










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkHidrobiológica
Print version ISSN 0188-8897
Hidrobiológica vol.17 suppl.1 Ciudad de México Jul. 2007
Cambios tisulares en el ostión Crassostrea virginica por exposición y depuración al cadmio
Tissue changes of the oyster Crassostrea virginica due to cadmium exposition and depuration
Xochitl Guzmán-García1, Alejandro Martínez-López1, Lizan Rodríguez-Medina1, Humberto González-Márquez2 y Alfonso Vázquez-Botello3
1 Alumna del Doctorado en Biología Experimental. Laboratorio de Ecotoxicología, Departamento de Hidrobiología. UAM-I. Av. San Rafael Atlixco 186, Col. Vicentina Iztapalapa, C.P. 09340. México, D. F. Tel. (55) 5804 64 74 Fax (55) 5804 4738. xgg@xanum.uam.mx
2 Laboratorio de Expresión Génica. Departamento de Ciencias de la Salud. UAM-I.
3 Laboratorio de Contaminación Marina, ICMyL, UNAM.
Recibido: 16 de octubre de 2005
Aceptado: 10 de julio de 2006
Resumen
Los niveles de cadmio se han incrementado en el Golfo de México, esto representa un riesgo potencial para la sobrevivencia del ostión Crassostrea virginica y sus consumidores, por lo que es importante evaluar los efectos del cadmio a través de los cambios histopatológicos derivados de la exposición a este metal y durante su depuración. Se analizaron ostiones expuestos a 100 µg/l de cadmio por 72 h y depurados por 24, 48, 72, 96, 120, 144, 168, 192, 216 y 230 h. Las muestras de ostión fueron incluidas en parafina, cortadas en microtomo y teñidas con la técnica de Hematoxilina-Eosina. El análisis histopatológico del esófago, intestino, glándula digestiva, branquia y manto mostró lesiones inflamatorias y la activación celular que incluye, células cafés y hemocitos como un mecanismo de desintoxicación. Los tejidos conjuntivos cercanos a esófago, intestino y glándula digestiva presentaron más células cafés y hemocitos que la branquia y el manto. Los cambios histopatológicos no fueron reversibles en 230 h, a pesar de que hay recuperación de los epitelios de la glándula digestiva. Este trabajo recomienda la evaluación histopatológica en la glándula digestiva para determinar el estado fisiológico del ostión y tomar medidas preventivas en la comercialización y cultivo de sus poblaciones naturales.
Palabras clave: Ostión, Crassostrea virginica, cadmio, histopatológico.
Abstract
Cadmium levels had increased in the Gulf of Mexico, this represents a potential risk for the survival of oyster Crassostrea virginica and for those who eat them, so it is important to evaluate the effects of cadmium trough histopathological changes derived from the exposition to this metal and during its depuration. Oysters exposed to 100 µg/l of cadmium for 72 h and depurated by 24, 48, 72, 96, 120, 144, 168, 192, 216, and 230 h were analyzed. Samples of oysters were included in paraffin, cutted in a microtome and stained following the Hematoxilin-Eosin technique. Histopathological analysis of oesophagus, intestine, digestive diverticulum, gills and mantle, showed inflammatory lesions and cell activation, including brown cells and haemocytes as a detoxification mechanism. The conjunctive tissue next to oesophagus, intestine and digestive diverticulum presented more brown cells and haemocytes than gills and mantle. Histopathological damage was not reversible in 230 h, although there was a recovery in epitheliums of digestive diverticulum. This work recommends the histopathological evaluation of digestive diverticulum to determine the physiological state of oysters and to take preventive measures in the commercialization and culture of their natural populations.
Key works: Oyster, Crassostrea virginica, cadmium, histopathological.
Introducción
El uso industrial de los metales ha provocado que la movilización antropogénica, en algunos casos, sea mayor que la natural, siendo el aporte antropogénico el que más daño ecológico causa (Gutiérrez-Galindo et al., 1991). Los estudios de la contaminación por metales pesados en el medio ambiente marino tienen el propósito de proteger los ecosistemas marinos costeros, en general, y la salud humana, en particular. Con este fin, se ha propuesto el empleo de organismos centinelas como organismos biológicos indicadores de la contaminación por metales pesados en el medio marino.
Los moluscos bivalvos, tales como las ostras, juegan un papel ecológico importante en los sistemas acuáticos. La filtración e ingestión que realiza de materia particulada suspendida (algas y sedimentos) de la columna de agua remueve contaminantes asociados con la materia suspendida. Las ostras son organismos sésiles incapaces de evadir condiciones desfavorables, se alimentan por filtración y se conoce que bioacumulan contaminantes como el cadmio (Cd), permitiendo el diseño de experimentos para estimar la cinética de bioacumulación y transplante en el laboratorio (Cruz-Rodríguez & Fu-Lin, 2002; Frías-Espericueta et al., 1999).
La bioacumulación de metales en bivalvos depende de las propiedades físico-químicas de los elementos en cuestión, así como de las necesidades metabólicas de los organismos y la disponibilidad de éstos en la columna de agua y/o alimentos (Frías-Espericueta et al., 1999).
El Cd es un elemento estable que no se degrada en el ambiente, es considerado como uno de los más tóxicos para un gran número de organismos marinos, siendo acumulado por los bivalvos en proporciones que en algunos casos, pueden constituir un serio peligro si son destinados a la alimentación humana (Gutiérrez-Galindo et al., 1991; Establier et al., 1978; Fisher et al., 2000).
En México la legislación establece 0.1 mg/l de Cd como Límite Máximo Permitido (LMP) en las descargas de aguas residuales en aguas y bienes nacionales (Diario Oficial de la Federación, 1997). Sin embargo, Villanueva & Botello (1998) han reportado concentraciones de Cd mayores al LMP en varias lagunas del Golfo de México, así como concentraciones en el ostión Crassostrea virginica (Gmelin, 1791) que superan los 2 µg/gps, recomendados para consumo humano.
El ostión Crassostrea virginica corresponde a una de las especies estuarinas explotadas con mayor intensidad en las costas mexicanas del Golfo de México, habita en aguas salobres, lagunas y esteros, tiene una amplia distribución y es una especie comercialmente valiosa (Rogers & García-Cubas, 1981; Frías-Espericueta et al., 1999). En el siglo pasado ocurrió la mayor declinación de poblaciones naturales por una variedad de causas incluyendo la sobreexplotación, el daño del hábitat, la degradación de la calidad de agua y el incremento en la susceptibilidad a enfermedades (Matthew et al., 2002).
Engel (1999), sugiere que el ostión C. virginica utiliza diferentes estrategias metabólicas en la captación o acumulación de metales. Diversos estudios señalan la bioconcentración de Cd en estudios de laboratorio (Zaroogian & Morris, 1981); sin embargo, la cuantificación de los contaminantes subestima el estado fisiológico de los organismos, por lo que el objetivo de este trabajo fue analizar los cambios histopatológicos durante la fase de desintoxicación en el ostión Crassostrea virginica expuesto a concentraciones subletales de Cd.
Materiales y métodos
Se procesaron 66 muestras del ostión Crassostrea virginica, procedentes de un bioensayo con exposición subletal a 100 µg/l de Cd y sus respectivos testigos. El bioensayo se realizó en dispositivos experimentales de 40 l. La fase de intoxicación tuvo una duración de 72 h y la fase de desintoxicación fue de 240 h con muestreos de 3 organismos a las 24, 48, 72, 96, 120, 144, 168, 192, 216 y 230 h. Los ostiones fueron fijados con formol al 10 %, posteriormente se deshidrataron, aclararon e infiltraron en un procesador de tejidos automático modelo LEICA TP 1020. La inclusión del tejido se realizó en parafina con puntos de fusión de 49 - 51º y de 58 - 60 °C utilizando el centro de inclusión modelo LEICA EG 1140 H y LEICA EG 1140 C para la obtención de los bloques. Posteriormente se hicieron 3 cortes seriados de 5-7 µm de grosor con un micrótomo modelo HM 315. Los cortes fueron teñidos con la técnica de Hematoxilina-Eosina (H-E) y revisados en microscopio óptico. Se trabajó el esófago, el intestino, la glándula digestiva, las branquias y el manto. Para evaluar la fase de desintoxicación se utilizó como parámetro de clasificación el número de las células cafés (llamadas así, porque contienen vesículas con tinción café en el citoplasma; Zaroogian & Yevich, 1994) y hemocitos presentes en los tejidos. Se determinó el número de células cafés realizando un conteo en 6 campos a 100X, por corte y en sus réplicas (3). Un total de 18 campos fueron revisados para cada tejido y se obtuvo en ellos el promedio de células cafés.
La densidad de hemocitos se utilizó para la evaluación cualitativa, ésta fue clasificada en 4 categorías:
1 = hemocitos en tejido conjuntivo periférico al órgano.
2 = hemocitos con infiltración baja en el órgano.
3 = hemocitos con infiltración moderada en el órgano.
4 = hemocitos con infiltración elevada, no se distinguen límites entre tejidos.
Se graficaron el número de células cafés y la categoría de hemocitos presentes en el tejido conjuntivo adyacente a esófago, intestino, glándula digestiva, branquia y manto desde la intoxicación (72 h) hasta el final del periodo de desintoxicación (230 h).
Resultados
El análisis histopatológico de ejemplares de Crassostrea virginica expuestos a Cd, evidenció respuestas inflamatorias, degeneración de cilios en el intestino, daño en los epitelios de los túbulos de la glándula digestiva, basofilia e hialinización. En el esófago de los ostiones se observó tinción basófila en el epitelio de los palpos e incluso algunas secreciones posiblemente mucosas (Fig.1A). En esta misma estructura, el tejido conjuntivo presentó pigmentos cafés llamados en estos organismos células cafés durante la fase de depuración (Fig.1a). En el intestino de los organismos intoxicados se observó la degeneración de los cilios en la parte luminar del epitelio que lo recubre (Fig.1B). Los organismos de la fase de depuración presentaron edemas, ruptura del epitelio y presencia de células cafés en el tejido conjuntivo que rodea el intestino (Fig.1b).
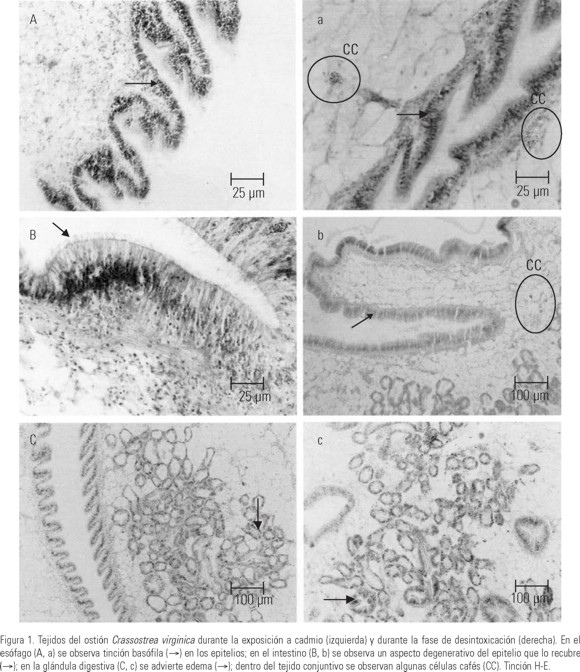
Los epitelios que revisten la luz de los túbulos de la glándula digestiva prácticamente desaparecieron y sufrieron deformación (Fig.1C). Sin embargo, al final del periodo de depuración se observó la recuperación de la apariencia ciliada (Fig. 1c).
Las laminillas branquiales presentaron en la porción apical tinción basófila (Fig.2A) muy probablemente por la presencia de hemocitos como una reacción de defensa ante la presencia de Cd (Fig.2a). En el manto la respuesta fue similar, el lóbulo sensorial externo presentó tinción basófila, en tanto que en la fase de depuración el manto mostró aspecto hialino (Fig. 2B, b).

Algunas respuestas celulares inducidas se manifestaron a través de la presencia de células cafés y hemocitos (Fig. 3A y 3B respectivamente). En el tejido conjuntivo de los organismos expuestos a Cd, se observaron las células cafés, en tonalidades de marrón a café claro con la tinción H-E (Fig. 3 A, a). Células cafés con diferente tamaño (1 a 5 µm) se observaron también en los canales de hemolinfa. Las células cafés de mayor tamaño presentaron forma de vesícula y tuvieron apariencia granular en el interior. Éstas se cuantificaron en el esófago, intestino, túbulos de la glándula digestiva y el manto (Fig. 4).

En el tejido conjuntivo, entre el intestino y los túbulos de la glándula digestiva, se observaron hasta 35% más células cafés que en el esófago y el manto. El mayor número de células cafés se registraron a las 96 h de depuración en el intestino (Fig. 4B), en tanto que la mayor incidencia de estas células en los túbulos de la glándula digestiva se presentó a las 144 h (Fig. 4C). Posteriormente a estos plazos prevaleció una tendencia hacia la disminución del número de células cafés. En las últimas horas de la fase de desintoxicación el número de células cafés fue similar en los organismos testigo, sin embargo la apariencia granulada de las vesículas cafés en organismos intoxicados, sugiere la captación de material, es decir, de Cd.
Los ostiones presentaron una respuesta hemocítica, como se muestra en la Fig. 3 (B,b) paralela al incremento en el número de células cafés, como respuesta a la inflamación aguda. Los hemocitos, posiblemente granulocitos por su actividad protectora, se presentaron en esófago, intestino, glándula digestiva, branquia y manto. La densidad de hemocitos fue mayor en el tejido cercano al intestino y las branquias con respecto al esófago, túbulos de la glándula digestiva y el manto. Este mecanismo de defensa fue evidente desde las 24 horas de la fase de desintoxicación y disminuyó su densidad a las 230 h (Fig. 5).
Discusión
Las alteraciones en la anatomía y en la función de los organismos analizados se relacionaron con diferentes reacciones histopatológicas, entre las que destaca la inflamación. La inflamación es una reacción de los tejidos a diferentes formas de lesión y entraña respuestas vasculares, neurológicas, humorales y celulares en el foco que sufre daño (Rubin & Farber, 1990).
La célula expuesta a una agresión subletal persistente tiene un repertorio limitado de respuestas (como son la producción de células cafés y los hemocitos en el caso del ostión), cuya expresión se interpreta como evidencia de lesión celular. Zaroogian & Yevich (1994) han señalado que la actividad de las células cafés de C. virginica se relaciona con el proceso de incorporación, acumulación, desintoxicación y degradación de material extraño de la hemolinfa y que aparecen conjuntamente con amebocitos, los cuales fagocitan partículas y envuelven solutos, mientras que las células cafés embeben solutos eficientemente como un mecanismo de defensa.
Los resultados de este trabajo señalan que se presentó un incremento de células cafés y hemocitos en los ostiones intoxicados con respecto a los testigos. Esto evidencia que, tanto las células cafés como los hemocitos participan en la desintoxicación de Cd en los tejidos de C. virginica.
El intestino y los túbulos de la glándula digestiva forman parte de una de las principales vías de incorporación de tóxicos, por lo que, se considera que la activación de células cafés podría ser un mecanismo primario de defensa en estos órganos. En la región del manto y en la branquia se observaron pocas células cafés, posiblemente por ser un órgano externo más relacionado con el agua circundante se favorece la eliminación de estas células.
En los organismos desintoxicados, la reparación se asumió como un proceso en donde las células destruidas fueron substituidas por nuevas células. Si el estrés desaparece a tiempo o, si la célula es capaz de soportar la agresión en la lesión celular, se restablece la integridad estructural y funcional completa (Rubin & Farber, 1990). La acción de las células cafés constituye un mecanismo importante de protección ante la presencia del Cd. Zaroogian y Yevich (1994) han demostrado que, en las llamadas células cafés existen algunas más pequeñas que son más activas en la acumulación de metal que las células cafés grandes, las células pequeñas contienen vesículas con lisosomas llamados "jóvenes" y las células cafés más grandes presentan forma de vesículas con grandes lisosomas "viejos", menos activos ó cuerpos residuales.
La activación de los hemocitos fue más notoria en los túbulos de la glándula digestiva, así como en el intestino. Éstos actúan como un mecanismo de defensa en los órganos internos análogo al de las células cafés. Otros autores han reportado un incremento en la densidad de hemocitos en el ostión C. virginica por exposición a Cd, estos hemocitos pueden captar iones de Cd de la hemolinfa que rodea los tejidos y transportarlo al lecho epitelial para ser eliminado del organismo (Fisher et al., 2000).
La presencia de hemocitos fue una respuesta claramente activada evidente en el esófago, la branquia, el manto y la glándula digestiva. Existen reportes que señalan el bloqueo de los canales de calcio por la acumulación de Cd en la branquia de C. virginica (Roesijadi & Unger 1993), esto implicaría que existe un límite fisiológico de captación de iones, que puede generar lesión y respuesta humoral. Posiblemente por ello la branquia permaneció con una densidad de hemocitos constante a lo largo de las observaciones. Otros autores han demostrado que la concentración de los metales traza en C. virginica puede ser grande, pero la toxicidad llega a ser neutralizada por el contenido de metales en granulocitos (Okazaki & Panietz, 1981; Zaroogian & Morris, 1981).
Se consideró que existen diferencias histopatológicas importantes entre los tejidos internos (el intestino y la glándula digestiva) y externos (el esófago, las branquias y el manto) de estos moluscos. Es posible que en los tejidos internos los iones de Cd se unan estrechamente a los grupos sulfhidrilo en láminas del mucus, en tanto que los tejidos externos se encuentran con el agua marina por lo que la depuración pasiva puede verse favorecida. La respuesta en los tejidos internos sugiere que su principal función es el almacén y, posteriormente la eliminación de metales traza a través de las células cafés, los hemocitos y el mucus. Es necesario ampliar este tipo de estudios con la evaluación de inmunomarcadores: actividad fagocítica, producción de especies oxígeno-reactivas (radicales libres) que provocan daño celular, densidad de hemocitos y la relación del número y estabilidad lisosomal de las células cafés (Matthew et al., 2002) de manera que se establezcan parámetros útiles en la evaluación de la inmunocompetencia celular-mediada en ostiones.
El estado histopatológico de los túbulos de la glándula digestiva y del intestino resultante de la exposición a Cd se asoció, de manera más clara, con los mecanismos de desintoxicación en el ostión Crassostrea virginica por lo que la evaluación en la glándula digestiva es recomendable en los estudios de la fisiología y monitoreo ambiental. La activación de mecanismos de desintoxicación a través de los macrófagos llamados células cafés y las células de la hemolinfa, disminuyó paulatinamente con el tiempo de depuración. Las células cafés se mantuvieron presentes hasta el final del periodo de depuración en forma de vesículas granuladas, constatando la unión del metal. Diferentes mecanismos de desintoxicación contribuyeron a la reparación de los tejidos epiteliales de los túbulos de la glándula digestiva, sin embargo los cambios histopatológicos no fueron reversibles completamente en 230 h de depuración, por lo que se sugiere en futuros estudios prolongar este tiempo para la evaluación de la recuperación de daños.
El uso de la herramienta histopatológica permitió realizar el diagnóstico de daños y evidenció cambios importantes que facilitan la selección de inmunomarcadores en tejidos específicos, así como, plazos de experimentación adecuados para la interpretación de las respuestas biológicas. La evaluación de estas respuestas es necesaria para la evaluación del impacto de enfermedades en las pesquerías y puede ser usado como medida preventiva de riesgo de la salud pública por ingestión de bivalvos contaminados.
Bibliografía
CRUZ-RODRÍGUEZ, A.L. & E.C. FU-LIN. 2002. Heat-shock protein (HSP70) response in the eastern oyster, Crassostrea virginica exposed to PAHs sorbed to suspended artificial clay particles and to suspended field contaminated sediments. Aquatic Toxicology 60: 157-168. [ Links ]
DIARIO OFICIAL DE LA FEDERACIÓN. 1997. Norma Oficial Mexicana NOM-001-SEMARNAT-1996 que establece los Limites Máximos Permitidos de contaminantes en las descargas de aguas residuales en aguas y bienes nacionales. Secretaría de Medio Ambiente Recursos Naturales y Pesca. México, D. F., enero 6: 68-86. [ Links ]
ENGEL, W.E. 1999. Accumulation and cytosolic partitioning of metals in the American oyster Crassostrea virginica. Marine Environmental Research 47: 89-102. [ Links ]
ESTABLIER, R., M. GUTIÉRREZ & A. RODRÍGUEZ. 1978. Acumulación de cadmio en el músculo y hepatopáncreas del langostino (Penaeus kerathurus) y alteraciones histopatológicas producidas. Investigación Pesquera 42 (2): 299-304. [ Links ]
FISHER, W.S., L.M. OLIVER, J.T. WINSTEAD & E.R. LONG. 2000. A survey of oysters Crassostrea virginica from Tampa Bay, Florida: associations of internal defense measurements with contaminants burdens. Aquatic Toxicology 51: 115-138. [ Links ]
FRÍAS-ESPERICUETA, G.M., A.M. ORTIZ-ARELLANO, I.J. OSUNA-LÓPEZ & J.A. RONSON-PAULIN. 1999. Heavy metals in the rock oyster Crassostrea iridescens (Filibranchia: Ostreidae) from Mazatlan, Sinaloa, Mexico. Revista Biología Tropical 47(4): 843-849. [ Links ]
GUTIÉRREZ-GALINDO, E.A., M.G. FLORES, G.P. GARCÍA, J.A. CELAYA & G.J. ARMENTEROS. 1991. Metales pesados en tejidos y en biodepósitos sedimentarios del ostión Crassostrea gigas de la zona de cultivo de bahía San Quintín, Baja California, México. Investigación Marina. CICIMAR. 6 (1): 175-186. [ Links ]
MATTHEW, J.J., A.H. RINGWWOOD, E.R. LACY, A.J. LEWITUS, J.W. KEMPTON, P. S. GROSS, G.W. WARR & R.W. CHAPMAN. 2002. Potential indicators of stress response identified by expressed sequence tag analysis of hemocytes and embryos from the American Oyster, Crassostrea virginica. Marine Biotechnology 4: 81-93. [ Links ]
OKAZAKI, K.R. & H.M PANIETZ. 1981. Depuration of twelve trace metals in tissues of the oysters Crassostrea gigas and C. virginica. Marine Biology 63: 113-120. [ Links ]
ROGERS, P. & A. GARCÍA-CUBAS. 1981. Evolución gonádica a nivel histológico del ostión Crassostrea virginica (Gmelin, 1971) del sistema fluvio lagunar Atasta-Pom, laguna de Términos, Campeche, México. Anales del Instituto de Ciencias Del Mar y Limnología. Universidad Nacional Autónoma de México 8(1): 21-42. [ Links ]
ROESIJADI, G. & M. E. UNGER. 1993. Cadmium uptake in gills of the mollusk Crassostrea virginica and inhibition by calcium channels blockers. Aquatic Toxicology 24: 195-206. [ Links ]
Rubin, E. & J. L. Farber. 1990. Patología. Editorial Médica Panamericana, S. A de C. V. México, D. F. 2-58. [ Links ]
VILLANUEVA, F. & A. V. BOTELLO. 1998. Metal pollution in coastal areas of Mexico. Environmental Contamination and Toxicology 157: 53-94. [ Links ]
ZAROOGIAN, G. E. & P. YEVICH. 1994. The nature and function of the brown cell in Crassostrea virginica. Marine Environmental Research 37: 355-373. [ Links ]
ZAROOGIAN, G. E. & G. MORRIS. 1981. Effect of cadmium body burdens in adult Crassostrea virginica on fecundity and viability of larvae. Environmental Contamination and Toxicology 27: 344-348 [ Links ]

