revista Museo de La Plata, 33, 89–117. [ Links ]
Cardozo, A., 1974, Auquénidos: La Paz, Editorial Centenario, 230 pp. [ Links ]
Cartajena, I., 2003, Los conjuntos arqueofaunísticos del Arcaico Temprano en la Puna de Atacama, Norte de Chile: Berlín, Alemania, Freie Universitat Berlin, tesis doctoral, 230 pp. [ Links ]
Cartajena, I., Núñez, L., Grosjean, M., 2007, Camelid domestication in the western slope of the Puna de Atacama, Northern Chile: Anthropozoologica, 42(2), 155–173. [ Links ]
Chong–Díaz, D., Jensen, A., 2004, Informe geológico del sitio de hallazgo de restos de vertebrados en el sector urbano de la ciudad de Calama, Sector de Kamac Mayu: Antofagasta, Chile, Informe de Impacto Ambiental, 5 pp. [ Links ]
Cione, A.L., Tonni, E.P., 1995. Chronostratigraphy and "Land–mammal ages" in the Cenozoic of southern South America: principles, practices, and the "Uquian" problem: Journal of Paleontology, 69, 135–159. [ Links ]
Cuvier, G., 1800, Lecond d' anatomie comparée: Paris, Baudouin, t. 1: xxxi, 521 pp.; t. 2: xvi, 697 pp. [ Links ]
De la Torre, M.U., 1981, Determinación de la edad de la vicuña según características dentales y desgaste dentario: Perú, Universidad Nacional San Luis Gonzaga de Ica, 40 pp. [ Links ]
Driesch, A. von den, 1999, A guide to measurement of animal bones from archaeological sites, Peabody Museum Bulletins 1: Massachusetts, Harvard University Press, 148 pp. [ Links ]
Ferrero, B. 2005, El registro fósil de los camélidos (Artiodactyla: Camelidae) en el Pleistoceno de la provincia de Entre Ríos, su comparación con el de la Región Pampeana y Uruguay: Ameghiniana, 42(4), 28. [ Links ]
Ferrero, B.S., 2006, Análisis discriminante de camélidos fósiles y vivientes (Artiodactyla: Camelidae) de la República Argentina ¿Una herramienta para su sistemática?, in IX Congreso Argentino de Paleontología y Bioestratigrafía, Córdoba, Argentina, Resúmenes, p. 108. [ Links ]
Franklin, W., 1983, Constrasting socioecologies of South America's wild camelids: the vicuña and the guanaco, in Eisenberg, J. y Kleiman, F. (eds.), Advances in the Study of Animal Behaviour: Kansas, American Society of Mammalogists, Special Publication, 7, 573–629. [ Links ]
Gelfo, J., López, P., Martínez, I., 2008, Una manada de Macrauchenia (Mammalia, Litopterna) del Pleistoceno tardío de Kamac Mayu, Calama II Región, Chile (resumen), in 3er Congreso Latinoamericano de Paleontología de Vertebrados, Neuquén, Argentina: Universidad Nacional del Comahue, Resúmenes, p. 18. [ Links ]
Gervais, P., 1867, Sur une nouvelle collection d'ossementes fossiles de Mammifères recueillé par M. Fr. Seguin dans la Conféderation Argentine: Comptes rendus des séances de l' Académie des Sciences, Paris, 65, 279–282. [ Links ]
Gervais, H., Ameghino, F., 1880, Los mamíferos Fósiles de la América Meridional/Les mammifères fossiles de l'Amérique du Sud: Paris, F. Savy, 225 pp. [ Links ]
Gray, J.E., 1821, On the natural arrangement of vertebrose animals: London Medical Repossitory, Monthly Journal and Review, 15(1), 296–310. [ Links ]
Guérin, C., Faure, M., 1999, Palaeolama (Hemiauchenia) niedae nov. sp., nouveau camelidae du nordeste brésilien et sa place parmi les lamini d'Amérique du Sud: Geobios, 32(4), 620–659. [ Links ]
Herre, W., 1952, Studien ubre die tilden und domestizierten Tylopoden Südamerikas: Der Zoologische Garten (Neue Folge), 19(2/4), 70–98. [ Links ]
Lichteinstein, G., Vilá, B., 2002, Vicuña use by Andean Comunities: an overview: Mountain Research and Development, 23(2), 198–2002. [ Links ]
López–Aranguren, D.J., 1930, Camélidos fósiles argentinos: Anales de la Sociedad Científica Argentina, 109, 15–39, 97–126. [ Links ]
López, P., Labarca. R., 2005, Macrauchenia (Litopterna), Hippidion (Perissodactyla), Camelidae y Edentata en Calama (II Región): Comentarios Taxonómicos y Tafonómicos: Santiago, Chile, Museo Nacional de Historia Natural, Noticiario Mensual, 355, 7–10. [ Links ]
López, P., Jackson, D., Jackson, D., 2005, El género Palaeolama P. Gervais, 1867 (Artiodactyla, Camelidae): una evaluación taxonómica regional en el extremo meridional del semiárido de Chile (IV Región): Santiago, Chile, Boletín de Museo Nacional de Historia Natural, 54, 129–140. [ Links ]
Marín, J., Zapata, B., González, B., Bonacic, C., Wheeler, J., Casey, C., Bruford, M., Palma, E., Poulin, E., Alliende, M., Spotorno, A., 2007, Sistemática, taxonomía y domesticación de alpacas y llamas: nueva evidencia cromosómica y molecular: Revista Chilena de Historia Natural, 80(2), 121–140. [ Links ]
Massone, M., Prieto, A., 2004, Evaluación de la modalidad cultural Fell 1 en Magallanes: Chungara, 36, 303–315. [ Links ]
May, G., Hartley, A., Chong Díaz, G., Stuart, F., Turner, P., Kape, S., 2005, Eocene to Pleistocene lithostratigraphy, chronostratigraphy and tectono–sedimentary evolution of the Calama basin, Northern Chile: Revista Geológica de Chile, 32(1), 33–58. [ Links ]
Menegaz, A.N., 2000, Los camélidos y cérvidos del Cuaternario del sector Bonaerense de la Región Pampeana, La Plata, Argentina: Universidad Nacional de La Plata, tesis doctoral, 240 pp. [ Links ]
Menegaz, A., 2001, Los camélidos y cérvidos fósiles del sector bonaerense de la región Pampeana durante el Cuaternario: Universidad Nacional de La Plata, Facultad de Ciencias Naturales y Museo La Plata, doctoral thesis, 206 pp. [ Links ]
Menegaz, A., Ortiz–Jaureguizar, E., 1995, Los artiodáctilos, in Alberdi, M.T., Leone, G., Tonni, E. (eds.), Evolución biológica y climática pampeana durante los últimos cinco millones de años. Un ensayo de correlación con el Mediterráneo occidental: Madrid, Consejo Superior de Investigaciones Científicas, Museo Nacional de Ciencias Naturales, Monografías, 12, 311–337. [ Links ]
Menegaz, A., Salemme, M., Ortiz–Jaureguizar, E., 1988, Una propuesta de sistematización de los caracteres morfométricos de los metapodios y las falanges de Camelidae, in Ratto, N., Haber, A. (eds.), De Procesos, Contextos y Otros Huesos. Análisis de vertebrados e invertebrados en sitios arqueológicos: Universidad de Buenos Aires, Facultad de Filosofía y Letras, Instituto de Ciencias Antropológicas, 53–64. [ Links ]
Menegaz, A., Goin, F., Ortiz Jaureguizar, E., 1989, Análisis morfológico y morfométrico multivariado de los representantes fósiles y vivientes del género Lama (Artiodactyla, Camelidae). Sus implicancias sistemáticas, biogeográficas, ecológicas y biocronológicas: Ameghiniana, 26(3–4), 153–172. [ Links ]
Mengoni–Goñalons, G., Yacobaccio, H., 2006, The domestication of south american camelids. A view from the South–Central Andes, in Zedar M. A., Bradley. G., Emshwiller E., Smith B. D. (eds.), Documenting domestication. New genetic and archaeological paradigms: Los Angeles, University of California Press, 228–244. [ Links ]
Molina, J.I., 1782, Saggio sulla storia naturale del Chili: Bologna, Stamperia di S. Tommaso d'Aquino, 367 pp. [ Links ]
Mones, A., 1986, Paleovertebrata sudamericana. Catálogo sistemático de los vertebrados fósiles de América del Sur. Parte I: Courier Forschungsinstitut Senckenberg, 82, 1–625. [ Links ]
Naranjo, J.A., Paskoff, R.P., 1981, Estratigrafía de los depósitos Cenozoicos de la región de Chiu–Chiu–Calama, Desierto de Atacama: Revista Geológica de Chile, 13–14, 79–85. [ Links ]
Noriega, J.I., Ferrero, B.S., 2007, La paleontología de vertebrados en el Cuaternario de la provincia de Entre Ríos (Argentina): Estado actual y perspectivas, in Workshop de Quaternário Do RS "Integrando conhecimentos": Canoas, Brasil, Resúmenes, p. 51–52. [ Links ]
Owen, R., 1838, Fossil Mammalia, in Darwin, C. (ed.), The zoology of the voyage of H. M. S. Beagle, under the command of Capitain Fitzroy, during the years 1832–1836: London, Smith Elder and Co., 40 pp., 7 pl. [ Links ]
Owen, R., 1848, The archetype and homologies of vertebrate skeleton: London, Van Voorst, 203 pp. [ Links ]
Prieto, A., Canto, J., 1997, Presencia de un lamoide atípico en Cueva Lago Sofía 4 (Ultima Esperanza) y Tres Arroyos (Tierra del Fuego) Región de Magallanes, Chile: Anales del Instituto de La Patagonia, 25, 147–150. [ Links ]
Ribeiro, A.M., Scherer, C.S., Pitana, V.G., 2007, Mamíferos do Pleistoceno do Rio Grande do Sul, Brasil: Estado Atual do Conhecimento, in Workshop de Quaternário Do RS "Integrando conhecimentos",: Canoas, Brasil, Resúmenes, p. 25–28. [ Links ]
Roth, S., 1899, El mamífero misterioso de la Patagonia Grypotherium domesticum. II. Descripción de los restos encontrados en la Caverna de Última Esperanza: Revista del Museo de La Plata, 9, 421–453. [ Links ]
Scherer, C.S., Ferigolo, J., Ribeiro, A.M., 2007, Contribution to the knowledge of Hemiauchenia paradoxa (Artiodactyla, Camelidae) from the Pleistocene of southern Brazil: Revista Brasileira de Paleontología, 10(1), 35–52. [ Links ]
Tauber, A., 1999, Hallazgos de una vicuña en el Pleistoceno de la Provincia de Córdoba, República Argentina: Ameghiniana, 36(1), 55–62. [ Links ]
Torres, H. 1992, South American camelids: an action plan for their conservation. IUCN/SSC South American Camelids Specialist Group: 58 p. Gland, Switzerland. [ Links ]
Ubilla, M., 2004, Mammalian biostratigraphy of Pleistocene fluvial deposits in northern Uruguay, South America: Proceedings of the Geologist's Association, 115, 347–357. [ Links ]
Ubilla, M., Perea D., Goso Aguilar, C., Lorenzo, N., 2004, Late Pleistocene vertebrates from northern Uruguay: tools for biostratigraphic, climatic and environmental reconstruction: Quaternary International, 114,129–142. [ Links ]
Ubilla, M., Perea, D., Rinderknecht, A., Corona, A., 2007, Mamíferos del Cuaternario de Uruguay. Bioestratigrafía y Paleoambientes, in Workshop de Quaternário Do RS "Integrando conhecimentos". Canoas, Brasil, Resúmenes, p. 45–47. [ Links ]
Webb, S.D., 2006, The great American biotic interchange: Patterns and processes: Annals of the Missouri Botanic Garden, 93(2), 245–257. [ Links ]
Weinstock, J., Shapiro, B., Prieto, A., Marín, J. C., González, B., Thomas, P., Willerslev, E., 2009, The Late Pleistocene distribution of vicuñas (Vicugna vicugna) and the "extinction" of the gracile llama (''Lama gracilis"'): New molecular data: Quaternary Science Reviews, 28(15–16), 1369–1373. [ Links ]
Wheeler, J.C., 1984, On the origin and early development of camelid pastoralism in the Andes, in Clutton–Brock, J., Grigsons, C. (eds.), Animals and Archaelogy: Oxford, BAR International Series 202, 395–410. [ Links ]
Woodburne, M.O., Cione, A.L., Tonni, E.P., 2006, Central American provincialism and Great Biotic Interchange, in Carranza–Castañeda, O., Lindsay, E.H. (eds.), Advances in late Tertiary vertebrate paleontology in Mexico and the Great American Biotic Interchange: Ciudad de México, Universidad Nacional Autónoma de México, Instituto de Geología y Centro de Geociencias, Special Publication, 4, 73–101. [ Links ]

