











 text new page (beta)
text new page (beta) Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO 
 Permalink
PermalinkINTRODUCCIÓN
Los bosques albergan gran diversidad de especies, protegen el suelo, contribuyen a la estabilidad del clima, proveen de bienes y servicios y proporcionan una amplia gama de productos maderables y no maderables (Bergh y Promis, 2011). Sin embargo, el cambio climático podría favorecer el desarrollo de plagas forestales o de especies invasoras en los bosques (Pérez et al., 2007) y en este sentido se ha demostrado que la temperatura es el factor más importante debido a que influye en la supervivencia, mortalidad y en el número de generaciones de insectos, modificando la distribución geográfica de las plagas (Kocmánková et al., 2010). Así mismo, cuando los bosques se ven afectados por factores de estrés, naturales o antropogénicos, se favorece el brote de plagas (Salinas et al., 2010).
En México se cuenta con aproximadamente 42 % de las especies conocidas de Pinus, su distribución altitudinal varía entre 1500 y 4000 msnm (Perry, 1991; Sánchez, 2008). Los insectos descortezadores del género Dendroctonus son huéspedes de varias de estas especies de pinos; por los daños causados se les conoce como las plagas forestales más dañinas de los bosques de pino (Rodríguez et al., 2013). Los descortezadores han causado la muerte de millones de árboles; en Canadá las pérdidas maderables han sido de hasta 2.5 millones de m3 en los últimos 20 años (Miller y Borden, 2010). En México, los daños más notables se observaron recientemente. CONAFOR (2015) registró que entre 2004 y 2014 la superficie afectada fue de 30,365 ha, atribuidas a 12 especies de insectos descortezadores. Sánchez et al. (2003) encontraron en el estado de Coahuila brotes activos de Dendroctonus spp., afectando un volumen total de 36,749 m3 de madera en rollo. Las especies de descortezadores registradas fueron: D. adjunctus Blandford, D. brevicomis LeConte, D. pseudotsugae Hopkins, D. mexicanus Hopkins, D. parallelocollis Chapuis y D. valens LeConte.
Dendroctonus adjunctus es considerada como una plaga primaria y D. brevicomis como secundaria. La primera se distribuye ampliamente en la mayoría de los sistemas montañosos y la segunda presenta distribución restringida (Salinas-Moreno et al., 2010). En México existe poca información sobre la abundancia a diferentes altitudes de estas especies. Algunos estudios relacionados con el tema fueron descritos por Salinas-Moreno et al. (2010), donde mencionan el intervalo de distribución altitudinal de 2500 a 3000 msnm y de 1680 a 3300 msnm, respectivamente. Rodríguez et al. (2010) mencionaron una distribución altitudinal de D. adjunctus de 1600 hasta 3929 msnm, con mayor incidencia de insectos en primavera y otoño. El presente estudio tuvo como objetivo evaluar la fluctuación poblacional, altitudinal y temporal de D. adjunctus y D. brevicomis y su relación con temperatura y variables dasométricas del sitio en el ejido Santa Rita, en Arteaga Coahuila.
MATERIALES Y MÉTODOS
Descripción física y biológica del área de estudio
La localidad Santa Rita, en Arteaga Coahuila, se ubica entre 25º 17’ 36’’ y 25º 16’ 15’’ de latitud norte y entre 100º 30’ 52’’ y 100º 27’ 52’’ de longitud oeste. El clima es templado subhúmedo con lluvias escasas todo el año, variando de 300 a 700 mm anuales; su fórmula climática es BS1K (x’), el intervalo de temperatura es de 8 a 20 ºC, con rango altitudinal del área entre 1300 y 3700 msnm. El tipo de vegetación es bosque de coníferas (INEGI, 2009), cuyas principales especies son: Pinus rudis Endl, P. ayacahuite Ehren, Pseudotsuga menziesii Mirb., Abies vejarii Martínez, Ceanothus coeruleus Lag, C. buxifolius Willd y Garrya ovata Benth (Cano et al., 2007).
Muestreo de insectos descortezadores
En el sitio de estudio se establecieron dos transectos (A y B) los cuales presentaban exposición sur. Cada transecto se conformó por ocho elevaciones, una cada 100 m, iniciando desde 2600 hasta 3300 msnm. A una altura de 1.5 m de un árbol no hospedero de cada elevación se colocó una trampa Lindgren® de ocho embudos con atrayente (frontalina, endo-brevicomina y alfa-pineno) y a 50 m en distancia horizontal se colocó una trampa sin atrayentes. El vaso colector de insectos se colgó sobre la trampa. Para conservar los insectos colectados se utilizó anticongelante marca PRESTONE AF EX al 33 %.
Recolección de insectos, variables climáticas y datos dasométricos del sitio
El periodo de estudio fue de dos años (febrero 2015 a febrero 2017). Las trampas se revisaron cada 15 días, resultando en un total de 52 colectas. Los insectos capturados se llevaron al laboratorio para su identificación con apoyo de un estereoscopio LEICA modelo EZA y claves taxonómicas de Cibrián et al. (1995). En un árbol cercano a la trampa con feromona se instaló un data logger modelo EL-USB-2, a una altura aproximada de 1.5 m, el cual registró temperatura (ºC), humedad relativa (%) y punto de rocío (ºC) cada 30 minutos. La información almacenada se descargó cada seis meses usando el software EasyLogUSB. Para obtener las características del sitio se delimitó una parcela de forma cuadrada (50 x 50 m) tomando como punto central la trampa. De cada árbol se tomó la información: especie, diámetro normal (cm), altura total (m), diámetro de copa (m), densidad de copa (%), estado fitosanitario (sano e infestado), pendiente (%) y exposición (N, S, E y O). El equipo utilizado para medir estas variables fue: distanciómetro Leica DISTO™ D810 touch, densiómetro esférico convexo (DSM43), densiómetro esférico cóncavo (DSM43A), forcípula Haglöf Mantax, cinta diamétrica (283D/10M) y GPS Garmin (GPSMAP 64).
Análisis de datos
Para caracterizar la distribución temporal y altitudinal de cada especie de descortezador, los datos de abundancia se organizaron en los siguientes niveles: por año, estación del año, altitud y transecto. Previo a la comparación y por ser información proveniente de conteos periódicos, los datos se sometieron a una prueba de normalidad de Shapiro-Wilk (α = 0.05); al no cumplir con este supuesto, la comparación de abundancia de insectos en cada nivel se realizó usando la prueba no paramétrica de rangos de Kruskal-Wallis (KW) al 95 % de confiabilidad (Kruskal y Wallis, 1952). Para comparar la abundancia de cada especie de descortezador por año en cada transecto se usó el total de repeticiones sin importar la altitud (n = 208 por especie por año). Entre transectos el análisis se realizó incluyendo el muestreo de ambos años sin importar la altitud (n = 416 repeticiones por especie). En la comparación de abundancia de descortezadores entre estaciones, las repeticiones variaron desde n = 48 hasta 52 (por el número de semanas, pero separadas por año de evaluación y por transecto). La comparación de abundancia entre altitudes incluyó el total de muestras por sitio sin importar el año (n = 52).
Para comprobar si existe relación de la abundancia (promedio de 52 fechas de colecta, ambos años) de D. adjunctus y D. brevicomis en cada sitio de muestreo (altitud) con la temperatura (promedio de todos los registros de temperatura de cada 30 minutos durante dos años) y con las características del sitio (área basal por hectárea, diámetro normal promedio por sitio, altura promedio por sitio y número de árboles por hectárea) se realizó un análisis de correlación de Spearman (n = 8 pares de datos, cada uno derivado del promedio de 52 muestreos). Los análisis estadísticos fueron realizados con el Software InfoStat 2016.
RESULTADOS Y DISCUSIÓN
Fluctuación de D. adjunctus y D. brevicomis entre años y transectos
La abundancia de D. adjunctus y D. brevicomis en el transecto A durante el primer año de evaluación (28/02/15 - 20/02/16) fue más alta (p = 0.0178) que en el segundo año, con medias de 8.36 y 8.50 individuos por trampa para cada especie, mientras que en el segundo año la población fue de aproximadamente 4.5 individuos por trampa para ambas especies. En el transecto B la abundancia en ambas especies fue igual en los dos años (p = 0.9515 y 0.2033) (Cuadro 1). La abundancia de D. adjunctus y D. brevicomis entre transectos fue diferente (p = 0.0001); en el transecto A, con 6.33 y 6.36 individuos en promedio, hubo 2 a 4 veces más individuos que en el transecto B, con 2.63 y 1.64 insectos, respectivamente (Cuadro 1). Esto se debe a que en el transecto A el 33.3 % de las especies arbóreas son hospederas de descortezadores, y en B solo el 20.0 %. Tanto en A como en B, P. rudis es la especie hospedera dominante, incluso el 70 % de los árboles en las altitudes de 3100 a 3200 msnm en A son de P. rudis. La temperatura promedio en los transectos A y B (9.45 y 9.32 ºC, respectivamente) es muy similar, por lo que la presencia de especies hospederas parece ser más importante (Cuadro 2).
Cuadro 1 Prueba de Kruskal-Wallis de la abundancia de Dendroctonus adjunctus y D. brevicomis entre años y transectos altitudinales en Santa Rita, Arteaga Coahuila, México.
| Especie | Transecto | Año | Rango | Medias | Gl | C | H | p |
|---|---|---|---|---|---|---|---|---|
| Comparación entre años (n = 208 por especie) | ||||||||
| D. adjunctus | A | 21/02/16 - 18/02/17 | 196.68 b | 4.31 | 1 | 0.72 | 4.02 | 0.0178 |
| 28/02/15 - 20/02/16 | 220.32 a | 8.36 | ||||||
| B | 28/02/15 - 20/02/16 | 208.79 | 3.31 | 1 | 0.64 | 0.0024 | 0.9515 | |
| 21/02/16 - 18/02/17 | 208.21 | 1.93 | ||||||
| D. brevicomis | A | 21/02/16 - 18/02/17 | 198.43 b | 4.23 | 1 | 0.74 | 2.92 | 0.0477 |
| 28/02/15 - 20/02/16 | 218.57 a | 8.50 | ||||||
| B | 28/02/15 - 20/02/16 | 213.90 | 2.28 | 1 | 0.52 | 0.84 | 0.2033 | |
| 21/02/16 - 18/02/17 | 203.10 | 1.00 | ||||||
| Comparación entre transectos (n = 416 por especie) | ||||||||
| D. adjunctus | B | 399.77 b | 2.62 | 1 | 0.68 | 4.03 | 0.0147 | |
| A | 433.23 a | 6.33 | ||||||
| D. brevicomis | B | 380.79 b | 1.64 | 1 | 0.64 | 18.37 | <0.0001 | |
| A | 452.21 a | 6.36 | ||||||
Medias con letras iguales por columna no son estadísticamente diferentes (Kruskal-Wallis, 0.05). Gl: grados de libertad, C: factor de corrección del estadístico por observaciones empatadas, H: estadístico de la prueba no corregido por empates, p: significancia estadística. Nota: la comparación por año incluyó el muestreo en todas las altitudes en ese año; en transectos se incluyeron ambos años de muestreo en todas las altitudes.
Sánchez et al. (2003) reportaron brotes de D. adjunctus y D. brevicomis en Pseudotsuga menziesii, Abies vejarii, Pinus rudis, P. teocote Schiede ex Schltdl, P. pseudostrobus Lindl y P. arizonica Engelm en la Sierra de Arteaga Coahuila, lo cual concuerda con las especies hospederas de este estudio (P. rudis y Abies vejarii). Torres y Sánchez (2006) mencionaron que bosques densos de Pinus rudis (466 árboles ha-1), son más susceptibles a brotes de D. adjunctus y D. brevicomis debido a la competencia entre árboles y a la susceptibilidad de estrés. Los resultados obtenidos en el presente estudio comprueban que en el transecto A, con 378 árboles ha-1, hubo mayor densidad de arbolado, causando una mayor presencia de descortezadores. En Estados Unidos y Canadá las especies hospederas para D. adjunctus son P. arizonica, Pinus engelmannii Carr., P. flexilis, Pinus leiophylla Schiede ex Schltdl. & Cham., Pinus ponderosa Douglass ex Lawson, Pinus strobiformis Engelm, mientras que D. brevicomis se presenta en Pinus coulteri D. Don y P. ponderosa (Bentz et al., 2010). En Coahuila las especies afectadas son P. rudis, P. flexilis y Abies vejarii; aunado a esto, se ha pronosticado que el aumento de temperatura (de 3 a 5 ºC) para Estados Unidos y Canadá propiciará aumento en las poblaciones de plagas (Dukes et al., 2009).
Fluctuación estacional de D. adjunctus y D. brevicomis
La abundancia de D. adjunctus fue estadísticamente diferente entre estaciones del año (p < 0.0001), siendo mayor en primavera del 2015 en ambos transectos con 19.46 (transecto A) y 8.35 (transecto B) insectos en promedio y casi nula en otoño del 2015 (n = 0.21 en transecto A y n = 0 en B) (Figura 1 A y 1 C). D. brevicomis presentó un patrón similar; las estaciones de mayor vuelo de descortezadores fueron verano del 2015 (transecto A) y primavera del 2015 (transecto B) promediando 15.64 y 6.13 insectos, respectivamente en cada estación. Otoño del 2015 y 2016 fueron las estaciones donde hubo menor presencia de D. brevicomis (n = 0.94 transecto A y n = 0.33 transecto B) (Figura 1 B y 1 D). En el transecto A las temperaturas más altas se registraron en primavera y verano del 2015, siendo de 10 y 13 ºC, respectivamente, ocurriendo aquí la mayor abundancia de insectos. En las estaciones con menor presencia de descortezadores (otoño 2015 y 2016) las temperaturas fueron de 9.10 y 8.83 ºC (Figura 2 A); para el año 2016 se presentó un patrón similar en la abundancia de descortezadores (Figura 1A y 1B).
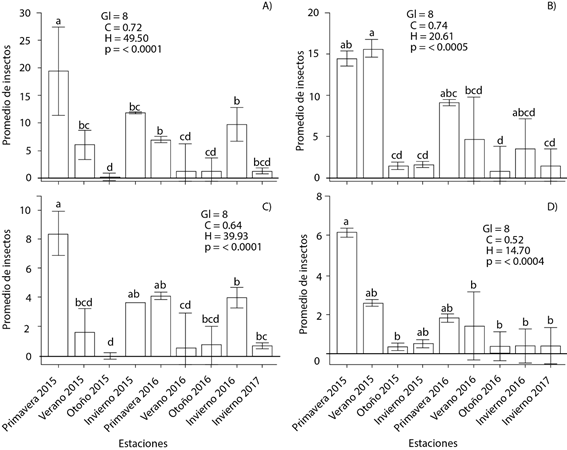
Figura 1 Prueba de rangos de Kruskal-Wallis de la fluctuación temporal en el tamaño de población de Dendroctonus adjuntus (A y C) y D. brevicormis (B y D) en los transectos A (A y B) y B (C y D) en Santa Rita, Arteaga Coahuila, México. Las barras indican los valores medios (± error estándar). Medias con letras iguales (minúsculas) no son estadísticamente diferentes (Kruskal-Wallis, 0.05). Gl: grados de libertad, C: factor de corrección del estadístico por observaciones empatadas, H: estadístico de la prueba no corregida por empates, p: significancia estadística.
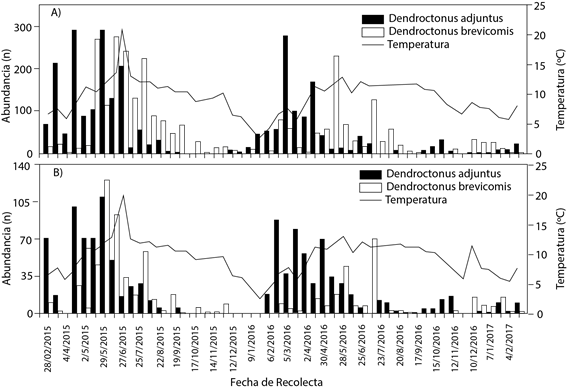
Figura 2 Fluctuación poblacional total de Dendroctonus adjunctus y D. brevicomis por fecha de colecta y su relación con temperatura promedio en los transectos A (A) y B (B) en Santa Rita, Arteaga Coahuila, México. Nota: en el eje “Y” se representa la abundancia total registrada en todas las altitudes en cada fecha de colecta. La temperatura es el promedio aritmético de todos los datos registrados por los sensores en todas las altitudes durante el periodo de una fecha de colecta a la siguiente, en cada transecto.
Primavera del 2015 se caracterizó por presentar temperaturas de hasta 13 ºC y alta abundancia de descortezadores de hasta 400 en total (Figura 2 B), la mayor abundancia registrada en esta estación se debe a las temperaturas altas, ya que en las estaciones con menor presencia de descortezadores la temperatura fue menor a 10 ºC. Los resultados obtenidos concuerdan con Sánchez-Martínez et al. (2008) y Rodríguez et al. (2010), al reportar primavera como la estación de mayor incidencia de descortezadores (n = 20 y 6.53 en promedio, respectivamente) en P. arizonica y P. hartewgii Lindl. Estos autores mencionan a otoño con alta incidencia de descortezadores, lo cual no concuerda con este estudio, ya que aquí otoño fue la estación con menor número de individuos. En el estado de Veracruz, en cambio, se observaron hasta 170 individuos de D. adjunctus en P. rudis durante la primavera (Rodríguez et al., 2013). Por su parte, Vázquez et al. (2007) mencionaron que la mayor fluctuación de D. mexicanus en Michoacán (ambos estudios en México) se presentó de marzo a junio, durante actividades de aprovechamiento, cuando los árboles fueron más susceptibles a ataques de insectos.
Fluctuación altitudinal de D. adjunctus y D. brevicomis
La prueba de Kruskal-Wallis demostró que la mayor abundancia de D. adjunctus en ambos transectos (A y B) ocurre a los 3300 msnm (n = 12.62 transecto A y n = 9.75 transecto B) (Figura 3 A y 3 C). D. brevicomis mostró un patrón similar, mayor en el transecto A, a los 3000 msnm y en el transecto B a los 3300 msnm con medias de 18.92 y 4.62 insectos, respectivamente (Figura 3 B y 3 D). La menor cantidad de insectos de ambas especies ocurrió de los 2600 a 2800 msnm. El intervalo altitudinal de 3000 a 3300 m se caracterizó por presentar alto porcentaje de la especie hospedera (P. rudis), mayor densidad del arbolado (400 a 800 árboles ha-1) en ambos transectos y alto porcentaje de mortalidad de árboles (hasta 70 % en ambos transectos) (Cuadro 2).
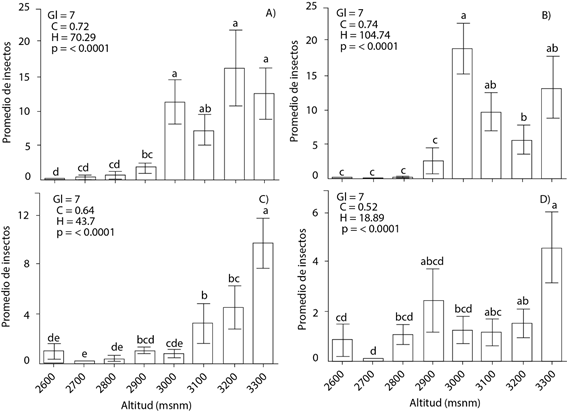
Figura 3 Prueba de Kruskal-Wallis de los valores promedio del número de insectos de Dendroctonus adjunctus (A y C) y D. brevicomis (B y D) a lo largo del gradiente altitudinal de los transectos A (A y B) y B (C y D) en Santa Rita, Arteaga Coahuila, México. Las barras indican medias (± error estándar). Medias con letras iguales (minúsculas) no son estadísticamente diferentes (Kruskal-Wallis, 0.05). Gl: grados de libertad, C: factor de corrección del estadístico por observaciones empatadas, H: estadístico de la prueba no corregido por empates; p: significancia estadística.
Cuadro 2 Características dasométricas promedio en dos transectos altitudinales (A y B) en Santa Rita, Arteaga Coahuila, México.
| Altitud | AB (m2 ha-1) | Dn (cm) | H (m) | N (ha-1) | T (°C) | M (%) | Especies de coníferas (%) | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Pm | Av | Pr | Pf † | Ps † | Pc | |||||||
| Transecto A | ||||||||||||
| 2600 | 4.89 | 27.33 | 16.56 | 72.00 | 9.66 | 16.67 | 50.00 | 27.78 | 11.11 | 11.11 | ||
| 2700 | 10.88 | 23.85 | 13.77 | 176.00 | 9.80 | 22.73 | 36.36 | 61.36 | 2.28 | |||
| 2800 | 15.97 | 28.21 | 16.36 | 216.00 | 9.78 | 48.15 | 85.19 | 5.56 | 7.40 | 1.85 | ||
| 2900 | 11.52 | 17.71 | 14.49 | 384.00 | 9.28 | 41.67 | 8.33 | 56.25 | 35.42 | |||
| 3000 | 25.58 | 25.62 | 13.93 | 420.00 | 9.14 | 54.29 | 0.95 | 23.81 | 71.43 | 3.81 | ||
| 3100 | 22.74 | 26.26 | 13.50 | 352.00 | 9.38 | 69.32 | 30.68 | 69.32 | ||||
| 3200 | 26.01 | 17.90 | 9.11 | 816.00 | 9.39 | 69.61 | 29.90 | 70.10 | ||||
| 3300 | 20.91 | 19.12 | 9.21 | 584.00 | 9.20 | 68.49 | 2.74 | 47.26 | 50.00 | |||
| Transecto B | ||||||||||||
| 2600 | 1.93 | 23.39 | 11.76 | 36.00 | 10.34 | 33.33 | 33.33 | 11.11 | 55.56 | |||
| 2700 | 14.43 | 35.52 | 17.25 | 132.00 | 10.22 | 45.45 | 81.82 | 3.03 | 15.15 | |||
| 2800 | 9.19 | 26.58 | 12.50 | 124.00 | 10.15 | 25.81 | 48.39 | 25.81 | 12.90 | 12.90 | ||
| 2900 | 19.58 | 20.53 | 10.02 | 508.00 | 9.52 | 41.73 | 28.35 | 70.08 | 1.57 | |||
| 3000 | 12.31 | 25.17 | 11.47 | 180.00 | 9.04 | 31.11 | 2.22 | 20.00 | 75.56 | 2.22 | ||
| 3100 | 21.21 | 18.77 | 12.40 | 616.00 | 8.69 | 35.06 | 11.04 | 59.09 | 27.92 | 1.95 | ||
| 3200 | 19.18 | 22.77 | 12.13 | 344.00 | 8.81 | 40.70 | 53.49 | 40.70 | 5.81 | |||
| 3300 | 48.29 | 27.03 | 17.04 | 744.00 | 7.78 | 70.97 | 29.57 | 16.13 | 54.30 | |||
AB: área basal, Dn: diámetro normal, H: Altura total, N: número de árboles, T: temperatura promedio, M: mortalidad, Pm: Pseudotsuga menziesii, Av: Abies vejarii, Pr: Pinus reflexa, Pf: Pinus flexilis, Ps: Pinus rudis, Pc: Pinus cembroides, (†): especies hospederas.
Los resultados obtenidos concuerdan con los intervalos de distribución altitudinal reportados por Salinas-Moreno et al. (2004; 2010) en México (D. adjunctus en intervalo de 3100 a 3500 msnm y D. brevicomis de 1680 a 3300 msnm). Estos autores también reportaron el mayor porcentaje de incidencias de D. adjunctus en P. hartwegii (58 a 83.3 %) y D. brevicomis en P. durangensis Martínez y P. engelmannii (31 a 40 %), lo cual no coindice con las especies hospederas de este estudio. Los brotes de D. adjunctus en la Sierra Raspadura, Chihuahua se presentaron entre 2560 y 2860 msnm, debido a la mayor distribución de especies hospederas (P. arizonica) y altas densidades de Pinus (109 árboles ha-1) (Sánchez y Silva, 2008). En Arizona, U.S.A. el rango de distribución para D. adjunctus se encuentra aproximadamente entre 2500 y 2700 msnm y D. brevicomis entre 2050 y 2250, lo cual varía con el presente estudio, debido a la ubicación en la que se hizo el muestreo en este trabajo; de la misma forma, la mayor abundancia de insectos se atribuye a mayor densidad de especies hospederas (Pinus ponderosa) y temperaturas máximas de más de 14.5 ºC (Hayes et al., 2008; Williams et al., 2008).
Relación de D. adjunctus y D. brevicomis con temperatura y variables del sitio
Las correlaciones entre la abundancia de D. adjunctus y D. brevicomis en los sitios a lo largo de los transectos fueron significativas (r > 0.70 y p < 0.03) en ambos transectos. La abundancia de ambas especies de descortezadores se correlacionó positiva y significativamente con el área basal (r > 0.70 y p < 0.05), con el número de árboles por hectárea (r > 0.76 y p < 0.04), pero negativamente con la temperatura promedio en los sitios (r > 0.60 y p < 0.08) (Cuadro 3). Los resultados obtenidos en el presente estudio son similares a los de Sánchez y Silva (2008), donde mencionan correlaciones significativas entre la abundancia de insectos con el área basal (r = 0.806; p > 0.0001). Hayes et al. (2008) demostraron que entre más área basal exista mayor será la abundancia y mayor el ataque de Dendroctonus. Los resultados obtenidos y los reportados por otros autores han demostrado que las características de los sitios de estudio (altitud, área basal, diámetro, especies hospederas y la densidad del arbolado) influyen en la fluctuación de los insectos descortezadores.
Cuadro 3 Correlación entre variables del sitio y temperatura con Dendroctonus adjunctus y D. brevicomis (n = 8, promedio de 52 colectas) en dos transectos altitudinales en Santa Rita, Arteaga Coahuila, México.
| AB (m2 ha-1) | Dn (cm) | H (m) | N ha-1 | T (°C) | D. adjunctus | ||
|---|---|---|---|---|---|---|---|
| Transecto A | |||||||
| D. adjunctus | r | 0.9048 | -0.5238 | -0.8095 | 0.9762 | -0.6429 | |
| p | 0.0167 | 0.1658 | 0.0322 | 0.0098 | 0.0890 | ||
| D. brevicomis | r | 0.8024 | -0.2515 | -0.5270 | 0.7785 | -0.8862 | 0.8264 |
| p | 0.0165 | 0.5479 | 0.1796 | 0.0229 | 0.0034 | 0.0114 | |
| Transecto B | |||||||
| D. adjunctus | r | 0.7143 | -0.4286 | -0.1190 | 0.7619 | -0.7857 | |
| p | 0.0588 | 0.2568 | 0.7528 | 0.0438 | 0.0376 | ||
| D. brevicomis | r | 0.6905 | -0.2619 | -0.3095 | 0.7857 | -0.7619 | 0.7857 |
| p | 0.0677 | 0.4884 | 0.4128 | 0.0376 | 0.0438 | 0.0376 | |
R: coeficiente de correlación de Spearman, p: significancia estadística, AB: área basal, Dn: diámetro normal promedio del sitio, H: altura total promedio del sitio, N: número de árboles, T: temperatura promedio. Nota. La abundancia usada en el análisis representa el promedio de 52 muestras comparado con el valor de las variables dasométricas y ambientales en cada sitio.
CONCLUSIONES
La mayor abundancia de D. adjunctus y D. brevicomis ocurre en primavera y verano debido a las altas temperaturas que se presentan en estas estaciones del año. La menor presencia ocurre en otoño e invierno. D. adjunctus se registró principalmente en la altitud de 3300 msnm, mientras que D. brevicomis entre 3000 y 3300 msnm, debido a las condiciones ambientales (microclima) y al número de especies hospederas que se presenta en cada altitud. La abundancia de insectos de D. adjunctus y D. brevicomis se correlacionó significativamente con el área basal, el número de árboles y la temperatura promedio del sitio. Queda de manifiesto que los brotes de descortezadores se pueden prevenir mediante tratamientos silvícolas, disminuyendo la densidad del arbolado y manejando el área basal para reducir la competencia entre las especies arbóreas existentes en el rodal.

