











 text new page (beta)
text new page (beta) Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO 
 Permalink
Permalink
INTRODUCCIÓN
De acuerdo con Nelson et al. (2016) el orden de los Carangiformes está formado por seis familias, entre las cuales se encuentra Carangidae, la cual agrupa 30 géneros, con 152 especies, de las cuales 24 se distribuyen en el Golfo de México.
En las costas mexicanas se comercializan bajo el nombre de “jurel” varias especies del género Caranx, con un promedio de pesca de 18.9 mil toneladas anualmente (SAGARPA, 2019).
El “jurel” Caranx hippos (Linnaeus, 1766) llega a medir hasta 1.2 m de longitud y pesar 25 kg. Se distribuye desde Nueva Escocia, Canadá hasta Uruguay, incluyendo el Golfo de México (Castro-Aguirre et al., 1999). De acuerdo con Arreguín-Sánchez & Arcos-Huitrón (2011) su explotación pesquera en México se encuentra en estado máximo de aprovechamiento.
La mayoría de los peces del género Caranx tienen un rol ecológico como depredadores, además forman parte de la dieta de depredadores tope como tiburones, marlines, dorados y túnidos, manteniendo el flujo de la energía en la cadena trófica (Hoffmayer & Parsons, 2003; Junior et al. 2004; Estupiñan-Montaño et al., 2009; Vázquez-Ozuna et al., 2020).
Son pocos los estudios previos sobre la anatomía de los carángidos, entre ellos destacan Kong & Iratchet (1998), quienes describen el neurocráneo de Trachurus murphyi (Nichols, 1920), Caranx caballus (Günther, 1868), Seriola lalandi (Valencinnes, 1833), Trachinotus paitensis (Cuvier, 1832), Selene peruviana (Guichenot, 1866) y Selene brevoorti (Gill, 1863), y el estudio de Kobelkowsky & Beltrán-Vinalay (2020) sobre la morfología de la cavidad visceral y la esplacnología de Selene vomer (Linnaeus, 1758).
A pesar de su importancia ecológica y económica, los estudios que detallen aspectos anatómicos relacionados con la alimentación son escasos. El análisis anatómico de la cavidad bucal de un pez puede brindar información sobre los hábitos alimenticios, puesto que participa en la captura y primer procesamiento del alimento, en donde intervienen varias estructuras relacionadas al tipo de presa como dentición, numero de branquiespinas, grado de desarrollo de ciertos músculos entre otras. Por otra parte, una de las características de la familia Carangidae es la diversidad de la forma corporal, lo que favorece la comparación de las estructuras de especies con diferentes hábitos alimenticios.
MATERIALES Y MÉTODOS
Los ejemplares de C. hippos, se colectaron mediante varias artes de pesca en un área cercana a la boca de las lagunas de Tampamachoco, Tuxpan y de laguna Grande en Vega de Alatorre, Veracruz, localidades del litoral del Golfo de México. Se fijaron en formaldehido al 10% y se conservaron en alcohol etílico al 70%. En total se procesaron 20 ejemplares adultos con un intervalo de talla de 35.7 a 64.9 cm.
Las disecciones se realizaron bajo un microscopio estereoscópico Olympus SZ6, siguiendo la técnica de disección del tracto digestivo de los teleósteos de Kobelkowsky & Figueroa (2018).
Se consideró a la cavidad bucofaríngea o cavidad orobranquial como parte del tracto digestivo, que se inicia con la abertura de la boca y termina en el borde posterior de los huesos faríngeos. Se analizaron anatómicamente y describieron los componentes de la cavidad bucofaríngea, así como también las estructuras relacionadas indirectamente con la misma, y se ordenaron como esqueleto visceral, musculatura visceral y dentición.
Con el objetivo de hacer comparaciones anatómicas de estructuras relacionadas con la alimentación se seleccionaron los siguientes carángidos: Chloroscombrus chrysurus (Linnaeus, 1766), Decapterus punctatus (Cuvier, 1829) Selene vomer (Linnaeus, 1758) y Trachinotus carolinus (Linnaeus, 1766), que se distribuyen también en el Golfo de México y que representan la diversidad de la familia Carangidae.
Las ilustraciones se realizaron mediante un tubo de dibujo acoplado a un microscopio estereoscópico Wild M3Z.
La terminología osteológica siguió el criterio de Gregory (1959) y la miológica, el de Winterbottom (1974).
RESULTADOS
Cavidad bucofaríngea. La cavidad bucofaríngea de Caranx hippos se extiende desde la boca hasta la región posterior del aparato branquial. A esta cavidad le da forma y sostén el esqueleto visceral y su correspondiente musculatura.
Posteriormente al borde de la boca se forman los componentes de la válvula oral.
El paladar está tapizado por un gran número de placas pequeñas con dientes finos; su borde posterior marca el inicio de la porción faríngea.
El piso de la cavidad bucofaríngea muestra en su porción anterior a la lengua, estructurada por el cartílago glosohial y el hueso basihial; a continuación, se observan las ramas inferiores de los arcos branquiales y sus correspondientes branquiespinas. Tanto el paladar como el piso de la cavidad bucofaríngea tienen hacia atrás continuidad con el esófago.
Esqueleto visceral (Fig. 1). Se compone en ambos lados por las siguientes series de huesos: las mandíbulas superior e inferior, la serie palatina, la suspensión mandibular, el aparato hioideo, la serie opercular y el aparato branquial. Algunos de los elementos esqueléticos se interconectan mediante cartílagos y ligamentos.
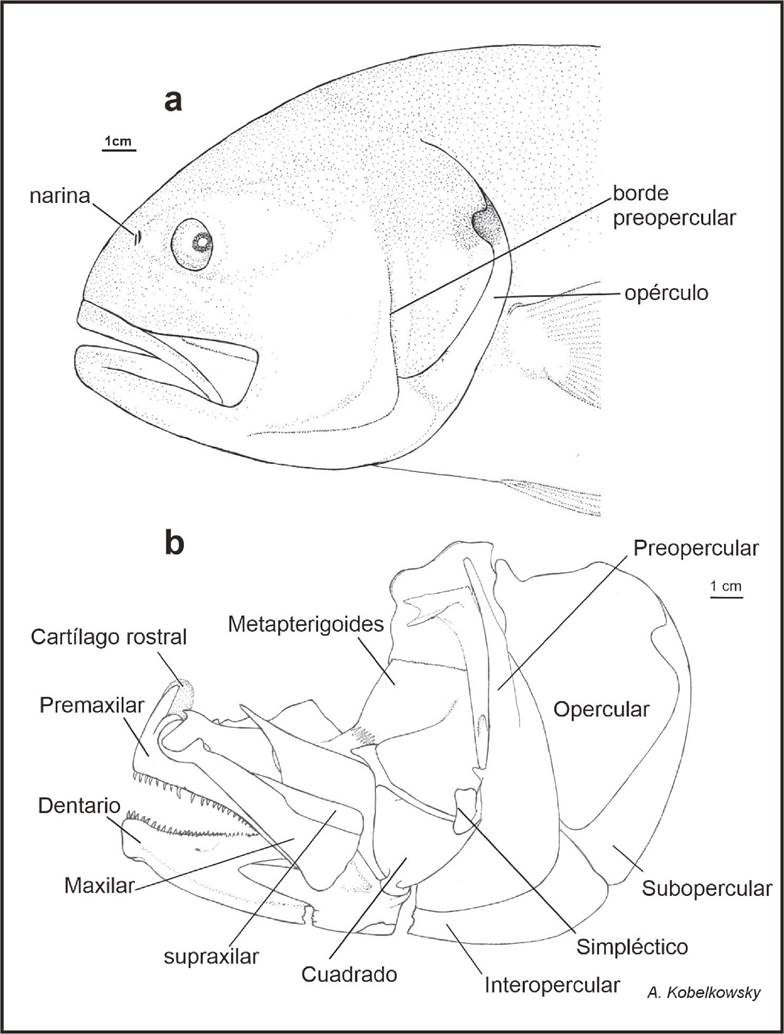
Figura 1 Región cefálica de Caranx hippos. a) Vista lateral izquierda de la región cefálica de Caranx hippos. b) Vista lateral del esqueleto visceral.
La mandíbula superior, de cada lado, está formada por el premaxilar, el maxilar y el supramaxilar. Entre los premaxilares se encuentra el cartílago rostral, de aspecto “ovoide”. La mandíbula inferior está constituida por los huesos dentario, ánguloarticular y retroarticular; en la cara interna de este último se ubica el coronomeckeliano y el cartílago de Meckel (Fig. 2).

Figura 2 Esqueleto visceral de Caranx hippos. Vista lateral izquierda. a) Arco mandibular, serie palatina, suspensión mandibular. b) Detalle de la cara interna del Arco mandibular, serie palatina, suspensión mandibular c) Aspecto del proceso anterior del ectopterigoides. d) Cresta lateral del hiomandibular de Caranx hippos. e) vista dorsal del suborbital 3.
El premaxilar desarrolla dos procesos ascendentes, articular y maxilar. El maxilar es delgado y ligeramente curvo; en su porción anterior o cabeza desarrolla un proceso articular dorsal y otro ventral. Se articula con el palatino y el premaxilar.
Entre el maxilar y el etmoides se extiende el ligamento etmo-maxilar, mientras que entre los procesos anteriores dorsales de ambos maxilares se forma el ligamento intermaxilar. El supramaxilar es alargado, comienza aproximadamente al nivel de la parte media del maxilar (Fig. 2a).
El dentario es robusto, presenta el proceso coronoides y en su superficie lateral lleva una rama de la línea lateral con varios orificios. En su borde bucal lleva dos filas de dientes cónicos y curvados hacia la cavidad oral. En su región posterior se encuentra la escotadura donde se intercala el hueso ánguloarticular.
El ánguloarticular es triangular, presenta el proceso coronoides y cerca de su extremo posterior desarrolla una foseta articular que recibe al cóndilo del cuadrado, posterior a la cual, se desarrolla un proceso corto que recibe desde el preopercular al ligamento preopérculo-ánguloarticular. En su superficie interna se observa el cartílago de Meckel y el coronomeckeliano de aspecto romboidal, bien desarrollado (Fig. 2b)
El retroarticular es un hueso pequeño, unido firmemente a la porción posteroventral del ánguloarticular. Este hueso recibe desde del interopercular al ligamento interoperculo-retroarticular.
La serie palatina está formada por el palatino, endopterigoides, metapterigoides y ectopterigoides. El palatino desarrolla un proceso anterior que se articula con el maxilar y tiene contacto con el ligamento etmo-palatino y el ligamento maxilo-palatino (Fig. 2c). El hueso más amplio es el metapterigoides, mientras que el endopterigoides es laminar y recibe parte del músculo adductor arcus palatini. El ectoterigoides es amplio, desarrolla un proceso del cual se extiende un ligamento hasta el prefrontal.
La suspensión mandibular está constituida por los huesos hiomandibular, metapterigoides, simpléctico y cuadrado.
El hiomandibular tiene tres cóndilos, el anterior se articula al hueso esfenótico, el medio al pterótico, y el tercero se articula con el opercular. En su superficie lateral externa se inserta el músculo levator arcus palatini, el cual está limitado posteriormente por un reborde vertical del mismo hiomandibular. Cabe destacar que el hiomandibular tiene contacto con la extensión del suborbital 3 mediante una cresta con dos puntas que se origina del mismo (Fig. 2d).
Asimismo, el hiomandibular constituye la suspensión del aparato hioideo, mediante el hueso interhial y un ligamento corto (interhial-hiomandibular). El metapterigoides es laminar y se opone externamente a la parte posterior del endopterigoides, manteniendo un espacio para una porción del músculo adductor arcus palatini. El simpléctico es laminar, es el hueso más pequeño de esta serie, se ubica en la escotadura posterior del cuadrado y se une a este mediante sutura. El cuadrado es el hueso más grande de la serie, su porción anterior es triangular y forma un cóndilo que se articula al hueso ánguloarticular y la posterior es rectangular la cual se adosa ampliamente al preopercular.
La serie opercular está formada por los huesos preopercular, opercular, interopercular y subopercular. La superficie interna de los huesos interoperculares y subopercular recibe al conjunto de radios branquiostegos. El preopercular es semitriangular relativamente de tamaño medio, con borde ligeramente aserrado; se acopla fuertemente con el hiomandibular y el cuadrado; lleva una rama del sistema sensorial de la línea lateral, la cual continúa hasta la mandíbula inferior. El opercular es laminar de forma trapezoidal, su borde anterior es grueso y el posterior delgado, donde se aprecia una escotadura. Se articula mediante una fosa al cóndilo posterior del hiomandibular (Fig. 2d). Desde dicha fosa se extiende horizontalmente un reborde que delimita el área de inserción de los músculos dilatador operculi y levator operculi. El interopercular es laminar de borde inferior curvo, parcialmente cubierto por el preopercular, de su extremo anterior parte un ligamento hacia el hueso ánguloarticular. El subopercular es laminar y alargado, desarrolla un proceso antero dorsal que bordea el ángulo inferior del opercular. Además, se observó la presencia de un ligamento que va del opérculo a la cresta pterótica.
El aparato hioideo está suspendido desde el extremo inferior de los hiomandibulares, está constituido por dos barras hioideas y dos huesos impares. Cada barra hioidea está integrada por el hipohial dorsal, hipohial ventral, ceratohial, epihial y 7 radios branquiostegos; los huesos impares son el basihial y el urohial (Fig. 3).

Figura 3 Cavidad bucofaríngea de Caranx hippos. a) Vista de los componentes de la cavidad bucofaríngea in situ. b) Detalle de la boca. c) Vista ventral del aparato hioideo. d) Vista lateral y posterior del urohial.
El basihial está precedido por el cartílago glosohial, formando ambos elementos a la lengua. Cada hipohial ventral recibe ventralmente a un músculo hyohyoidei abductor. Los ceratohiales son largos y se unen con los huesos vecinos por sutura; cada ceratohial sostiene los branquiostegos del 1 - 4 y a los epihilales 6 y 7, quedando el 5 entre estos dos huesos (Fig. 3a y 3b).
Los epihiales están ligeramente curvados y se articulan con los interhiales, estos son relativamente pequeños y cilíndricos, se unen al extremo inferior de los hiomandibulares.
El urohial forma en su porción anterior un proceso ascendente que recibe al músculo rectus communis. La sección transversal de su porción anterior tiene forma de “Y” invertida y recibe al músculo sternohyoideus que también tiene contacto con el cleitrum (Fig. 3c y 3d).
Los radios branquiostegos, aumentan gradualmente su amplitud de adelante hacia atrás, mientras que los tres primeros son delgados y se fijan en unas concavidades que están sobre la cara interna del ceratohial, los restantes son planos y amplios, se unen a la cara externa del ceratohial y del epihial de la siguiente manera: el 4 está sobre el ceratohial, el 5 sobre la unión del ceratohial y el ephial y el 6 y 7 en el ephial.
El aparato branquial está suspendido desde la región ótica del neurocráneo por los faringobranquiales 1 y está constituido por cinco arcos branquiales, formados a su vez por los huesos impares basibranquiales 1-4, y los huesos pares hipobranquiales 1-3, ceratobranquiales 1-5, epibranquiales 1-4 y faringobranquiales 1-4.
Los basibranquiales son elementos esbeltos, de los cuales el más largo es el 3 y el más corto es el 1. El basibranquial 4 se curva hacia abajo y sirve como punto de inserción de los músculos obliquui ventrales III. Posterior al basibranquial 4 se localiza el cartílago basibranquial.
Los hipobranquiales son cortos y muestran su superficie ventral cóncava, de la cual se originan los músculos obliquui ventrales I, II y III.
Los ceratobranquiales son los elementos más largos del aparato branquial, los ceratobranquiales 1- 4 son acanalados ventralmente para dar paso a las arterias branquiales aferentes; los ceratobranquiales 4 muestran una concavidad ventral para la inserción del músculo transversus ventralis IV. Los ceratobranquiales 5 están modificados en los huesos “faríngeos inferiores”, cada uno con un proceso anterior y otro posterior relativamente largos. En su superficie dorsal llevan dientes, y en la ventral tienen una concavidad donde se inserta el músculo transversus ventralis V.
Cada epibranquial forma un proceso dorsal que recibe a un músculo levator externi; sus extremos superiores son de cartílago. Los cartílagos correspondientes a los epibranquiales 2 - 4 se unen a la superficie dorsal de los huesos faringobranquiales, mientras que del proceso dorsal del epibranquial 1 se une al faringobranquial 2 mediante el cartílago interarcual.
Los faringobranquiales superiores son de forma irregular y en total son 4; de ellos, solo del 2 al 4 llevan dientes que varían en tamaño. El 1 es corto, cilíndrico y constituye la suspensión del aparato branquial, desde la región ótica del neurocráneo. El 2 es rectangular, el 3 es de tendencia triangular, lleva los dientes más robustos y por último el 4 es ovalado. Los faríngeos inferiores son triangulares y llevan dientes que varían en longitud y diámetro.
Musculatura visceral (Fig. 4). El músculo adductor mandibulae, está formado por las secciones A1, A2, A3 y Aw. La sección A1 se origina cerca del borde del preopercular; cerca de su extremo anterior tiene contacto con el hueso lacrimal y mediante un tendón se inserta al maxilar y la superficie externa del articular. La sección A2 se ubica en la región lateral de la mejilla; se inserta mediante un tendón al proceso coronoides del dentario (Fig. 4a). Entre las fibras musculares de esta sección pasa otro tendón que también se inserta en el dentario. La sección A3 se ubica por debajo de la sección A2 y sobre el cuadrado. La sección Aw se ubica en la cara interna del dentario.

Figura 4 Musculatura visceral de Caranx hippos. Vista lateral izquierda. a) Relaciones topográficas del adductor mandibuale. b) Músculo Levator arcus palatini. c)Músculo Dilatador operculi y el Ligamento opercular-pterótico.
Los músculos que permiten el movimiento del paladar y la suspensión mandibular se originan en el neurocráneo y son el adductor arcus palatini que se inserta en el endoterigoides y metapterigoides, el levator arcus palatini que está dividido en dos secciones debido a la cresta que se desarrolla en hiomandibular (Fig. 4b) y el adductor hyomandibulae que se inserta en la superficie interna del mismo hueso.
La musculatura hioidea está constituida por los músculos protactor hyoidei y stenohyodei. El protactor hyoidei se origina de la superficie externa del aparato hioideo y se inserta mediante un ligamento a la parte interna de la sínfisis mandibular. El sternohyoideus se origina en el cleitrum y se inserta en ambas caras del urohial. Cabe mencionar que el músculo dilatador operculi se extiende hasta la cresta esfenótica (Fig. 4c).
Los músculos de los arcos branquiales se pueden dividir en dos grupos, dorsales y ventrales. Los dorsales se componen de cuatro levatores externi, que se originan en el pterótico y aumentan su longitud del primero al último. El levator posterior se origina de la cápsula ótica y se inserta en el epibranquial 4. El músculo retractor dorsalis está formado por varios paquetes que se originan de las partes laterales y ventrales de las vértebras 2 - 4 y se insertan en los faringobranquiales.
Los músculos ventrales se componen de pharyngoclavicularis externus e internus, transversi ventrales anterior y posterior, y el rectus comunis. El músculo pharyngoclavicularis internus, se origina de la porción media de la superficie anterior del cleitrum y se insertan en el hueso faríngeo inferior. El músculo pharyngoclavicularis externus se origina cerca del borde de la porción inferior del cleitrum y se inserta en el faríngeo inferior. El músculo transversus ventralis IV pasa del ceratobranquial 4 de un lado al otro. El transversi ventralis V pasa del ceratobranquial 5 de un lado al otro. El músculo rectus communis se origina en el ceratobranquial 5 y se inserta en el borde superior del urohial.
Dentición. (Figs. 5, 6 y 7). En C. hippos la dentición está presente en el premaxilar, dentario, aparato branquial, lengua, vómer, palatino, aparato hioideo. Además, existen dientes muy finos en la superficie de la cavidad oral.
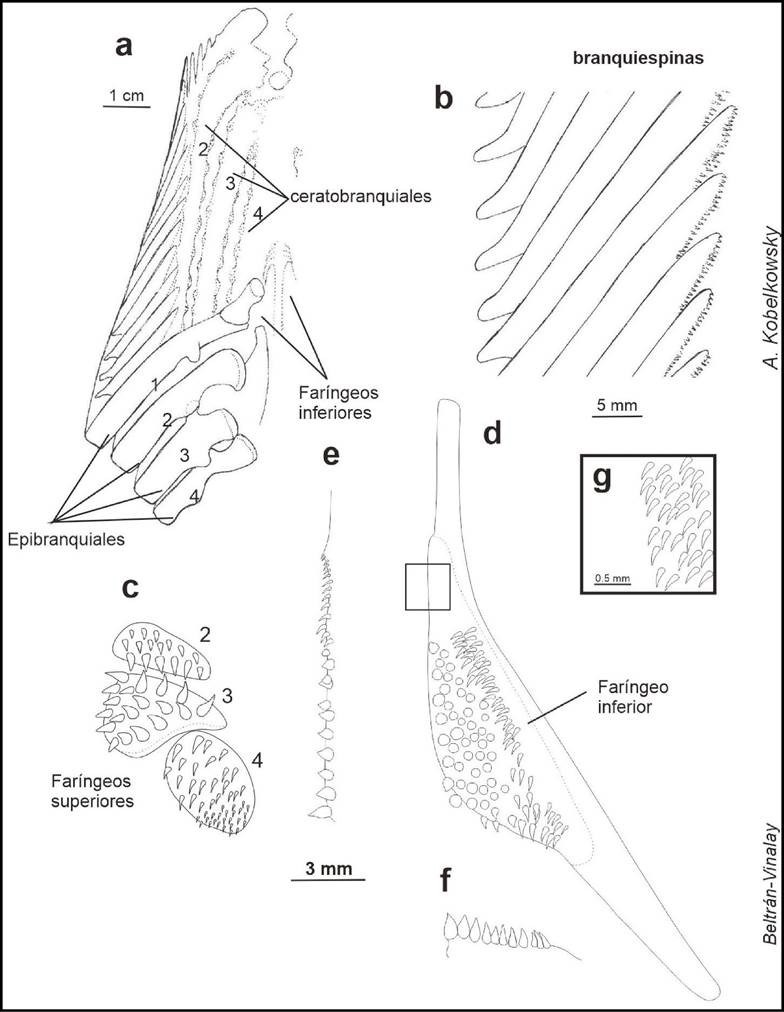
Figura 5 Aparato branquial de Caranx hippos. a) Vista dorsal. b) Aspecto dorsal de las branquiespinas del borde externo del primer arco branquial. c) Vista dorsal de huesos faríngeos superiores. d) Vista dorsal del hueso faríngeo inferior. e) Detalle del borde lateral del hueso faríngeo inferior. f) Detalle de borde posterior del hueso faríngeo inferior. g) Detalle de los dientes de la región anterior de hueso faríngeo inferior.
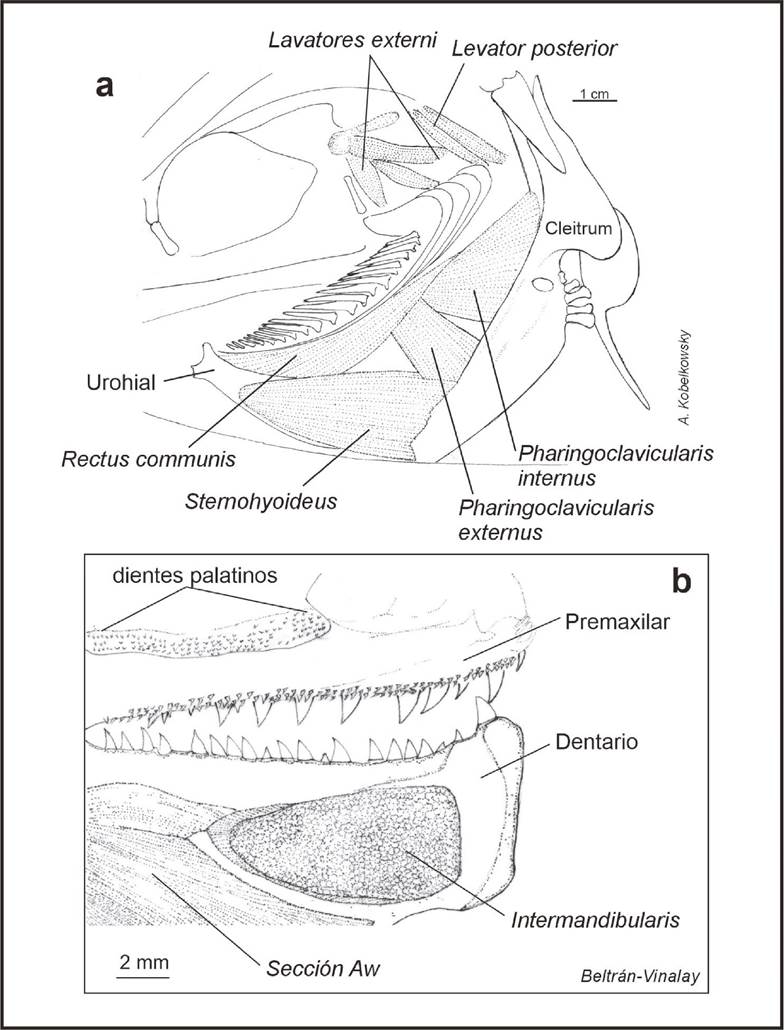
Figura 6 Arcos branquiales y boca de Caranx hippos. Vista lateral izquierda. a) Vista lateral izquierda del aparato branquial, mostrando los principales músculos que lo relacionan con otros elementos esqueléticos. b) Detalle de los dientes platinos.

Figura 7 Dentición de Caranx hippos. a) Localización de los dientes vomerinos. b) detalle de los dientes vomerinos. c) vista dorsal del premaxilar. d) vista lateral del premaxilar. e) Vista de la cara interna del premaxilar. f) Vista dorsal del dentario. g) Vista lateral del dentario. h) Vista de la cara interna del dentario.
Los dientes del premaxilar están arreglados en una hilera de elementos grandes y cuatro hileras de pequeños, que van disminuyendo gradualmente de tamaño a medida que se acercan a la porción posterior.
El dentario lleva una hilera de dientes medianos y cerca de la sínfisis se inserta una segunda con solo dos grandes.
El aparato branquial lleva dientes en las branquiespinas y en los huesos faríngeos. En las branquiespinas del primer arco los dientes son pequeños y se ubican en su borde interno. Los dientes de las branquiespinas restantes son un poco más grandes que los anteriores. También, están presentes numerosas placas pequeñas con dientes muy finos; dichas placas son de forma irregular y cubren el epitelio de los basibranquiales 1, 2 y 3.
Todos los dientes de los faringobranquiales superiores e inferiores son cónicos y curvos en dirección al esófago, su tamaño y grosor es variable.
Los dientes del faringobranquial 2 son en general robustos y van aumentando de tamaño de la porción anterior a la posterior. En el faringobranquial 3 los dientes también son robustos, pero su tamaño disminuye del borde exterior al borde interior. Los del faringobranquial 4 son delgados, agudos y disminuyen de tamaño de la región anterior a la región posterior.
Los dientes del borde exterior de cada hueso faríngeo inferior (ceratobranquial 5) son notablemente pequeños, pero aumentan de tamaño a medida que se acercan al borde interior, en donde los dientes son robustos y ligeramente romos.
También están presentes dientes pequeños en la lengua, el vómer, el palatino y unos muy finos sobre placas pequeñas distribuidas sobre el epitelio de la cavidad oral. Además, se observaron placas ovaladas con dientes en la cara interna del aparato hioideo y en el envés de los opérculos.
Aspectos comparativos de la morfología de la cavidad bucofaríngea de C. hippos con otras especies de Carángidos. (Tabla 1).
Tabla 1 Comparación de los elementos de la cavidad bucofaríngea de Caranx hippos, Chlorocombrus chrysurus, Decapterus punctatus, Selene vomer y Trachinotus carolinus.
| Caranx hippos (Linnaeus, 1766) | Chloroscombrus chrysurus (Linnaeus, 1766) | Decapterus punctatus ( Cuvier, 1829) | Selene vomer (Linnaeus, 1758) | Trachinotus carolinus (Linnaeus, 1766) | |
|---|---|---|---|---|---|
| Branquiespinas del primer arco | 18 | 35 | 47 | 30 | 10 |
| Esqueleto visceral | completo | completo | completo | sin endopterigoides | completo |
| Proceso del ectoterigoides | * | * | * | * | x |
| Cresta del hiomandibular | * | x | X | x | x |
| Cresta esfenótica | * | * | * | * | * |
| D. Premaxilar | Grandes y pequeños | pequeños | pequeños | muy pequeños | x |
| D. Dentario | Grandes y medianos | pequeños | pequeños | muy pequeños | x |
| D. Vomer | pequeños | pequeños | pequeños | pequeños | x |
| D. Palatino | pequeños | pequeños | pequeños | muy pequeños y romos | x |
| D. Lengua | pequeños | pequeños | delgados y medianos | muy pequeños y romos | x |
| D. Ectoterigoides | x | pequeños | X | x | x |
| D. Endoterigoides | x | pequeños | X | x | x |
| D. Faringobranquiales 2 | robustos | robustos | medianos | robustos | fusionados |
| D. Faringobranquiales 3 | robustos | redondeados | redondeados | robustos | fusionados |
| D. Faringobranquiales 4 | delgados y agudos | delgados | delgados | robustos | fusionados |
| D. Faringeobranquiales inferiores | varían en diámetro | varían en diámetro | varían en diámetro | varían en diámetro | fusionados |
| Placas dentadas sobre el aparato braquial | * | * | * | muy abundantes | x |
| Placas dentadas sobre el epitelio oral | * | x | X | x | x |
| M. Adductor mandibulae | bien desarrollado | normal | normal | poco desarrollado | desarrollado |
| M. Adductor arcuspalatini | normal | poco desarrollado | normal | poco desarrollado | muy desarrollado |
| M. Levatores externi e interni | normal | poco desarrollado | normal | normal | muy desarrollados |
| M. Retractor dorsalis | normal | normal | normal | normal | muy desarrollado |
| M. Dilatator operculi extendido hasta la Cef | * | * | * | * | * |
| Branquiespinas del primer arco | 18 | 35 | 47 | 30 | 10 |
| Esqueleto visceral | completo | completo | completo | sin endopterigoides | completo |
| Proceso del ectoterigoides | * | * | * | * | x |
| Cresta del hiomandibular | * | x | X | x | x |
| Cresta esfenótica | * | * | * | * | * |
| D. Premaxilar | Grandes y pequeños | pequeños | pequeños | muy pequeños | x |
| D. Dentario | Grandes y medianos | pequeños | pequeños | muy pequeños | x |
| D. Vomer | pequeños | pequeños | pequeños | pequeños | x |
| D. Palatino | pequeños | pequeños | pequeños | muy pequeños y romos | x |
| D. Lengua | pequeños | pequeños | delgados y medianos | muy pequeños y romos | x |
| D. Ectoterigoides | x | pequeños | X | x | x |
| D. Endoterigoides | x | pequeños | X | x | x |
| D. Faringobranquiales 2 | robustos | robustos | medianos | robustos | fusionados |
| D. Faringobranquiales 3 | robustos | redondeados | redondeados | robustos | fusionados |
| D. Faringobranquiales 4 | delgados y agudos | delgados | delgados | robustos | fusionados |
| D. Faringeobranquiales inferiores | varían en diámetro | varían en diámetro | varían en diámetro | varían en diámetro | fusionados |
| Placas dentadas sobre el aparato braquial | * | * | * | muy abundantes | x |
| Placas dentadas sobre el epitelio oral | * | x | X | x | x |
| M. Adductor mandibulae | bien desarrollado | normal | normal | poco desarrollado | desarrollado |
| M. Adductor arcuspalatini | normal | poco desarrollado | normal | poco desarrollado | muy desarrollado |
| M. Levatores externi e interni | normal | poco desarrollado | normal | normal | muy desarrollados |
| M. Retractor dorsalis | normal | normal | normal | normal | muy desarrollado |
| M. Dilatator operculi extendido hasta la Cef | * | * | * | * | * |
Los * = presencia, x= ausencia, D = dentición, M = músculo y Cef = cresta esfenótica
Se reconoce un patrón morfológico de los elementos que conforman la cavidad bucofaríngea en los carángidos, que concuerda con lo descrito anteriormente para otros teleósteos superiores, sin embargo, se reconocen algunas variaciones anatómicas.
Comparando el esqueleto visceral se encontró que en S. vomer el endopterigoides está ausente. En C. hippos, C. chrysurus, D. punctatus y S. vomer el ectopterigoides desarrolla un largo proceso del cual se extiende el ligamento ectopterigoides-prefrontal; dichas estructuras no se presentan en T. carolineus. Solo en C. hippos el hiomandibular desarrolla la cresta lateral y la extensión posterior del suborbital 3. Por otra parte, en S. vomer el hiomandibular es notablemente estrecho; en las demás especies no se observa ninguna de estas dos variaciones. En T. carolinus se destaca que el premaxilar y el dentario son cortos, frágiles y sin dientes. En contraste, el premaxilar y dentario de C. hippos es robusto y presenta dentición bien desarrollada.
Las variaciones más notables de la musculatura visceral son las siguientes: en C. hippos el adductormandibulae está bien desarrollado, contrario a lo observado en S. vomer. Los músculos levatores externi, levatores interni y levator posterioris, así como los retractores dorsales, se encuentran notablemente desarrollados en T. carolinus. En C.hippos, el músculo levator arcus palatini está dividido en dos secciones debido al desarrollo de la cresta del hiomandibular, que solo se presenta en esta especie.
La dentición está presente en el premaxilar, dentario, vómer, el palatino, la lengua y el aparato branquial en todas las especies, excepto en C. chrysurus que además lleva dientes en el ectoterigoides y endoterigoides, mientras que en T. carolinus solo está presente en las branquiespinas y en los huesos faríngobranquiales, formando placas gruesas.
DISCUSIÓN
La organización de la dentición, esqueleto y musculatura visceral de Caranx hippos corresponde al patrón descrito en otros perciformes carnívoros por Gregory (1959), Kobelkowsky (2007) y Kobelkowsky & Pantoja-Escobar (2007, 2009), y que es congruente con la estrategia trófica de los teleósteos superiores, que de acuerdo con Vandewalle et al. (2000) y Gerking (1994), implica que la presa es succionada por una corriente de agua generada por el abrupto descenso del piso de la cavidad bucofaringea, posteriormente la presa recorre dicha cavidad, donde es procesada por la dentición del aparato branquial, principalmente por los faringobranquiales superiores e inferiores. Lo anterior sugiere que el aparato branquial está relacionado directamente con el tipo de presa y en particular los dientes faríngeos. Esto corresponde a lo observado en C. hippos y en las especies analizadas con fines comparativos. El jurel C. hippos presenta las características típicas de un pez que se alimenta de otros teleósteos: dentición mandibular y faríngea bien desarrollada, así como también el músculo adductor mandibulae, bajo número de branquiespinas. Estas características también están descritas por Kobelkowsky & Pantoja-Escobar (2009) y por Winterbottom (1974), para Lutjanus griseus, un pez carnívoro depredador. Algunos autores como Fagade & Olaniyan (1973) y Saloman & Naughton (1984), analizaron la dieta de C. hippos y concluyen que se alimenta principalmente de peces y peneidos, lo cual de acuerdo con su anatomía es la dieta esperada.
En contraste, Trachinotus carolinus no presenta dentición mandibular y la faríngea está fusionada en gruesas placas, los músculos levatores externi, interni, posterior; rectocomunis y pharingoclaviculares internus y externus están notablemente desarrollados, lo cual capacita a esta especie para el consumo de presas duras; Armitage & Alevizon (1980) y Denadai et al. (2013) determinaron que la dieta de T. carolinus se basa en el consumo de bivalvos como Dorax variabilis, Tellina spp. Tivela mactroides.
La dentición mandibular de las demás especies no está muy desarrollada, la faríngea consta principalmente de dientes pequeños, redondeados, delgados y relativamente robustos, lo cual corresponde a peces que se alimentan de zooplancton y otras presas pequeñas, que concuerda con lo reportado para C. chrysurus por Chaves & Umbria (2003) y Cunha et al. (2000); para D. punctatus, fue reportado por Stanton (1987) y Donaldson & Clavijo (1994); y para S. vomer, por Buchheister & Latour (2015). De acuerdo con la Tabla 1 podemos enlistar algunas características de los peces que se alimentan principalmente de zooplacton, dentición mandibular poco desarrollada, alto número de branquiespinas y dientes faríngeos pequeños.
De las especies antes mencionadas, sobresale Selene vomer, en el cual el músculo adductor mandibulae está poco desarrollado en comparación con C. hippos, C. chrysurus y D. punctatus, pero muy similar al del lenguado Syacium papillosum descrito por Kobelkowsky & Rojas-Ruiz (2017), el cual también es un pez muy comprimido.
De las variaciones morfológicas encontradas destacan el proceso del ectoterigoides y el ligamento ectoterigoides-prefrontal, el desarrollo de la cresta del hiomandibular que solo se presenta en C. hippos, y el desarrollo de la cresta esfenótica. A pesar que Kong & Iratchet (1998) describen el neurocráneo de varias especies de carángidos, no mencionan la cresta, solo hacen referencia al hueso esfenótico; y por último, la notable extensión del músculo dilatador operculi. Estas dos últimas podrían ser características de la familia Carangidae, anteriormente no descritas.
De acuerdo con lo anterior, se concluye que C. hippos es una especie morfológicamente bien adaptada a una dieta de tipo carnívora principalmente piscívora, pues presenta las características típicas de un pez depredador, tales como capacidad de expansión de la cavidad bucofaríngea, dentición mandibular bien desarrollada, bajo número de branquiespinas, dientes faríngeos cónicos y curvos (lo que impide que la presa escape), así como un músculo adductor mandibulae bien desarrollado. Además, una gran cantidad de placas pequeñas con dientes finos en el epitelio de la cavidad oral, lo que sugiere un posible aumento en la eficiencia del descamado de la presa.

