











 text new page (beta)
text new page (beta) English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO 
 Permalink
PermalinkINTRODUCTION
Plants are susceptible to a wide diversity of pathogens that can affect their physiology, growth and cause death (Creissen et al., 2016). Synthetic fungicides are widely used to control phytopathogenic fungi; however, these chemicals can exert a negative impact on the environment, favoring the development of pathogen resistance and reducing beneficial native microorganisms (Law et al., 2017). As a viable alternative, antagonistic fungi, bacteria and actinomycetes can be used in agriculture as biocontrol agents, since they inhibit the development of phytopathogens either by competition for space, nutrients, oxygen or by the production of antimicrobial substances (Narayanasamy, 2013). Additionally, some of these anti-pathogenic microorganisms produce plant growth promoting substances, mainly indole acetic acid and siderophores, increasing their importance in agriculture (Gopalakrishnan et al., 2011; Khamna et al., 2009).
The activity of actinomycetes has been mainly related to their ability to produce various secondary metabolites during the sporulation, which is highly vulnerable to the nutrient source (Hwang et al., 2014). They can exist as filamentous or sporogenous forms, which are differentially induced by environmental conditions. The Actinomycetales order comprises a wide diversity of genera, with the Streptomyces genus being one of the most recognized worldwide due to its distribution and ability to produce many of the known antibiotics (Berdy, 2005). Actinomycetes are morphologically diverse in color and mycelial structure, and may or may not produce aerial mycelium or substrate mycelium and diffusible pigments (Hazarika and Thakur, 2020). Streptomyces strains with different morphologies have also been evidenced, despite being phylogenetically close (Sengupta et al., 2015). Each genus and species of actinomycetes has particular requirements for growth, as well as a distinctive production of metabolites and, consequently, a different antagonistic activity. The diversity and activity of actinomycetes depend on their geographical origin due to the influence of environmental conditions (Law et al., 2017). Hence, biocontrol agents, including actinomycetes, usually show their maximum efficacy when applied in regions with environmental conditions similar to those where they were isolated (Suprapta, 2012); however, before being proposed as potential biocontrol agents in crop protection, they must be fully characterized biologically, biochemically, morphologically and molecularly (Jeffrey, 2008). Thus, the objective of this study was to isolate and characterize native actinomycetes from different agroecosystems of Chihuahua, Mexico with putative antagonistic activity against phytopathogenic fungi.
MATERIALS AND METHODS
Reagents
All reagents used were analytical grade (Sigma Aldrich, St. Louis, Missouri, USA) and a DNA Clean & Concentrator kit from Zymo Research (Irvine, California, USA).
Actinomycete isolation and screening for antifungal activity
A total of forty composite soil samples (250 g) were collected from the rhizosphere (10-15 cm depth) of oak forests, grasslands and agroecosystems (bean, maize and apple crops) from nine regions of Chihuahua, Mexico. For actinomycete isolation, serial dilutions (1:10) were performed in test tubes containing 9 mL of sterile peptone water by adding 1 g of sieved soil, to generate 105 to 107 dilutions. Aliquots (50 µL) of each suspension were spread in duplicate by diffusion technique on 90-mm Petri dishes containing one of the following 12 semi-selective media: Bennett’s Medium (AB), Glycerol Nutrient Agar (ANG), Ashby’s Glucose Agar (ASH), Czapek Peptone Agar (CPA), Czapek Dox Agar with 3% glucose (CZAS), Gauze’s Medium No. 1 (GAU), Glycerol Glycine Agar (GGA), GYM Streptomyces Agar (GYM), LB Medium Lenox with agar (LA), Miller with agar (LB), Oatmeal Agar [ISP Medium 3 (OAT)] and V8 Juice Agar (V8) (Atlas, 2010). The isolates were confronted in vitro with Fusarium equiseti and Sclerotium rolfsii to preliminarily screen them according to their antagonistic activity using the arbitrary scale of Pérez-Corral et al. (2015). Only actinomycete isolates with an inhibition of radial growth (IRG) > 50 % were evaluated for morphology, molecular characterization, hydrolyzation of various substrates, use of different energy sources and antagonistic activity against F. equiseti, F. oxysporum, Alternaria alternata and S. rolfsii.
Morphological characterization
Actinomycetes were cultured in four media (PDA, ISP Medium 2, ISP Medium 3 and ISP Medium 4) at 28 ± 1 °C in darkness. The macroscopic characteristics (pigment production and color of the aerial and substrate mycelium) and microscopic characteristics (arrangement and shape of conidia) were evaluated after 7, 14 and 21 d and were used to group the isolates. Microscopic characters were observed using an optical microscope (Carl Zeiss, Jena, Germany) at 1000x magnification, according to Shirling and Gottlieb (1966).
Molecular characterization
The extraction of genomic DNA (gDNA) was performed according to Ruiz-Cisneros et al. (2017). The gDNA was used to amplify the 16S rRNA gene region by PCR using the following pair of primers: F243 (5’-GGATGAGCCCGCGGCCTA-3’) and R1378 (5’-CGGTGTGTACAAGGCCCGGGAACG-3’) (Heuer et al., 1997). The PCR products were separated by electrophoresis on a 1 % agarose gel and subsequently were sequenced at Macrogen Company (Rockville, Maryland, USA). The obtained sequences were assembled and deposited to NCBI database (GenBank).
A phylogenetic tree was constructed by maximum likelihood algorithm and Tamura 3-parameter model to observe the grouping of actinomycetes, using Mega X software (Kumar et al., 2018), with values based on 1000 bootstrapping replicates, after multiple alignment of data by CLUSTAL_X software. An all-versus-all pairwise nucleotide sequence similarity alignment matrix was constructed with EzBioCloud (https://www.ezbiocloud.net/).
Biochemical characterization
The ability of actinomycete isolates to hydrolyze starch, gelatin, casein or urea was determined according to Atlas (2010). Media used were starch agar, gelatin agar, skim milk agar and Christensen base agar. In the first three media, clear zones around the actinomycete colonies were observed for the positive isolates, while the hydrolysis of urea was evidenced by a color change from yellow to pink. The production of hydrogen sulfide (H2S), siderophores and indole acetic acid (IAA) were determined according to Gopalakrishnan et al. (2011) and Louden et al. (2011). For the production of hydrogen sulfide and siderophores, actinomycete isolates were cultured on iron-triple sugar agar (TSI test) and chrome azurol S (CAS), respectively, after incubation at 28 ± 1 °C for 4 and 7 d, respectively, a dark color (black/purple) and orange halos were observed in the medium for the positive isolates, respectively. Production of IAA was determined in test tubes containing ISP medium 2 supplemented with L-tryptophan, positive isolates showed a pink color in the broth medium (Gopalakrishnan et al., 2011; Khamna et al., 2009). Various compounds [citrate, chicory inulin, cellulose, cellobiose, D-sorbitol, D (+)-xylose, mannitol, dulcitol, L (+)-arabinose, adonitol, maltose, myoinositol, lactose, D(-)- fructose, L (-)-sorbose, sucrose, D (+)-galactose, D (+)-trehalose, crab shell chitin and soluble starch] were evaluated as energy sources for actinomycetes; all of them were sterilized using 50 mL of ethyl ether for every 10 g of medium and then, the solvent was evaporated to dryness. One hundred mL of sterile distilled water were added to the medium to obtain a final concentration of 1 %. For the evaluation, the keys proposed by Shirling and Gottlieb (1966) were used.
In vitro antagonistic capacity
Fusarium equiseti, F. oxysporum, A. alternata and S. rolfsii were used to evaluate the in vitro antagonistic effect of selected actinomycete isolates. Explants (7 mm diameter) of each putative antagonistic actinomycete isolate were inoculated at four equidistant spots in a Petri dish (90 mm diameter containing Czapek Dox Agar medium) and incubated at 28 ± 1 °C for 10 d. Subsequently, an explant (7 mm diameter) of the phytopathogen (unquantified mycelium and conidia) was placed at the center of the Petri dish and incubated as described above. Radial growth of the fungal colonies alone (control) and in confrontation with antagonistic actinomycetes were measured every 24 h, until fungi filled the control Petri dish. The IRG was evaluated according to Evangelista-Martínez (2014).
Statistical analysis
The study was carried out using a completely randomized design. Each fungal colony was measured in four orientations with ten experimental units each. The data were analyzed using the Statistical Analysis System software version 9.1 (SAS Institute, 2004) for a balanced analysis of variance, and means were separated by the Tukey test (P ≤ 0.05).
RESULTS AND DISCUSSION
Actinomycete isolates
A total of 1763 actinomycete isolates were obtained from the soil samples. Soil has been shown to be exceptionally rich in Streptomyces, with many species having antifungal activity (Saravana Kumar et al., 2014). The V8, GAU and GYM media were the most efficient, with which 16.7, 14.1 and 12.8 % of the total isolates were obtained, respectively (Table S1). The content of calcium carbonate, starch and yeast extract in these media probably favored the actinomycete isolation because calcium carbonate inhibits Gram-negative bacteria and fungi, while starch and yeast extract are sources of readily available nutrients for actinomycetes (Rodríguez et al., 2018).
Table S1 Substrate hydrolysis and production of plant growth promoters by selected actinomycete isolates.
| Isolate code | Antagonistic strain | Substrate† | Production of plant growth promoters | |||||||
| Hydrogen sulfide | Citrate | Gelatin | Starch | Urea | Casein | Indole acetic acid†† | Siderophores¶ | |||
| A01 | Streptomyces fildesensis CIAD-CA01 | - | - | + | + | - | + | 2 | 2 | |
| A02 | S. paradoxus CIAD-CA02 | + | + | + | + | + | + | 2 | 2 | |
| A03 | S. longisporoflavus CIAD-CA03 | - | + | + | + | + | + | 2 | 2 | |
| A05 | S. caeruleatus CIAD-CA05 | + | + | + | + | + | + | 2 | 2 | |
| A06 | S. turgidiscabies CIAD-CA06 | - | + | + | + | + | + | 1 | 2 | |
| A07 | S. cangkringensis CIAD-CA07 | - | + | + | + | + | + | 1 | 3 | |
| A08 | S. turgidiscabies CIAD-CA08 | - | - | + | + | + | + | 2 | 1 | |
| A10 | S. diastatochromogenes CIAD-CA10 | + | - | + | + | + | + | 1 | 3 | |
| A11 | Streptomyces sp. CIAD-CA11 | - | + | + | + | - | + | 1 | 3 | |
| A13 | Amycolatopsis lexingtonensis CIAD-CA13 | - | - | + | - | + | + | 0 | 0 | |
| A14 | S. ossamyceticus CIAD-CA14 | - | - | + | + | + | + | 2 | 2 | |
| A15 | Saccharopolyspora shandongensis CIAD-CA15 | - | + | + | + | + | + | 4 | 2 | |
| A18 | S. glomeratus CIAD-CA18 | - | - | - | + | - | - | 2 | 2 | |
| A19 | Streptomyces sp. CIAD-CA19 | - | - | - | + | + | - | 0 | 3 | |
| A20 | Nocardioides albertanoniae CIAD-CA20 | - | + | + | + | - | + | 3 | 2 | |
| A23 | S. cellostaticus CIAD-CA23 | - | - | + | + | + | + | 1 | 2 | |
| A24 | S. pseudovenezuelae CIAD-CA24 | + | + | + | + | - | + | 0 | 2 | |
| A25 | S. flavidovirens CIAD-CA25 | + | + | + | + | + | + | 1 | 2 | |
| A27 | S. misionensis CIAD-CA27 | + | - | + | + | - | - | 2 | 2 | |
| A29 | S. nogalater CIAD-CA29 | - | + | + | + | - | + | 3 | 2 | |
| A30 | A. lurida CIAD-CA30 | - | - | + | - | + | + | 0 | 4 | |
| A32 | S. canus CIAD-CA32 | + | + | + | + | + | + | 2 | 2 | |
| A33 | S. fimbriatus CIAD-CA33 | - | + | + | + | + | + | 0 | 1 | |
| A36 | S. canus CIAD-CA36 | + | - | + | + | + | + | 1 | 2 | |
| A37 | Streptomyces graminilatus CIAD-CA37 | - | - | + | + | + | + | 0 | 1 | |
| A40 | S. bungoensis CIAD-CA40 | + | - | + | + | + | + | 0 | 1 | |
| A41 | S. heliomycini CIAD-CA41 | - | + | + | + | + | + | 0 | 2 | |
| A42 | S. graminilatus CIAD-CA42 | - | + | + | + | + | + | 2 | 0 | |
| A43 | S. tricolor CIAD-CA43 | - | + | + | + | + | + | 1 | 2 | |
| A44 | S. heliomycini CIAD-CA44 | - | - | + | + | + | + | 2 | 1 | |
| A45 | S. kanamyceticus CIAD-CA45 | - | + | + | + | - | + | 0 | 2 | |
| A47 | Streptomyces sp. CIAD-CA47 | - | - | + | + | - | + | 0 | 2 | |
| A48 | S. kanamyceticus CIAD-CA48 | - | + | + | + | - | + | 0 | 2 | |
| A67 | S. lydicus CIAD-CA67 | - | - | - | + | - | - | 0 | 3 | |
| A68 | S. glebosus CIAD-CA68 | - | - | - | + | + | + | 1 | 2 | |
| A71 | S. scabrisporus CIAD-CA71 | - | - | + | - | - | + | 0 | 0 | |
| A73 | S. neopeptinius CIAD-CA73 | + | - | + | + | - | + | 2 | 3 | |
| A77 | Streptomyces sp. CIAD-CA77 | - | + | + | + | - | + | 1 | 2 | |
| A79 | S. bottropensis CIAD-CA79 | - | - | + | + | + | + | 1 | 2 | |
| A81 | Streptomyces sp. CIAD-CA81 | - | - | + | + | + | + | 0 | 2 | |
| A82 | Streptomyces sp. CIAD-CA82 | - | - | + | + | + | + | 2 | 3 | |
| A86 | S. libani CIAD-CA86 | - | - | + | + | - | - | 0 | 2 | |
| A91 | S. bobili CIAD-CA91 | + | + | + | + | + | + | 1 | 1 | |
| A95 | S. tubercidicus CIAD-CA95 | - | - | + | + | - | + | 2 | 2 | |
† Substrate hydrolysis: positive (+), negative (-). †† Production of indole acetic acid: negative (0), positive (1), low production (2), average production (3), high production (4). ¶ Production of siderophores: negative (0), positive (1), halo of 1-3 mm (2), halo of 4-6 mm (3), halo > 7 mm (4) according to Gopalakrishnan et al. (2011).
The efficiency of the medium to isolate microorganisms is highly dependent on their particular nutrimental requirements; thus, a formulation can favor a group of microorganisms but not others (Tanaka et al., 2014). The nutritional content of the medium influence the production of antimicrobial compounds (Abdelmohsen et al., 2015). Bredholdt et al. (2007) obtained 2680 actinomycete isolates from Trondheim fjord (Norway) sediments and highlighted the importance of using selective methods and media for the isolation of a wide variety of microorganisms; moreover, colonies of a microorganism can show different macroscopic characteristics in different media, such as the color of mycelium and diffusible pigments (Barakate et al., 2002).
Pigment production is one of the distinctive characteristics of actinomycetes, which depends on the medium composition, growing conditions and colony age (Attimarad et al., 2012). Several secondary metabolites are produced by actinomycetes during the mycelial shift from substrate to sporulation phases, a process accompanied by the formation of aerial multinucleated mycelium (Flärdh and Buttner, 2009). Of the 1763 actinomycete isolates, 498 (28 %) and 557 (31.6 %) showed antagonistic activity against F. equiseti and S. rolfsii, respectively, while 44 (2.5 %) generated IRG values higher than 50 %, results that are similar to those of Attimarad et al. (2012), who found significant antibacterial activity in 35 % of their isolated actinomycetes. Ganesan et al. (2017) observed that 41.5 % of tested actinomycete isolates showed significant (4.7-25.4 %) antimicrobial activity. In this study, 23 of the 44 isolates with significant anti-pathogenic activity were isolated from apple orchard soil, eight from maize plots, seven from bean plots, and six from grassland soil (Table S1). Antagonistic microorganisms were abundant in the apple orchard soil, probably due to frequent addition of organic matter. The abundance and diversity of actinomycetes in soil is highly influenced by soil type, geographic origin and organic matter content (Arifuzzaman et al., 2010).
Morphology
The isolated actinomycetes grew well in all semi-selective media, showing hard colonies at the 7th day and a dusty appearance after 14 days due to fragmentation of the mycelium. The mycelium developed a gray, yellow, white or red pigmentation in 44.9 %, 26.1 %, 17.1 % and 11.9 % of the cultures, respectively, at the 21st day. In other experiments, proportions of 40 % of isolated actinomycetes have been found with gray mycelium, while only 2 % of them developed a red mycelium (Barakate et al., 2002). In this study, both substrate and aerial mycelium developed the same colors, except white. Sadeghi et al. (2014) observed that the color of the aerial mycelium of an actinomycete depended on the type of medium, showing a white color in ISP 3, 4 and 6, Czapek’s agar and nutritive agar. Diffusible pigments (yellow, red or gray) were observed in only four (9.1 %) isolates (Figure S1). Color plays an important role in the classification and identification of actinomycetes, especially for the Streptomyces genus due to its high morphological variability, with each isolate showing a specific color depending on the age and type of medium (Barakate et al., 2002). In this study, the isolates mainly showed straight, flexible, open spiral and spiral arrangements (Figure 1), as previously reported for these microorganisms (Shirling and Gottlieb, 1966) with a great morphological variability, especially for the Streptomyces species.

Molecular characterization
PCR amplicons showed a molecular size of ≈ 1000-1100 bp. The 16S rRNA gene was analyzed and compared with the NCBI/BLAST database, showing high similarity with different genera and species of actinomycetes, with an identity of 99 to 100 %. The high content of G + C (55-75 %) in actinomycete DNA is well known (Ganesan et al., 2017; Hwang et al., 2014). The main genera found in this study were Streptomyces (90.9 %), Amycolatopsis (4.6 %), Saccharopolyspora (2.3 %) and Nocardioides (2.3 %). DNA sequences were deposited in the GenBank under accession numbers MK968571 to MK968614. Similarly, Cheeptham et al. (2013) observed that 79.3 % of isolates from volcanic caves belonged to the Streptomyces genus; thus, the Streptomyces genus is highly predominant in soil and plant tissues.
Phylogenetic analysis of the actinomycete isolates showed separations within genera and species. Strains belonging to the genera Amycolatopsis and Saccharopolyspora were found within the same clade, while those of the Streptomyces genus were located in several subclades (Figure 2), coincidentally the latter were the most abundant in the agricultural soils of Chihuahua, Mexico and have the highest antifungal potential. These variations might be related to the genetic variability, as a result of their evolution and adaptation in a great range of environmental niches (agricultural soils) from where they were isolated (Lewis et al., 2010), which cause differences in the morphology, metabolism and antimicrobial activity (Quecine et al., 2008). Although various Streptomyces species are closely related, there are strains among the same species capable of producing different antifungal compounds (Al_husnan and Alkahtani, 2016), possibly giving them a competitive and evolutionary advantage in the symbiotic associations in the soil (Kinkel et al., 2012).
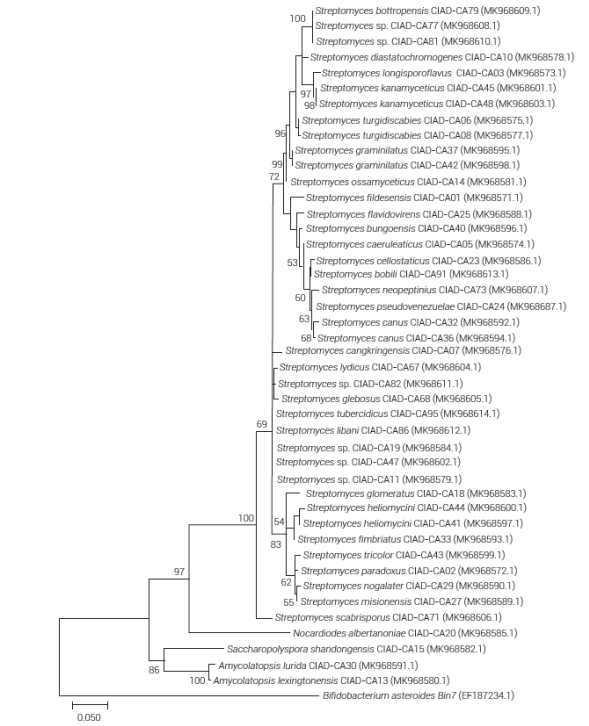
Figure 2 Phylogenetic tree constructed with the maximum likelihood algorithm and the Tamura 3-parameter model based on 16S rRNA gene region sequences of actinomycetes. Positions with less than 95 % site coverage were eliminated. The tree is drawn to scale, with branch lengths measured in number of substitutions per site. Scale bar represents five nucleotide substitutions per 100 nucleotides. Numbers at branch points indicate values as percentage based on 1000 bootstrap replicates. Support values exceeding 50 % are shown.
The use of energy sources was highly dependent on the type of isolate (Table 2). Sucrose, maltose, lactose and D (+)-trehalose were readily utilized by more than 82 % of the isolates. Soluble starch was the most utilized by the tested actinomycetes. L (-)-sorbose, chicory inulin and D-sorbitol were the least used. Arasu et al. (2008) found that similar energy sources as maltose, galactose, mannitol, glycerol, arabinose, xylose, starch and glucose are highly used by Streptomyces, while lactose was the least used. Findings of this study demonstrated that actinomycetes can use a wide diversity of carbon sources, varying among strains of the same species. Gil et al. (2009) demonstrated that actinomycetes have the ability to use a wide variety of compounds as energy source, mainly glucose, starch, amino acids and proteins.
Table 2 Use of energy sources by selected actinomycete isolates.
| Isolate code | Antagonistic strain | Chicory inulin | Celulose | Cellobiose | D-Sorbitol | Dulcitol | Sucrose | L(-)-Sorbose | D(+)-Xylose | D(-)-Fructose | Mannitol | Myo-Inositol | L(+)-Arabinose | Maltose | Lactose | D(+)-Galactose | Adonitol | D(+)-Trehalose | Soluble starch | Crab-shell chitin |
| A01 | Streptomyces fildesensis CIAD-CA01 | ± | + | + | + | + | ± | - | + | + | ± | ++ | ± | + | ± | ± | - | + | ++ | + |
| A02 | S. paradoxus CIAD-CA02 | ++ | - | - | - | - | + | - | ++ | - | - | - | - | - | - | ++ | ++ | ++ | ++ | ++ |
| A03 | S. longisporoflavus CIAD-CA03 | + | ± | ± | - | - | ++ | - | + | + | + | + | + | + | + | ± | - | + | + | + |
| A05 | S. caeruleatus CIAD-CA05 | - | - | ± | - | - | + | - | ± | ++ | + | - | + | + | + | ± | - | ± | ++ | + |
| A06 | S. turgidiscabies CIAD-CA06 | ± | ± | + | - | - | + | - | ± | - | ± | ± | ± | - | - | ± | ± | + | + | ± |
| A07 | S. cangkringensis CIAD-CA07 | - | ++ | - | - | ++ | - | ++ | ++ | - | - | - | - | - | ++ | ++ | - | - | ++ | ++ |
| A08 | S. turgidiscabies CIAD-CA08 | ± | ± | ± | - | ± | ± | - | - | - | - | - | + | - | ± | + | ± | + | ++ | ± |
| A10 | S. diastatochromogenes CIAD-CA10 | ± | + | + | ++ | ± | + | - | + | - | ± | - | + | ++ | ± | + | ± | ++ | ++ | - |
| A11 | Streptomyces sp. CIAD-CA11 | - | ± | ± | ++ | ± | ± | - | - | ± | - | - | - | ++ | ± | + | ± | ++ | ++ | ± |
| A13 | Amycolatopsis lexingtonensis CIAD-CA13 | - | + | ± | ± | ± | + | - | + | + | + | + | + | + | + | + | ± | + | + | - |
| A14 | S. ossamyceticus CIAD-CA14 | ± | ± | - | - | - | ± | - | ± | ± | ± | - | + | + | + | + | ± | + | + | ± |
| A15 | Saccharopolyspora shandongensis CIAD-CA15 | - | ++ | + | - | ± | ++ | - | - | - | - | - | - | + | - | - | + | ++ | ++ | - |
| A18 | S. glomeratus CIAD-CA18 | ± | ± | ± | - | ± | ± | ± | ± | + | ± | ± | - | + | ± | ± | - | + | + | ± |
| A19 | Streptomyces sp. CIAD-CA19 | - | ± | ± | + | ± | + | - | - | ± | ± | + | - | + | ± | ± | ± | + | + | + |
| A20 | Nocardioides albertanoniae CIAD-CA20 | - | + | ± | ++ | - | + | - | ± | - | + | - | ± | - | ++ | ++ | ++ | ++ | ++ | ± |
| A23 | S. cellostaticus CIAD-CA23 | - | - | + | - | ± | + | - | ± | ± | ± | ± | ± | ± | ± | ± | ± | + | + | ± |
| A24 | S. pseudovenezuelae CIAD-CA24 | ± | ± | ± | + | ± | + | - | ± | + | + | ± | + | + | ± | + | ± | ± | + | - |
| A25 | S. flavidovirens CIAD-CA25 | - | ± | + | - | ± | + | - | ± | + | ± | ± | + | + | + | + | - | + | + | - |
| A27 | S. misionensis CIAD-CA27 | - | + | ± | - | ± | - | - | + | + | + | ± | + | + | - | - | ± | ++ | ++ | ++ |
| A29 | S. nogalater CIAD-CA29 | - | - | ± | - | - | - | - | ++ | - | - | - | - | - | ± | ++ | - | ± | ++ | - |
| A30 | A. lurida CIAD-CA30 | - | + | + | - | ± | ± | - | ± | ± | ± | ± | ± | + | ± | + | + | ± | + | ++ |
| A32 | S. canus CIAD-CA32 | ++ | ++ | ++ | ± | ++ | ++ | - | - | ± | + | - | ± | ++ | + | + | ++ | ++ | ++ | ++ |
| A33 | S. fimbriatus CIAD-CA33 | ± | + | ± | ± | ± | ± | - | + | ± | ± | ± | ± | ++ | ++ | ++ | + | + | ++ | ++ |
| A36 | Streptomyces canus CIAD-CA36 | - | - | - | - | - | ± | - | ± | - | - | - | ± | ++ | + | ± | + | - | + | - |
| A37 | S. graminilatus CIAD-CA37 | ± | ± | ± | ± | ± | ++ | - | ± | ++ | ± | + | + | ++ | + | ± | ± | ± | ++ | + |
| A40 | S. bungoensis CIAD-CA40 | - | - | - | - | ++ | ++ | - | ++ | - | - | - | ++ | ++ | ++ | - | ++ | - | + | - |
| A41 | S. heliomycini CIAD-CA41 | - | ± | ± | + | ± | + | - | ± | + | - | ± | ± | + | + | + | + | ± | ++ | - |
| A42 | S. graminilatus CIAD-CA42 | ± | - | - | - | - | + | - | + | + | - | + | + | ++ | + | ± | - | ± | + | ± |
| A43 | S. tricolor CIAD-CA43 | - | ± | - | - | - | ± | - | - | - | - | - | - | - | ± | ± | - | ± | + | ± |
| A44 | S. heliomycini CIAD-CA44 | - | ± | ± | - | ± | + | - | ++ | + | + | + | ++ | ++ | + | ++ | ++ | ++ | ++ | ++ |
| A45 | S. kanamyceticus CIAD-CA45 | - | - | - | - | ± | - | - | - | - | ± | - | + | + | + | ± | ± | ++ | ++ | - |
| A47 | Streptomyces sp. CIAD-CA47 | - | - | - | ± | - | ± | - | - | ± | - | - | - | + | - | - | ± | ± | ++ | - |
| A48 | S. kanamyceticus CIAD-CA48 | ± | ± | ± | + | ± | ± | ± | ± | + | + | ± | + | + | + | ± | ± | + | + | ± |
| A67 | S. lydicus CIAD-CA67 | - | - | - | - | - | - | - | - | ± | - | - | - | + | + | - | - | - | ++ | - |
| A68 | S. glebosus CIAD-CA68 | ± | ± | ± | ± | ± | ± | - | ± | + | ± | ± | - | + | ++ | ± | ± | - | ++ | ± |
| A71 | S. scabrisporus CIAD-CA71 | - | ± | ± | ± | - | ± | - | ± | ± | - | ± | ± | + | - | ± | ± | ± | ++ | ++ |
| A73 | S. neopeptinius CIAD-CA73 | + | ± | + | + | + | + | - | ++ | ± | - | ± | - | ± | ++ | + | + | + | ++ | + |
| A77 | Streptomyces sp. CIAD-CA77 | + | ± | ± | ± | ± | ± | - | + | + | ++ | + | + | + | + | ++ | ± | + | ++ | + |
| A79 | S. bottropensis CIAD-CA79 | ± | - | - | ± | ± | + | ± | ± | + | - | ± | + | ++ | ± | ± | ± | ± | + | ± |
| A81 | Streptomyces sp. CIAD-CA81 | + | - | - | + | - | - | - | ± | - | ++ | + | + | ++ | + | ++ | ± | + | ++ | ± |
| A82 | Streptomyces sp. CIAD-CA82 | ± | ± | - | ± | ± | - | - | - | - | - | - | - | - | ++ | - | + | + | ++ | - |
| A86 | S. libani CIAD-CA86 | + | - | + | ± | + | + | - | - | ++ | - | - | - | ++ | ++ | ± | ± | - | ++ | ± |
| A91 | S. bobili CIAD-CA91 | - | ± | + | ± | - | - | - | - | - | ± | ± | ± | + | ± | ± | ± | - | + | - |
| A95 | S. tubercidicus CIAD-CA95 | - | - | - | - | - | ± | - | - | ++ | - | - | - | + | - | - | - | - | ++ | + |
Strong positive utilization (++), positive utilization (+), ambiguous utilization (±), negative utilization (-), according to Shirling and Gottlieb (1966).
Ninety three percent of the actinomycetes tested produced siderophores. Isolate A30 (A. lurida CIAD-CA30) showed the highest siderophore production (halo diameter > 7 mm) on CAS agar (Table 1); however, this isolate did not produce IAA and only showed antifungal activity against S. rolfsii (IRG > 86 %) (Figure 3). Twenty-seven isolates (61.36 %) produced siderophores, with halos of 4-6 mm. Three isolates (6.82 %) did not produce siderophores while the rest of isolates (29.55 %) showed halos of less than 4 mm (Table 1). Khamna et al. (2009) demonstrated that siderophores fix Fe3+ in the rhizosphere, making it inaccessible for phytopathogens, although these forms of Fe3+ can be used by some microorganisms and plants; thus, siderophores from actinomycetes negatively affect various fungal pathogens, protecting crops (Ahmed and Holmström, 2014). Siderophores can promote the solubility of mineral nutriments in the soil, making them available for microorganisms and plants (Singh et al., 2017). Many isolates (66 %) produced IAA (Table 1). Isolate A15 (Saccharopolyspora shandongensis CIAD-CA15) showed the highest IAA production, and like A. lurida CIAD-CA30, only showed antifungal activity against S. rolfsii (IRG > 100 %). Isolates A20 (Nocardioides albertanoniae CIAD-CA20) and A29 (S. nogalater CIAD-CA29) also produced IAA although to a lesser extent. In the rhizosphere, the plant exudates are the main sources of tryptophan, a precursor in auxin (IAA) biosynthesis, promoting root elongation (Khamna et al., 2009). The similarity in the Jaccard matrix with data of the biochemical tests (Table S2) allowed determining that the 44 tested isolates were different, in contrast to the similarity values of the 16S rRNA shown by some isolates (Table S3).
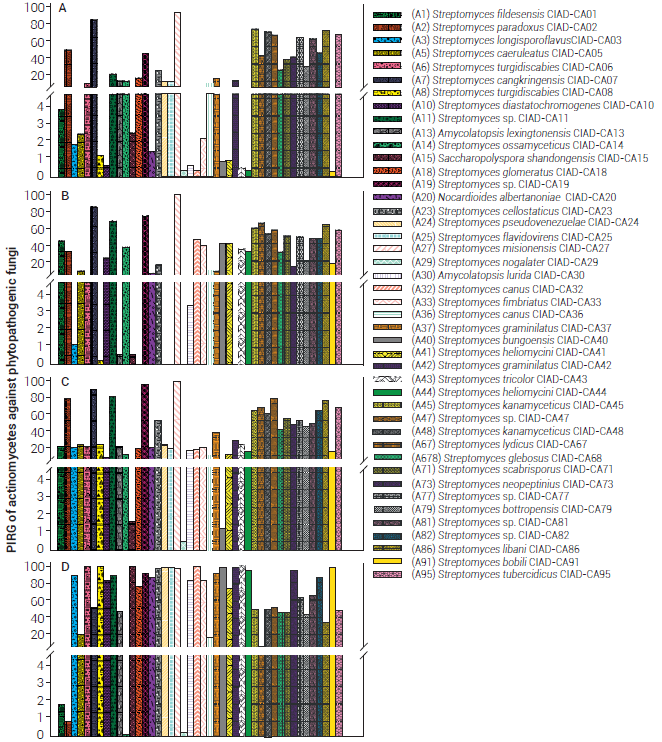
Figure 3 Inhibitions of radial growth (IRG) of actinomycetes against phytopathogenic fungi: A) Fusarium equiseti, B) Fusarium oxysporum, C) Alternaria alternata and D) Sclerotium rolfsii .
Isolates A11 (Streptomyces sp. CIAD-CA11), A19 (Streptomyces sp. CIAD-CA19), A47 (Streptomyces sp. CIAD-CA47), A86 (S. libani CIAD-CA86) and A95 (S. tubercidicus CIAD-CA95) showed a similarity of 100; however, values lower than 0.5 were found with the Jaccard analysis. Discordant results were found for A45 (S. kanamyceticus CIAD-CA45) and A48 (S. kanamyceticus CIAD-CA48), with a similarity in 16S rRNA of 99.8 and 0.8 in the Jaccard matrix, although belong to the same species they show different behavior; alike, A32 (S. canus CIAD-CA32) and A36 (S. canus CIAD-CA36) showed values of 99.3 and 0.5 respectively. This behavior was confirmed with the IRG values for each isolate confronted with phytopathogenic fungi (Figure 3), with IRGs of 100 and 16% against S. rolfsii, respectively, both isolated from soil of apple trees from different regions of Chihuahua, Mexico (Table S1).
Antagonistic capacity
The IRGs of actinomycetes ranged from 0.3 to 92.1 % for F. equiseti, from 0 to 98.4 % for F. oxysporum, from 0.4 to 97 % for A. alternata and from 0.2 to 100 % for S. rolfsii (Figure 3). Seven isolates completely inhibited (IRG = 100 %) the growth of S. rolfsii, probably due to their capacity to produce chitinases and glucanases, which degrade fungal cell walls (El-Tarabily et al., 2000). Isolates that caused IRG values > 50 % against F. equiseti, F. oxysporum, A. alternata and S. rolfsii were 9, 11, 14, and 29, respectively (Figure 3). Fifteen isolates inhibited the growth of more than one phytopathogen with IRG values > 50 %. Isolates A07 (Streptomyces cangkringensis CIAD-CA07), A27 (S. misionensis CIAD-CA27) and A45 (S. kanamyceticus CIAD-CA45) showed IRG values > 50 % for all phytopathogenic fungi; isolate A27 (S. misionensis CIAD-CA27) showed the highest IRGs (92.1-99.1 %). Sclerotium rolfsii was the most susceptible phytopathogen to most actinomycetes, while F. equiseti was the most resistant one (Figure 3). Isolate A27 (S. misionensis CIAD-CA27), obtained from a bean farmland, showed the highest antagonistic capacity, probably due to its ability to produce both volatile and non-volatile antifungal compounds, as has been observed for other actinomycetes with high antifungal activity (Boukaew et al., 2013). Boukaew and Prasertsan (2014) inferred that the activity of actinomycetes is due to the biosynthesis of compounds that induce alterations in the cell-wall structure of pathogens, leading to leakage of cytoplasmic material.
Several actinomycetes with the highest IRGs belong to the Streptomyces genus, although the non-streptomyces isolates A15 (S. shandongensis CIAD-CA15), A20 (N. albertanoniae CIAD-CA20) and A30 (A. lurida CIAD-CA30) also showed high IRG values (100, 87.1 and 86.2 %, respectively), at least against S. rolfsii.
Evangelista-Martínez (2014) observed high variability of IRGs with the confrontation of Streptomyces species with Curvularia sp. (53-73 %), Helminthosporium sp. (44-71 %), Aspergillus niger (81-96 %) and Fusarium sp. (42-57 %). Boukaew and Prasertsan (2014) found IRGs from 82 to 89 % between Streptomyces philanthi RM-1-138 and several phytopathogenic fungi such as Rhizoctonia solani, Pyricularia grisea, Bipolaris oryzae, F. fujikuroi, Ganoderma boninense, Colletotrichum gloesporioides and C. capsici. Inhibition of phytopathogens by actinomycetes can be due to both volatile and non-volatile inhibitory compounds (Boukaew et al., 2013). Cao et al. (2005) demonstrated that plant-growth promoting substances, like siderophores, are involved in the antagonistic activity of actinomycetes. Results of this study demonstrated the in vitro antagonistic capacity of some actinomycete isolates. They might be considered as prospective candidates for use as biocontrol agents against phytopathogenic fungi and also as plant growth promoters.
CONCLUSIONS
The main genera of actinomycetes found were Streptomyces, Amycolatopsis, Saccharopolyspora and Nocardioides. Based on biochemical, morphological and molecular characterization, the Streptomyces genus was the most predominant in agricultural soils of Chihuahua, Mexico. Ninety-three percent of the isolated actinomycetes produced siderophores, while only 66 % produced IAA. The inhibition of radial growth caused by actinomycetes against phytopathogenic fungi was variable, with S. rolfsii being the most susceptible and F. equiseti the most resistant pathogen. The study demonstrated a large amount and diversity of actinomycetes growing in different substrates.

