











 text new page (beta)
text new page (beta) Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO 
 Permalink
PermalinkINTRODUCCIÓN
Los crustáceos decápodos comprenden una gran variedad de especies que se explotan comercialmente, como los camarones, los langostinos, los cangrejos y las langostas. Dentro de estas últimas, las langostas australianas, Cherax quadricarinatus (von Martens, 1868); C. destructor Clark, 1936 y C. tenuimanus (Smith, 1912) son especies de agua dulce que han alcanzado un gran éxito en el mercado internacional en los últimos años. El cultivo de Cherax quadricarinatus (von Martens, 1868) se ha desarrollado ampliamente, tanto en Australia como en diversos países de regiones tropicales y subtropicales. Esto se debe a sus características biológicas que hacen factible su producción, ya que su desarrollo es directo al no presentar período larvario, alcanza tasas de crecimiento altas, con pesos de 60 a 100 g (talla comercial) en individuos de seis a ocho meses de edad, con un intervalo térmico de tolerancia amplio (20 - 30ºC) en comparación con los camarones peneidos, como Penaeus vannamei (Boone, 1931) que alcanza de 15 a 25 g en 4-5 meses, a temperaturas de 25 a 30°C (FAO, 2009). Además es resistente a las enfermedades y al manejo. Acepta el alimento balanceado desde el inicio de la alimentación exógena, las hembras presentan una fecundidad alta con varios eventos reproductivos por año. Tiene una gran aceptación por parte de los consumidores, lo que causa que alcance un atractivo valor comercial en el mercado (Jones & Ruscoe, 1996, Castillo-Corella et al., 2002, Campaña-Torres et al., 2005).
En condiciones de cultivo, las tasas de crecimiento y de mortalidad son los factores que más importancia tienen en la producción. Estas variables cambian con la edad de los individuos y con la calidad del hábitat (Thorp & Covich, 2001). Se ha observado que C. quadricarinatus utiliza refugios en condiciones naturales, para protegerse de la depredación, tanto inter como intra-específica. Este comportamiento de la especie se ha aplicado en su cultivo, en la fase de engorda con buenos resultados (Jones & Ruscoe, 2001; Manor et al., 2002; Barki et al., 2006; Arzola-González et al., 2012).
La fase temprana del período juvenil es una de las fases de desarrollo con mayor vulnerabilidad de esta especie (Gallo-García et al., 2012; Calvo et al., 2013). Sin embargo, existe escasa información de las condiciones necesarias para optimizar la producción y la sobrevivencia de juveniles en la fase de pre-engorda, por lo que el objetivo de este trabajo fue evaluar el efecto de diferentes densidades de refugios, para determinar la condición más adecuada que permita maximizar el crecimiento y la sobrevivencia durante el desarrollo temprano.
MATERIALES Y MÉTODOS
Los juveniles de C. quadricarinatus se obtuvieron por reproducción natural, de reproductores mantenidos en cautiverio. Se realizó un experimento completamente aleatorio, donde se evaluó el efecto del número de refugios, sobre el crecimiento y la supervivencia de los juveniles. Los niveles utilizados fueron: sin refugios (tratamiento 1:T1), un refugio por individuo (tratamiento 2: T2) y dos refugios por individuo (tratamiento 3: T3), cada tratamiento con dos réplicas. Los refugios consistieron en cilindros de PVC de 10 cm de longitud y 2.5 cm de diámetro. Se colocaron 30 juveniles en estadio III, de 15 días después de la eclosión (dde) e inicio de alimentación exógena, con un peso inicial de 114.3 ± 3.5 mg, en contenedores con un volumen de 1000 L y una superficie del fondo de 1 m2 (30 juveniles/m-2). Se les suministró alimento balanceado para camarón (48% de proteína, Silver Cup™) .La ración diaria fue del 10% de su peso húmedo. Todos los tratamientos se mantuvieron a una temperatura de 25°C. Se realizaron recambios de agua del 50% cada semana, la concentración de amonio se mantuvo en 0.05 mgL-1 y el oxígeno en 5 mgL-1. Se determinó el peso (P) y la longitud total (LT) cada dos semanas, así como la sobrevivencia, en cada tratamiento durante 92 días.
Análisis de datos. Los datos de peso y longitud se transformaron al logaritmo natural para normalizarlos. Se comprobó la homogeneidad de varianzas entre los tratamientos mediante la prueba Bartlett. Para comprobar que no hubiera diferencias de los pesos y longitudes iniciales entre réplicas ni entre tratamientos se aplicó un análisis de varianza. El crecimiento en LT y en P se ajustó a modelos de regresión lineal y el crecimiento relativo del peso con respecto a la longitud a un modelo potencial (P = aLTb) y se probó la significancia de los coeficientes de regresión (ANOVA < 0.05). Los modelos de crecimiento se compararon, con el método de comparación de pendientes de Tukey-Kramer (a = 0.05) (Sokal & Rohlf, 1981).
Los valores finales se analizaron mediante un modelo lineal generalizado (GLM) para evaluar el efecto de la edad y el número de refugios sobre el peso y la longitud y el contenedor como factor anidado, dentro del tratamiento del número de refugios. Finalmente, se realizó la prueba de Kolmogorov-Smirnov (a = 0.05) para comparar las distribuciones de frecuencias de peso y LT y una prueba de chi-cuadrada (R Core Team, 2016) para comparar la sobrevivencia entre los tratamientos.
RESULTADOS
Se comprobó la homocedasticidad y la normalidad de los pesos y longitudes. Así mismo, los valores iniciales fueron semejantes, tanto entre las réplicas como entre los tratamientos (ANOVA, p > 0.05). Al final del experimento, los juveniles de las réplicas de cada tratamiento, presentaron pesos y longitudes semejantes (ANOVA, p>0.05) por lo que los datos de las réplicas de cada tratamiento, se agruparon para ajustar los modelos de crecimiento (P y LT) y los modelos de peso-longitud.
Crecimiento. Las curvas de crecimiento en peso y en longitud total, se ajustaron a un modelo exponencial (y = a expxb) (ANOVA, p < 0.05), en todos los tratamientos: P1= 0.13e 0.033t, r2 = 0.74;P2 = 0.17e 0.042t, r2 = 0.87;P3 = 0.15e0.039t, r2 = 0.85 (Fig. 1) y LT1 = 18.1 e 0.011t, r2 = 0.73;LT2 = 19.6e 0.014t, r2 = 0.87;LT3 = 18.7e 0.013t, r2 = 0.84 (Fig. 2). La edad influyó significativamente en P (t = 25.243, p<0.0001) y en LT (t = 41.95, p<0.0001),así como también se presentó una interacción entre la edad y la proporción de refugios por individuo (tratamientos) sobre P (t =10.94, p<0.0001) y LT (t=9.813, p<0.0001).

Figura 1 Crecimiento en peso de C. quadricarinatus. T1 (∆): P1 = 0.13e0.033t, r2 = 0.74; T2 (●): P2 = 0.017e0.042t, r2 = 0.87; T3 (■): P3 = 0.15e0.039t, r2 = 0.85.
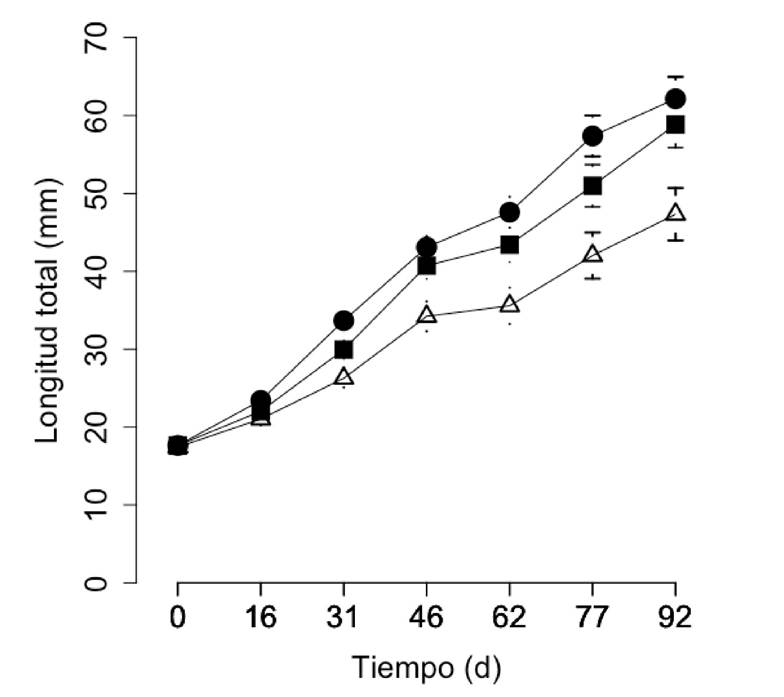
Figura 2 Crecimiento en LT de C. quadricarinatus. T1 (∆): LT1 = 18.1e0.011t, r2 = 0.73; T2 (●): LT2 = 19.6e0.014t, r2 = 0.87; T3 (■): LT3 = 18.7e0.013t, r2 = 0.84.
Después de 92 días, los promedios finales, tanto de P como de LT, fueron significativamente mayores en los tratamiento con refugios (T2: 6.17 g, 62.1 mm y T3: 5.18 g, 58.8 mm) comparados con los controles sin refugios (T1: 2.89 g, 47.3 mm) (ANOVA, F(2, 145)= 28.52 (peso), p < 0.05; F(2, 145) = 26.79 (LT), p < 0.05, N = 148).); las tasas de crecimiento mantuvieron la misma tendencia: T1 = 0.030 g d-1, T2 = 0.065 g d-1 y T3 = 0.055 g d-1 (Tukey-Kramer, p < 0.05, N = 148). T2 y T3 fueron semejantes en los valores promedio y en las tasas de crecimiento de P y LT (Tabla 1).
Tabla 1 Peso, longitud total (LT) y sobrevivencia finales de C. quadricarinatus por tratamiento.
| Tratamiento | P (g) | LT (mm) | Sobrevivencia (%) |
| Sin refugios (T1) | 2.9 ± 1.9 | 47.3 ± 11.59 | 80 ± 0.04 |
| 1 refugio/individuo (T2) | 6.15 ± 2.8 | 62.1 ± 10.08 | 85 ± 0.07 |
| 2 refugios/individuo (T3) | 5.19 ± 2.4 | 58.8 ± 10.3 | 81.5 ± 0.02 |
En cambio, la distribución de frecuencias, tanto de P como de LT, fueron diferentes en cada tratamiento, donde en T1se presentaron los individuos más pequeños y en T2 los más grandes (Kolmogorov-Smirnov, p < 0.001) (Figs. 3 y 4).
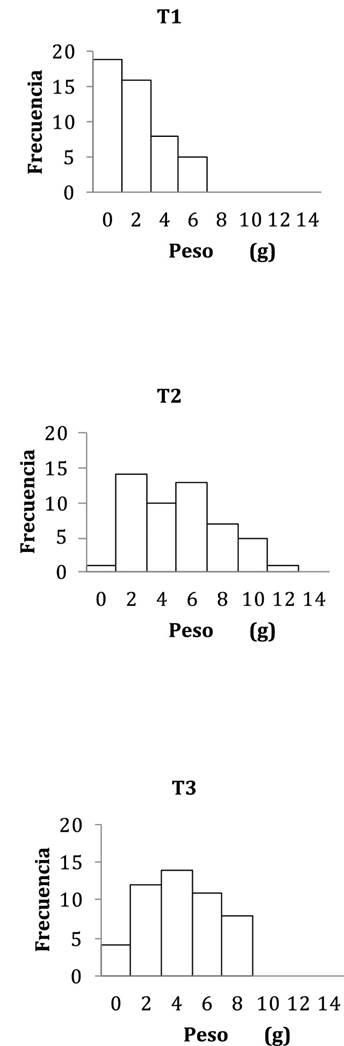
Figura 3 Frecuencias de las clases de peso (g) por tratamiento. T1: sin refugios, T2: 1 refugio/ind., T3: 2 refugios/ind.

Figura 4 Frecuencias de las clases de talla (mm) por tratamiento. T1: sin refugios, T2: 1 refugio/ind., T3: 2 refugios/ind.
Sin embargo, la sobrevivencia fue semejante en todos los tratamientos, T1: 80%, T2: 85% y T3: 81.7% (chi-cuadrada, p > 0.05) (Tabla 1)al final del período de prueba.
Crecimiento relativo Peso- Longitud. El crecimiento relativo de P con respecto a la LT, fue del tipo alométrico positivo (b > 3,ts, p < 0.05), en todos tratamientos. La comparación de las tasas de crecimiento demostró que no hay diferencias de esta variable, entre los tratamientos (Tukey-Kramer, p > 0.05) T1: P = 1x10-5 TL3.26, r2 = 0.996; T2: P = 3x10-5 TL3.09, r2 = 0.998; T3: P = 2x10-5TL3.11, r2 = 0.994 (Fig. 5).

DISCUSIÓN
La heterogeneidad del hábitat es uno de los factores principales que influyen en la calidad del mismo y que afectan la reproducción, la alimentación, el crecimiento y la sobrevivencia, en los crustáceos decápodos y en peces (Able, 1999; Phelan et al., 2000). En condiciones naturales, la vegetación sumergida es un factor muy importante en los sitios de crianza de los decápodos de agua dulce, ya que provee protección contra la depredación y alimento por la fauna asociada a ella (Wilson et al., 1990a, 1990b).
El número de refugios y de individuos, son factores importantes a considerar en el cultivo de juveniles tempranos de C. quadricarinatus. Esta especie es territorial y utiliza refugios como protección, la ausencia de éstos provoca canibalismo, mayor competencia por el espacio y disminuye la tasa de crecimiento, lo que implica un efecto negativo del hábitat. En este estudio, la presencia de refugios favorecieron el crecimiento de los juveniles ya que disminuyen la competencia por el espacio. La ausencia de refugios redujo el crecimiento de los juveniles de manera significativa, en contraste con los tratamientos con refugios, donde se obtuvieron las mayores tasas de crecimiento, si bien, se observó que un refugio por individuo es suficiente en esta etapa del desarrollo, ya que no hubo diferencias entre las tasas de crecimiento ni en la sobrevivencia de los tratamientos T2 y T3. Se ha demostrado que las especies de Cherax, bajo condiciones de cultivo, presentan un crecimiento y sobrevivencia denso-dependientes (Barki & Karplus, 2004). Tanto los refugios como la densidad de los individuos son factores importantes a considerar. En juveniles tempranos de C. quadricarinatus se han obtenido pesos finales bajos (0.32- 0.62 g) en individuos de 30 - 40 días de edad, con densidades iniciales muy altas (100 - 200 org/m2) aún con la presencia de refugios (Parnes & Sagui, 2002; Gallo-García et al., 2012; García-Ulloa & Pinzón-López, 2012; Calvo et al., 2013). Las densidades altas causan diferencias de tamaño, aún entre individuos de la misma edad y peso iniciales, debido a que los individuos dominantes (alfa) limitan el alimento a los individuos pequeños y menos competitivos, de manera que éstos alcanzan tamaños menores (enanos). Este efecto fue notable en el tratamiento sin refugios, ya que los individuos tuvieron pesos y longitudes, marcadamente menores a los obtenidos en los otros tratamientos y esta diferencia se incrementó a partir de los 46 días, a pesar de que se utilizó la misma densidad, los individuos eran de la misma edad y con pesos y longitudes iniciales iguales. Se ha atribuido esta variación en el crecimiento, al control social en el crecimiento en los decápodos territoriales, como los langostinos (Macrobrachium rosenbergii De Man 1879) (Karplus, 2005), las langostas (Homarus americanus Milne Edwards 1837) (Cobb et al., 1982) y las langostas de agua dulce (C. destructor, Verhoef & Austin, 1999; C. quadricarinatus, Karplus & Barki 2004; Barki et al., 2006).
La variación del peso con respecto a la longitud, describe el cambio de forma asociado al incremento en tamaño y es una manera indirecta de analizar el grado de robustez en los decápodos. La relación alométrica entre el peso y la longitud, es la más frecuente en los decápodos. En el presente trabajo, se obtuvo una relación alométrica positiva en todos los tratamientos, sin diferencias entre ellos, a pesar de que los individuos que se mantuvieron sin refugios alcanzaron un tamaño menor, lo que indica que la ausencia de refugios no afecta la robustez de las langostas. Resultados semejantes se han obtenido en juveniles cultivados más avanzados de esta especie (Jones, 1995). La relación alométrica del peso con respecto a la longitud en los decápodos, puede variar dependiendo de la ontogenia, el sexo y de las especies. En hembras de C. quadricarinatus y C. destructor el crecimiento es alométrico negativo (Austin et al., 1997). Juveniles de Penaeus monodon Fabricius 1798, tuvieron un crecimiento alométrico positivo mayor que los pre-adultos (Primavera et al., 1998). Por el contrario, Procambarus acutusacutus Girard 1852, en juveniles avanzados y los machos con respecto a las hembras, los individuos son más pesados con el incremento en la longitud (Mazlum et al., 2007). En Procambarus alleni Faxon 1884, el crecimiento es alométrico positivo, sin diferencias entre machos y hembras (Acosta & Perry, 2000).
Finalmente, en este trabajo se demostró que la presencia de refugios, si bien no incrementó la supervivencia, se obtuvo una tasa de crecimiento más alta e individuos más grandes, en comparación con el tratamiento sin refugios. Jones (1995) y Karplus et al. (1995) obtuvieron resultados semejantes con esta especie y Austin et al. (1997) y Verhoef & Austin (1999) con C. destructor.

