










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkActa botánica mexicana
On-line version ISSN 2448-7589Print version ISSN 0187-7151
Act. Bot. Mex n.110 Pátzcuaro Jan. 2015
Phenotypic variability and seed yield of Jatropha curcas L. Introducen to Tunisia
Variación fenotípica y rendimiento de Jatropha curcas L. introducida a Túnez
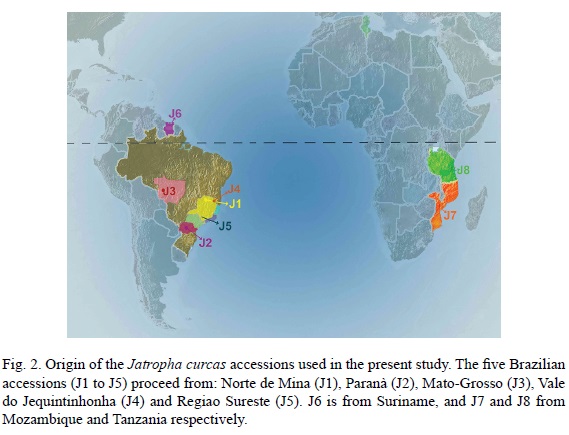
Ezzeddine Saadaoui1, José Javier Martín2, Rebeh Bouazizi1, Chokri Ben Romdhane1, Mohamed Grira3, Saad Abdelkabir3, Med Larbi Khouja4 and Emilio Cervantes2,5
1 University of Carthage, Regional Station of Gabès-Laboratory of Management and Valorization of Forest Resources, National Institute of Research in Rural Engineering, Waters and Forests (INRGREF), Tunisia.
2 Instituto de Recursos Naturales y Agrobiología de Salamanca, Consejo Superior de Investigaciones Científicas, Apartado 257, Salamanca, Spain.
3 Regional Commission for Agricultural Development, Gabès, Tunisia.
4 University of Carthage, Laboratory of Ecology (INRGREF), Tunisia.
5 Author for correspondence: ecervant@usal.es
Recibido en agosto de 2012.
Aceptado en septiembre de 2014.
ABSTRACT
Eight accessions of Jatropha curcas (six American and two African) were sown in the south of Tunisia (Gabès) to study its agricultural properties and development under local semi-arid conditions. Traits observed included general aspects of plant development (plant height, canopy circumference, number of nodes and seed yield); leaf characteristics (petiole length, limb length, limb width and foliar surface); and seed traits (size and shape). Morphological analysis of seeds included seed image area, perimeter, length/width ratio, circularity index and J index. This index reflects the similarity of seed images with an ellipse showing differences in seed shape between accessions. Jatropha curcas genotypes introduced in Tunisia are polymorphic, their seed yield is low and variable between genotypes. Seed morphology analysis gave an indication in relation with the productivity in some of the accessions. The comparative agronomic studies are crucial before early selection.
Key words: diversity, morphology of Jatropha curcas, seed shape, seed yield, Tunisia.
RESUMEN
Se sembraron en el sur de Túnez (Gabès) semillas de ocho accesiones de Jatropha curcas (seis americanas y dos africanas) para estudiar sus propiedades agrícolas y desarrollo en condiciones locales semiáridas. Los caracteres observados incluyen aspectos generales del desarrollo de planta (altura, circunferencia del follaje, número de nodos y producción de semillas); características de las hojas (longitud del peciolo, longitud del limbo, anchura del limbo y superficie foliar); y caracteres de la semilla (tamaño y forma). El análisis morfológico de las semillas incluyó el área de las imágenes, el perímetro, la relación longitud/anchura, el índice de circularidad y el índice J. Este índice refleja la semejanza de las imágenes de cada semilla con una elipse mostrando diferencias en la forma de las mismas entre accesiones. Los genotipos de J. curcas introducidos en Túnez son polimórficos, su producción de semilla es baja y variable entre genotipos. El análisis morfológico de las semillas da una idea en relación con la producción en algunas de las accesiones. Estudios agronómicos comparativos son cruciales antes de la selección.
Palabras clave: diversidad, forma de la semilla, morfología de Jatropha curca, rendimiento, Túnez.
INTRODUCTION
Jatropha curcas L. (Physic nut) is a perennial, monoecious, deciduous, stem-succulent shrub belonging to the family Euphorbiaceae. It has a smooth bark, sturdy branches, and thick papery leaves (Fig. 1). The leaves are 8 to 18 cm wide, shiny and glabrous, with exiguous and pilose stipules. The petiole is 10-16 cm long. The inflorescence is monoecious. The fruit is a capsule, 3-4 cm long and 2.5-3.0 cm wide. The seeds, 1.5-2.0 cm long and 1.0-1.2 cm wide, are elliptical and black. Dormancy is induced by fluctuations in rainfall and temperature/light. The branches contain latex. It has well-developed roots; normally five roots are formed from seedlings, one central and four peripheral. A tap root is not usually formed by vegetative propagation (Heller, 1996; Ye et al., 2009).

The genus Jatropha includes 175 species in the world (Dehgan and Webster, 1976; Ye et al., 2009). The center of diversity of Jatropha is the Mesoamerican region (Mexico and Central America), which is illustrated by the fact that more than 100 out of 175 species of Jatropha are native to that region. There are 41 native species in Mexico, of which 31 are strictly endemic (Jiménez and Vega, 2011). Several species of Jatropha are native to South America (Ovando-Medina et al., 2011a). Jatropha curcas is native of Belize, Costa Rica, El Salvador, Guatemala, Honduras, Mexico, Nicaragua and Panama, and widely present throughout Central America, Africa and Asia, where it is grown as a hedge plant (Basha et al., 2009; Subramanyam et al., 2009; Ovando-Medina et al., 2011a).
Jatropha curcas is a polymorph species with variations reported in tree architecture, plant height, number of fruits, seed dimensions (length, width, lateral diameter) and seed weight (Das et al., 2010; Rafii et al., 2012; Xu et al., 2012; Guan et al., 2013). With respect to molecular markers, the level of variation is different in localities. In America, where J. curcas is autochthonous, molecular analysis revealed high genetic variation (Ovando-Medina et al., 2011b; He, 2011; Brasileiro et al., 2013; Salvador-Figueroa et al., 2014). In contrast, assessment of J. curcas germplasm, using molecular markers in Asia and Africa, indicated low genotypic diversity and close clustering of accessions from both regions indicating a common ancestor (Basha and Sujatha, 2007; Singh et al., 2010; Zhang et al., 2011; Xu et al., 2012). This narrow level of variation has been attributed to the small number of introductions and their vegetative propagation (Khurana-Kaul et al., 2012).
Jatropha curcas has mainly evolved for xerophytic adaptation and is naturalized in the seasonally dry tropics, particularly in the southern hemisphere where there is no severe and prolonged frost (Xu et al., 2012). It grows well with 600 mm of rainfall per year or more. Deficit irrigation and salinity stress significantly reduced the growth and leaf development of greenhouse-grown Jatropha plants (Niu et al., 2012).
Conventional agriculture uses seeds and cuttings for its propagation, but J. curcas is amenable for in vitro propagation (Kumar et al., 2011). The planting of this species was motivated by the belief that it grows easily with minimal management requirements, but soil restoration and water conservation techniques improve the growth and survival rates on degraded soils (Kagamèbga et al., 2011; Iiyama et al., 2012). In plantation, spacing was the important factor along with the cutting management program for long time cultivation (Srinophakun et al., 2011). Thus, the productivity of J. curcas varies with genotypes, varieties and ecological conditions (Sosa-Segura et al., 2012).
Jatropha curcas has acquired global interest and significant economic importance for its seed oil which can be converted to biodiesel, and is emerging as an alternative to petro-diesel (Sosa-Segura et al., 2012; Guan et al., 2013). Seeds contain about 30-65% oil (Nzikou et al., 2009; Basha et al., 2009; He, 2011).
Data concerning seed yield in Jatropha curcas are heterogeneous. In Mexico it ranged from 0.2 to 12 tons per hectare, depending on production conditions (Francis et al., 2005) while in Thailand, one year old plants produced 318 g seeds x plant-1 (794 kg per hectare; Heller, 1996). In India, in a study involving sixteen genotypes, three year old plants produced from 34 to 196 g seeds × plant-1 (Das et al., 2010). Rao et al. (2008) reported yields of more than 2 kg of dry seed per five year old plant in Central India. In Egyptian deserts, Jatropha trees, irrigated with primary treated municipal wastewater, produced 2 tons seed × 4000 m-2 (Tewfik et al., 2012). In general, yield is related with the age of the plants and thus, studies on grain yield need to include plants of certain age.
Recently, eight J. curcas accessions were introduced to Tunisia with the aim of studying the possibility of Jatropha culture and exploitation. Experimental sites are installed from North to South to estimate differences between accessions or environments relative to their development and productivity. This study aims to determine the morphological variation between eight germplasm accessions of J. curcas, grown under the same environmental conditions in the experimental Dissa site in the region of Gabès (South of Tunisia), and to analyze diversity in plant development, seed morphology and seed yield. This work will contribute to identify agronomic traits of interest and to select germplasm accessions which are better adapted to local environmental conditions.
MATERIALS AND METHODS
Plant material
Eight Jatropha curcas accessions were introduced to Tunisia from four countries: Brazil, Suriname, Mozambique and Tanzania (Fig. 2).
Experimental site
This study was conducted at the Dissa experimental station (Gabès, Tunisia; 33°54' N and 10°02' E; Fig. 1A), in an area of 1000 m2, at an altitude of approximately 44 meters. This region belongs to the lower semi-arid bioclimate; mean annual rainfall averages 130 mm, average annual Potential Evapo-Transpiration (PET) is about 1400 mm. The annual average temperature is 24.5 °C, maximum and minimum temperatures are 34 °C and 12.8 °C, respectively. The soil is calcimagnesic with 15-35% of gypsum (CaSO42H2O) at depths of 50-100 cm and 15% of Calcium carbonate (CaCO3). The soil texture was drained and sandy; 85% sand, 10% clay and 5% silt. PH is 7.7 and electric conductivity EC = 2.2 mS/cm.
Water of irrigation
Plants were irrigated with treated wastewater, characterized by pH = 6.9, EC = 4.670 mS/cm, COD (Chemical Oxygen Demand) = 168 mgO2/l and BOD5 (Biochemical Oxygen Demand) = 40 mg O2/l. The rate of Cl-, Ca++, Mg++ and SO42-of treated wastewater is 39.4, 0.44, 0.32 and 450 mg × l-1 respectively. The quantity and frequency of irrigation were of 50 l × week-1 for a surface of 3 m2.
Experimental protocol, development and yield of plants
Seedlings (25 per accession) were planted in March 2008. The experiment was conducted in a randomized block with 40 plants per plot, and 5 replicates. Each accession is represented by five plants in each plot. The spacing was 2 m x 3 m. Plant height (PH) was measured for each individual every year (2008-2013) and canopy circumference (CC) was measured in 2012. Mature fruits were harvested in the three periods of fruit maturity: August 2011, August and November 2012, and January and August 2013. Seeds obtained were weighed by electronic balance (SCALTEC SBA 41) for each plant (e = 0.1 g).
Morphological parameters
Petiole length (PL), limb length (LL) and limb width (LW) were measured by graduated ruler (e = 1 mm). Foliar surface (FS) was measured by a digital planimeter, with 0.1 cm2 resolution and 0.2% precision. Measurements where mean values of five repetitions. Number of nodes (NN) was counted on five branches of 20 cm.
Seed morphology
Seeds were placed in graph paper over a flat surface, oriented with their micropile to the right side and photographed with a Nikon D60 camera adapted in a tripod. Forty seeds per accession where used to a total of 320 images. Five measurements in relation with morphology where obtained from each image: area, perimeter, length/width, circularity index and J index.
Circularity index (Schwartz, 1980) is a measure of the similarity of a plane figure to a circle given by

it ranges from 0 to 1 giving the value of 1 for circles.
An ellipse can be defined as the locus of all points that satisfy the equation
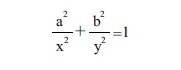
where (x, y) are the coordinates of any point on the ellipse, and (a, b) are the radius on the x and y axes respectively.
Seed images of Jatropha curcas were adjusted to an ellipse whose relation between the major and minor axes (a and b) equals the Golden Ratio (1.618). The J index was defined previously for quantification of Arabidopsis seed shape in comparison to a cardioid (Cervantes et al., 2010), and is used here to measure the adjustment of seed shape to the ellipse. To obtain the J index, composed images containing an ellipse and the seed (Fig. 3) were elaborated for each seed with the software image Photoshop CS5.1 (Adobe). Area quantification was done with Image J (Java Image Processing Program).

Quantification of the adjustment was done in each seed as a proportion between the areas in two regions: the common region in the ellipse and the seed image (area C), and the sum of total regions of both images, the seed and the ellipse (Fig. 3). The index of adjustment (J index) is defined by:
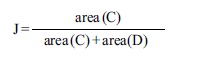
where area (C) represents the common region and area (D) the regions not shared. Note that J index ranges between 0 and 1, and decreases when the size of the non-shared region grows. It equals 1 when ellipse and seed image areas coincide, i.e., area (D) is zero. J Index was calculated for a total of 320 seeds (40 seeds per each of eight accessions).
Statistical analysis
For the eight morphological traits, the effect of the accessions was determined by the analysis of variance (ANOVA). The comparison of the averages was realized by using the test of Student-Newman-Keuls (α = 0.05). The statistical analysis also included the elaboration of a matrix of coefficient of simple linear correlation between variables. The statistical analyses were made with the software SAS 9.1 (Anonymous, 1992).
For seed morphology ANOVA was used for the comparison between different groups, post-hoc analysis was carried out using Tukey test (samples of similar sizes) and the analysis was done with SPSS 11.5.
RESULTS
Plant development and seed yield
The number of seedlings surviving varied from 11 (J8) to 24 (J3) (Table 1). After five years, mean plant height oscillated between 178 cm for J4 (Vale do Jequintinhonha) and 217 cm for J8 (Tanzanian accession) (Table 1). Canopy circumference (Table 1) oscillated between 368 (J4), and 594 cm, with higher values in the two African accessions (Mozambique, J7 and Tanzania, J8). Plants from Tanzania where higher and had larger canopy circumferences.
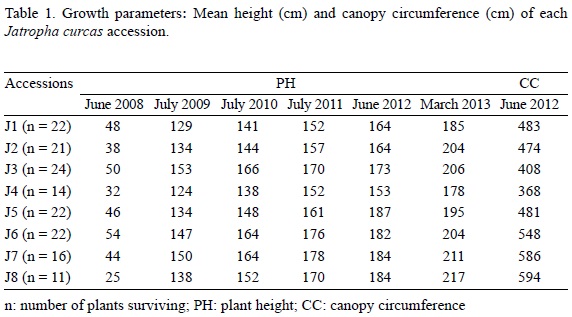
Plants from all accessions shed their leaves from December to April; the first leaves and flowers appeared in May. Seed production occurred in three periods: summer, autumn and winter. Seed yield was important in summer and winter, and limited in autumn. Only two Brazilian accessions (Norte de Mina, J1; and Mato-Grosso, J3), were productive in this season with 114 and 151 g of seeds, respectively (Table 2). Seed production was variable in the two other periods. In summer, the highest value was of 256 g (J1), and the lowest of 63 g (J7). In winter, the highest values were 140 g (J8) and the lowest ones of 64 g (J4). In total, Brazilian plants from Paranà (J2) had higher seed yield (467 g), and plants from Mozambique (J7) the lowest (158 g). Yield increased along the years of culture (Table 2).
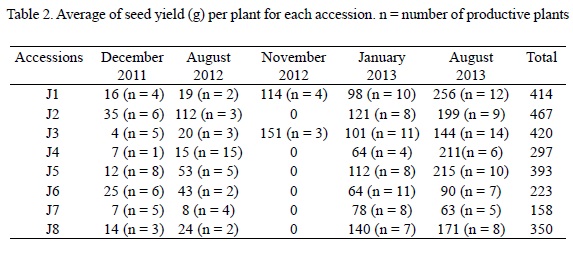
Positive correlation was found between plant height and canopy circumference (r2 = 0.87), total yield and yield of last harvest (August, 2013; r2 = 0.76), plant height and seed yield in winter (0.56), and canopy circumference and seed yield in summer (0.51) and in winter (0.62).
Morphological variability
Analysis of variance revealed high morphological variability with a significant (p < 0.05) effect of accession for all five characters under study (FS, LL, LW, PL, NN). The number of groups obtained in ANOVA differs according to the character considered (Table 3). For the parameters of the leaf, J4 forms an isolated group with higher values of LL and PL; and together with J5, both have higher values of LW and FS. All foliar parameters are lower for J7 and J3. The highest number of nods is observed for J3, and lowest for J4 (Table 3).
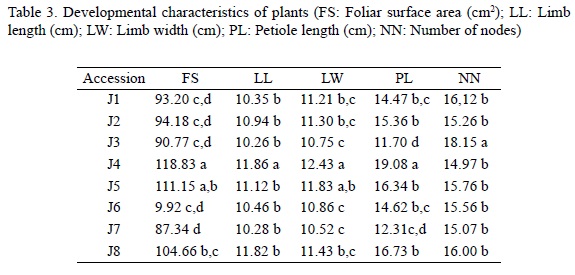
Significant (p < 0.05) positive correlations were found between all leaf magnitudes (leaf surface (FS), leaf length (LL), leaf width (LW) and petiole length (PL).
Seed morphology
Area and perimeter were lower in seed populations obtained from Suriname (J6) and Mozambique (J7) and higher in seed populations from Tanzania (J8). Length to width ratio was lower in J7 and higher in J3. Correspondingly, circularity index values were lower in J3 and higher in J7. Table 4 summarizes the results obtained for seed morphology.
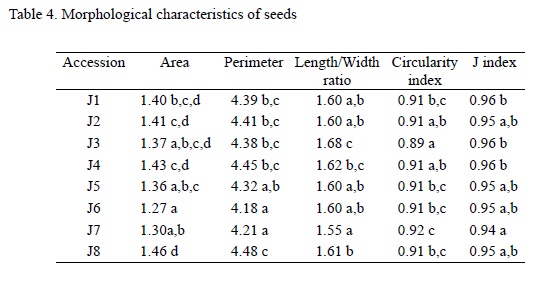
Seed shapes of Jatropha curcas adjust well to an ellipse whose relation between major and minor axes (a and b) equals the Golden Ratio (Fig. 3). The J index oscillates between 0.86 and 0.99 (mean value is 0.95). Mean values are lower in J7 and higher in J1, J3 and J4.
A negative correlation was found, as expected, between length to width ratio and circularity index (r2 = -0.97). Positive correlations were found between length to width ratio and J index (r2 = 0.76).
DISCUSSION
Yield obtained from the eight Jatropha curcas accessions used in this study was moderate or low in relationship to other published results as reported in the introduction. This may be due to the high latitude of the experimental site (33°54' N), whereas the reported limit for cultivation of this species is of 30° N (Anonymous, 2010). A cold winter season with temperature ranges below the optimum may be an important factor limiting seed production. Reduced seed yield may be a consequence of the climatic conditions during seed formation; high temperature and low moisture in summer and cold in winter, temperature being an important factor in growth and productivity of J. curcas (Maes et al., 2009). Nevertheless, grain yield increased with age of the plants, as shown by the correlation found between yield of the last harvest (August, 2013) and total yield, and further increases in yield may be expected in successive seasons.
The eight J. curcas accessions showed high phenotypic diversity, this result joins that obtained in America (Ovando-Medina et al., 2011b; He, 2011; Brasileiro et al., 2013), China, India and Philippines (Gohil and Pandya, 2009; Vijayanand et al., 2009; Das et al., 2010; Zapico et al., 2011; Guan et al., 2013). However, high morphological variability does not always reflect high genotypic variability. Xu et al. (2012) showed that high phenotypic diversity in most cases was not associated with genotypic diversity, indicating a strong influence for environment. It appears that the environment or particular genotype-environment combinations have predominant roles in the morphological variation among accessions (Ovando-Medina et al., 2011a; Rafii et al., 2012). Our results show high variability in foliar traits, in particular leaf surface. Plant height and seed yield are also different between accessions, this result due probably to genetic-environmental interaction (Heller, 1996). Epigenetics also plays an important role in the variability of Jatropha (Yi et al., 2010).
Previous reports using unique varieties present a relation between seed yield and agronomic traits like foliar surface, plant height, number of nodes per branch, and others (Gohil and Pandya, 2009; Rafii et al., 2012; Brasileiro et al., 2013). For example, Sun et al. (2012) identified a total of 28 QTLs (quantitative trait loci) underlying the growth and seed yield traits in Jatropha and showed a significant correlation between phenotypic traits and total seed weight. Plant height, stem diameter, branch number, female flower number and fruit number were suggested to be key factors for seed yields. Rafii et al. (2012) showed the important heredity of plant height and indicated that the genetic component was high (> 50%) for seed yield per plant, fruit × plant-1 and flowering bunches × plant-1. Brasileiro et al. (2013) used plant height for indirect selection in populations of the physic nut in order to obtain superior genotypes. Our data obtained with plants of eight different accessions grown during five years do not support the existence of general correlations between any of the considered morphological traits and grain yield. Plants in this experiment proceed from diverse origins and thus, genotype-environment interactions are multiple and may result in a lack of general correlation between any particular morphological character of the plants and yield, a property that may well be observed when considering a unique variety or accession on its own. In addition, the morphological traits were measured in a season previous to the increase in grain yield that took place in the last harvest of August 2013.
The images of Jatropha curcas seeds adjust well to an ellipse, whose relation between major and minor axis equals the Golden Ratio. Differences between accessions were found in seed size and shape, showing a trend where larger seeds had also higher values of J index and lower circularity index. Our results suggest a relationship between seed yield , size and shape. Seeds from the accessions J6 and J7 (Suriname and Mozambique respectively), that presented the lowest yield in the last seasons, were smaller and more circular, and their shape adjusted less well to an ellipse (lower J index values). The observed association between reduced yield and seed morphological traits opens the possibility for the development of tests addressed to evaluate the quality of seed lots, based on morphological properties of seeds. Seed shape analysis is simple to perform and may provide an indication on the condition of the seed stocks in relation to yield. This may contribute to germplasm selection for agronomical applications.
The introduction of J. curcas is a complex project; the programmes launched in Brazil, Nicaragua and India for introduction of J. curcas for varied purposes met with limited success due to the wide gap in potential and actual yields. The species has not been improved for productivity and most of the projects relied on naturally occurring unadapted populations, which are a result of the few initial introductions (Basha and Sujatha, 2007). Thus, Jatropha curcas is not common in Mediterranean regions with arid and semi-arid climates; plantations in these regions hold the risk of low productivity or irrigation requirement. Also, plantations in regions with frost risk hold the risk of damage due to frost (Maes et al., 2009). However, the high phenotypic variability and the difference in productivity represented a good potential for early selection. Proposed future research in this species comprises the description of introduced genotypes, the study of the physiological behavior and the improvement of seed yield and oil productivity.
LITERATURE CITED
Anonymous. 2010. Jatropha: A small holder bioenergy crop. ICM 8: 27-53. [ Links ]
Basha, S. D. and M. Sujatha. 2007. Inter and intra-population variability of Jatropha curcas (L.) characterized by RAPD and ISSR markers and development of population-specific SCAR markers. Euphytica 156: 375-386. [ Links ]
Basha, S. D., G. Francis, H. P. S. Makkar, K. Becker and M. Sujatha. 2009. A comparative study of biochemical traits and molecular markers for assessment of genetic relationships between Jatropha curcas L. germplasm from different countries. Plant Sci. 176: 812-823. [ Links ]
Brasileiro, B. P., S. A. Silva, D. R. Souza, P. A. Santos, R. S. Oliveira and D. H. Lyra. 2013. Genetic diversity and selection gain in the physic nut (Jatropha curcas). Genet. Mol. Res 12(3): 2341-2350. [ Links ]
Cervantes E., J. J. Martín, R. Ardanuy, J. G. de Diego and A. Tocino. 2010. Modeling the Arabidopsis seed shape by a cardioid: efficacy of the adjustment with a scale change with factor equal to the Golden Ratio and analysis of seed shape in ethylene mutants. J. Plant Physiol. 167: 408-410. doi:10.1016/j.jplph.2009.09.013 [ Links ]
Das, S, R. C. Misra, A. K. Mahapatra, B. P. Gantayat and R. K. Pattnaik. 2010. Genetic variability, character association and path analysis in Jatropha curcas. World Appl. Sci. J. 8(11): 1304-1308. [ Links ]
Dehgan, B. and G. Webster. 1979. Morphology and infrageneric relationships of the genus Jatropha (Euphorbiaceae). Univ. Cal. Publ. Bot. 74: 1-73 + 33 plates. [ Links ]
Francis, G., R. Edinger and K. Becker. 2005. A concept for simultaneous wasteland reclamation fuel production, and socio-economic development in degraded areas in India, potential and perspectives of Jatropha plantations. Nat. Resour. Forum 29(1): 12-24. [ Links ]
Gohil, R. H. and J. B. Pandya. 2009. Genetic evaluation of Jatropha (Jatropha curcas Linn.) genotypes. J. Agric. Res. 47(3): 221-228. [ Links ]
Guan, J., H. Yu, J. Zhang, R. Yang and Y-H. Fan. 2013. Study on seed morphology and genetic diversity of Jatropha curcas L. from different provenances. Afr. J. Food Sci. Technol. 5(2): 169-173. [ Links ]
He, W. 2011. Biochemical and genetic analyses of Jatropha curcas L. Seed composition. PhD Thesis. University of York. York, UK. 149 pp. [ Links ]
Heller, J. 1996. Physic Nut - Jatropha curcas L. Promoting the conservation and use of underutilized and neglected crops. 1. International Plant Genetic Resources Institute. Rome, Italy. Available from: URL: http://www.ipgri.cgiar.org/publications/pdf/161.pdf. [ Links ]
Iiyama, M., D. Newman, C. Munster, M. Nyabenge, G. W. Sileshi, V. Moraa, J. Onchieku, J. G. Mowo and R. Nadass. 2012. Productivity of Jatropha curcas under small holder farm conditions in Kenya. Agroforest. Syst. 87: 729-746. [ Links ]
Jiménez Ramírez, J. and K. Vega Flores. 2011. Jatropha mirandana (Euphorbiaceae), especie nueva de la cuenca oriental del río Balsas de los estados de Guerrero y Puebla, México. Novon 21(2): 192-195. [ Links ]
Kagamèbga, W. F., A. Thiombiano, S. Traoré, R. Zougmoré and J. I. Boussim. 2011. Survival and growth responses of Jatropha curcas L. to three restoration techniques on degraded soils in Burkina Faso. Ann. For. Res. 54(2): 171-184. [ Links ]
Khurana-Kaul, V., S. Kachhwaha and S. L. Kothari. 2012. Characterization of genetic diversity in Jatropha curcas L. germplasm using RAPD and ISSR markers. Indian J. Biotechnol. 11: 54-61. [ Links ]
Kumar, N., K. G. V. Anand and M. P. Reddy. 2011. Plant regeneration of non-toxic Jatropha curcas-impacts of plant growth regulators, source and type of explants. J. Plant Biochem. Biotechnol. 20(1): 125-133. [ Links ]
Maes, W. H., A. Trabucco, W. M. J. Achten and B. Muys. 2009. Climatic growing conditions of Jatropha curcas L. Biomass Bioenerg. 33: 1481-1485. [ Links ]
Niu, G., D. Rodriguez, M. Mendoza, J. Jifon, G. Ganjeguntel. 2012. Responses of Jatropha curcas to salt and drought stresses. International Journal of Agronomy IJA, Article ID 632026, 7 pages. doi:10.1155/2012/632026 [ Links ]
Nzikou, J. M., L. Matos, F. Mbemba, C. B. Ndangui, N. P. G. Pambou-Tobi, A. Kimbonguila, Th. Silou, M. Linder and S. Desobry. 2009. Characteristics and composition of Jatropha curcas oils, variety Congo-Brazzaville. Res. J. Appl. Sci. Eng. Tech. 1(3): 154-159. [ Links ]
Ovando-Medina, I., F. J. Espinosa-García, J. S. Núñez-Farfán and M. Salvador-Figueroa, M. 2011a. State of the art of genetic diversity research in Jatropha curcas. Sci. Res. Essays 6(8): 1709-1719. [ Links ]
Ovando-Medina, I., A. Sánchez-Gutiérrez, L. Adriano-Anaya, F. Espinosa-García, J. Núñez-Farfán and M. Salvador-Figueroa. 2011b. Genetic diversity in Jatropha curcas populations in the State of Chiapas, Mexico. Diversity 3: 641-659. [ Links ]
Rafii, M. Y., I. W. Arolu, M. H. A. Omar and M. A. Latif. 2012. Genetic variation and heritability estimation in Jatropha curcas L. population for seed yield and vegetative traits. J. Med. Plants Res. 6(11): 2178-2183. [ Links ]
Rao, G. R., G. R. Korwar, A. K. Shanker and Y. S. Ramakrishna. 2008. Genetic associations, variability and diversity in seed characters, growth, reproductive phenology and yield in Jatropha curcas (L.) accessions. Trees Struct. Funct. 22(5): 697-709. [ Links ]
Salvador-Figueroa M., J. Magaña-Ramos, J. A. Vázquez-Ovando, M. L. Adriano-Anaya and I. Ovando-Medina. 2014. Genetic diversity and structure of Jatropha curcas L. in its centre of origin. Plant Genet. Resour.-Charact. Util. 150: 1-10. doi:10.1017/S1479262114000550 [ Links ]
Schwartz, H. 1980. Two-dimensional feature-shape indices. Mikroskopie (Wien) 37(Suppl): 64-67. [ Links ]
Singh, L. P., S. Singh, S. P. Mishra and S. K. Bhatia. 2010. Molecular characterization of Genetic Diversity in Jatropha curcas. Genes, Genomes, Genomics 4(1): 1-8. [ Links ]
Sosa-Segura, M. P., M. A. Angulo-Escalante, J. B. Valdez-Torres, J. B. Heredia, T. Osuna-Enciso, R. Allende-Molar and B. D. Oomah. 2012. Phenology, productivity, and chemical characterization of Jatropha curcas L. as tool for selecting non-toxic elite germplasm. African J. Biotech. 11(93): 15988-15993. [ Links ]
Srinophakun, P., A. Saimaneerat, I. Sooksathan, N. Visarathanon, S. Malaipan, K. Charernsom and W. Chongrattanameteekul. 2011. Integrated research on Jatropha Curcas plantation management. World Renewable Energy Congress - Sweden. 8-13 May. Linköping, Sweden. http://dx.doi.org/10.3384/ecp11057232 (accessed 11/4/2014). [ Links ]
Subramanyam, K. D. Muralidhararao and N. Devanna. 2009. Genetic diversity assessment of wild and cultivated varieties of Jatropha curcas (L.) in India by RAPD analysis. Afr. J. Biotech. 8(9): 1900-1910. [ Links ]
Sun, F., P. Liu, J. Ye, L. C. Lo, S. Cao, L. Li, G. H. Yue and C. M. Wang. 2012. An approach for Jatropha improvement using pleiotropic QTLs regulating plant growth and seed yield. Biotechnol. Biofuels. 5: 42. doi: 10.1186/1754-6834-5-42. [ Links ]
Tewfik, S. R., S. I. Hawash, N. K. Atteya, G. I. El Diwani and I. H. Farag. 2012. Techno-economic appraisal of biodiesel from Jatropha curcas: An Egyptian case study. J. Agric. Sci. Technol. B 2(3B): 287-297. [ Links ]
Vijayanand, V., N. Senthil, S. Vellaikumar and M. Paramathma. 2009. Genetic diversity of Indian Jatropha species as revealed by morphological and ISSR markers. J. Crop Sci. Biotech. 12(3): 115-120. [ Links ]
Xu, W, S. Mulpuri and A. Liu. 2012. Genetic diversity in the Jatropha genus and its potential application. CAB Reviews 7(59): 1-15. doi: 10.1079/PAVSNNR20127059 [ Links ]
Ye, M., C. Li, G. Francis and H. P. S. Makkar. 2009. Current situation and prospects of Jatropha curcas as a multipurpose tree in China. Agroforest. Syst. 76: 487-497. [ Links ]
Yi, C., S. Zhang, X. Liu, H. TN Bui and Yan Hong. 2010. Does epigenetic polymorphism contribute to phenotypic variances in Jatropha curcas L.? BMC Plant Biol. (10): 259. doi:10.1186/1471-2229-10-259 [ Links ]
Zapico, F. L., S. K. Nival, C. H. Aguilar and M. N. Eroy. 2011. Phenotypic diversity of Jatropha curcas L. from diverse origins. J. Agric. Sci. Technol. 5(2): 215-219. [ Links ]
Zhang, Z., X. Guo, B. Liu, L. Tang and F. Chen. 2011. Genetic diversity and genetic relationship of Jatropha curcas between China and Southeast Asian revealed by amplified fragment length polymorphisms. Afr. J. Biotech 10(15): 2825-2832. doi: 10.5897/AJB10.838 [ Links ]

