











 text new page (beta)
text new page (beta) English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO 
 Permalink
PermalinkIntroduction
Plant growth promoting rhizobacteria (PGPR) are microorganisms capable of promoting plant growth through different mechanisms, such as improving nutrient absorption and mobility (Yang et al., 2009), antibiotic production, nitrogen fixing, plant hormone production, phosphate solubilization, siderophore production and biological control (Bashan and de-Bashan, 2005). The use of PGPR is an alternative for sugarcane (Saccharum spp.) nutrition since it is a crop that demands large quantities of nitrogen (N), varying from 120 to 320 kg ha-1 (Salgado et al., 2003). Brazil uses around 50 kg ha-1, because normally PGPR's are applied in the fertilization (Oliveira et al., 2006). The PGPR associated mainly with gramineous plants are Azospirillum lipoferum, A. brasilense and A. amazonense, Herbaspirillum seropedicae, Acetobacter diazotrophicus, Enterobacter agglomeran, E. cloacae, Bacillus azotofixans, B. polymyxa, Alcaligenes faecalis, Klebsiella sp., Azotobacter and Pseudomonas (Loredo-Osti et al., 2004). Plant response to inoculation with several soil microorganisms has been attributed to the inoculum's ability to synthesize and excrete substances in the rhizosphere, including diverse plant growth regulators such as auxins, gibberellins, cytocinins, ethylene and absicic acid (ABA) (Tilak et al., 2006). Of the bacteria isolated in the rhizosphere, 86% produce auxins and other growth regulators, although this varies widely among species and strains of the same species. It is also affected by conditions of cultivation, growth stage and substrate availability (Vestergard et al., 2009). Moreover, microorganisms can participate in inorganic phosphate solubilization and in organic phosphate mineralization, as well as in its immobilization (Pérez et al., 2007). The microbial mechanisms for solubilizing phosphate consist of phosphatase enzyme activity and synthesis of organic acids, such as citric, butyric, malonic, lactic, succinic, malic and gluconic acids, among others, produced by diverse bacteria known as phosphate solubilizing bacteria (PSB), which make phosphate available for absorption by plants (Paredes-Mendoza y Espinosa-Victoria, 2010).
With the aim of implementing environment-friendly techniques that tend to reduce chemical fertilizer use in sugarcane production, the effect of 24 plant growth promoting bacterial strains on the commercial sugarcane variety MEX 69290 was assessed.
Materials and methods
Biological Material
The Molecular Plant-Microbe Interaction Laboratory of the Colegio de Postgraduados, Mexico, provided twenty-four bacterial strains isolated from sugarcane. The strains belong to the species Aeromonas salmonicida, Burkholderia cepacia, Ochrobactrum anthropi, Pseudomonas sp., Shewanella putrefaciens, Sphingomonas paucimobilis and Stenotrophomonas maltophilia.
For the bioassay, 25 day-old sugarcane seedlings of the variety MEX69290 produced in vitro were used. The seedlings were obtained from the Biotechnology Laboratory at the Motzorongo sugar factory in the state of Veracruz, México.
Determination of Total Indoles
A suspension (10% v/v) of each bacterial strain was inoculated in liquid Luria Bertani (LB) culture medium supplemented with tryptophan (0.1 g L-1). The cultures were incubated at 28 °C for 48 h and later centrifuged at 5000 rpm for 10 min (Ghevariya and Desai, 2014). The supernatants were mixed with Salkowski reagent (600 mL de H2SO4 18 M, 4.5 g FeCl3 anhydrous gauged to 1 L with distilled water) in a 1:1 proportion. To estimate the concentration of total indoles, the supernatants were incubated at room temperature in darkness for 30 min, and absorbance at 539 nm was then read in a Genesys 20 spectrophotometer (Hartmann et al., 1983).
Determination of Phosphate Solubilization
The bacterial strains were cultivated in 50 mL Pikovskaya medium (Espinosa-Victoria et al., 2009) and incubated at 28 °C with constant shaking (160 rpm) for 5 days. Then, the medium was centrifuged for 15 min at 5000 rpm. One milliliter of supernatant was taken and three mL distilled water and 1 mL of the reagent nitro vanadate-molybdate (yellow complex) were added. Absorbance was read at 470 nm after 20 min of incubation. Phosphate solubilization, in terms of the concentration of soluble phosphates in the medium, was determined using a standard KH2PO4 curve (Jeon et al, 2003).
Experimental Unit, Transplant and Inoculation
The experimental unit was one plastic pot with 500 g of a mixture of clay soil from sugarcane field and agrolite in a 1:1 proportion. The clay soil collected from farm plots of the Colegio de Postgraduados, Campus Córdoba, Veracruz, was dried at room temperature in the shade. Later, it was sieved through a 2 mm mesh and autoclaved two times at 120 °C for 3 h. A sample of the farm soil was taken to make the following determinations: pH, water retention capacity, organic matter, total nitrogen, assimilable phosphorus, extractable potassium and texture (Benedicto-Valdés et al., 2005).
The 24 bacterial strains were propagated in nutrient broth. The culture media were incubated at 32 °C under shaking (220 rpm) for 24 h (Ghevariya and Desai, 2014). One milliliter of the bacterial culture (1 X 107 UFC) was used to inoculate each plant at the base of its stem.
Variables Assessed and Sampling
The assessed response variables were height, stem diameter, number of tillers, leaf area, and aerial and root dry biomass. Leaf area was determined with an area integrator (LI-COR-Biosciences). Dry biomass was estimated at constant weight in dry samples, which were maintained for 72 h at 70 °C. Sampling was destructive, and five plants of each treatment were used. Data of the different variables in sugarcane plants were taken 15, 30, 45, 60 and 75 days after inoculation (DAI).
Experimental Design and Data Analysis
The experiment was conducted in a greenhouse during 90 days. The experimental design was completely random distribution with 25 treatments (24 bacterial strains and one un-inoculated control) and 5 replications.
An analysis of variance and a test of comparison of means were performed on the data. The latter was done with the LCD test (P ≤ 0.05) using the statistical software SAS (Statistical Analysis System Institute, 2009) for all of the variables.
For the case of estimations in growth analyses, logarithmic transformation was done for the respective test of means.
To determine the leaf area index (LAI), the following equation was used: LAI= (LA/NP)*DP/10000 cm2, where LA is leaf area (cm2), NP is the number of sampled plants, and DP is the number of plants per m2. Also, net assimilation rate (NAR), relative growth rate (RGR) and absolute growth rate (AGR) were calculated using the following equations: NAR=[(PS2-PS1)/(LA2-AF1)]/ [(lnLA2-lnLA1)/(T2-T1)], RGR=(lnPS2-lnPS1)/(T2-T1) and AGR=(PS2-PS1)/(T2-T1). PS2 and PS1 correspond to plant dry weight at times 2 and 1 (T2 and T1); and lnLA2 and lnLA1 are the natural logarithm of the leaf area at two times (Hunt, 2003).
Results and discussion
Table 1 shows the production of total indoles of the 24 bacterial strains used. The values range from 0.11 to 117.34 μg mL-1 total indoles, corresponding to the species Stenotrophomonas maltophilia 46 and Pseudomonas luteola IMPCA244, respectively. The highest values are important indicators that surpass the values of indole production (2 to 49.66 mg mL-1) reported in other studies (Rana et al., 2011; Carcaño-Montiel et al., 2006). The species Pseudomonas luteola IMPCA244, Ochrobactrum anthropi IMP311 and Ochrobactrum anthropi N208 were statistically different (P ≤ 0.05) from the other 21 strains.
Table 1 Total indoles production and phosphate solubilization by 24 bacterial species used to inoculated sugarcane seedlings.
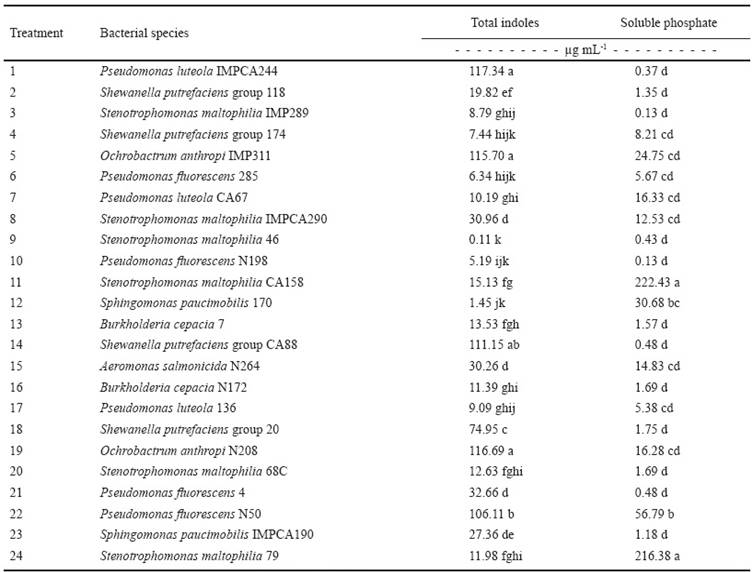
Different letters in the same column are statistically different (LCD; P ≤ 0.05).
Phosphate solubilization, expressed as soluble phosphates, of the 24 evaluated strains ranged from 0.13 to 222.43 μg mL-1 (Table 1); these two values corresponded to the bacteria Pseudomonas fluorescens N198 and Stenotrophomonas maltophilia CA158, respectively.
Sugarcane is predominantly cultivated in acid soils. In this study, pH was 5.8. For this reason, it is common to observe serious limitations in the supply of phosphorus available for the crop, underlining the importance of rhizobacteria that make available insoluble forms of P found in this type of soil.
Several studies have demonstrated that diverse microorganisms (Glickman and Dessaux, 1995) carry out in vitro synthesis of total indoles and other physiologically active growth regulators derived from tryptophan. The production of total indoles by a microorganism depends on several factors, such as the bacterial strain, the host plant, the culture conditions, crop development stage and the concentration of microorganisms in the substrate (Inui-Kishi et al., 2010).
Among the growth plant regulators produced by the bacteria, indolacetic acid (which forms part of total indoles) is one of the most important. It is widely distributed among cultivated plants, causing morphological changes in the root and directly regulating absorption of some minerals (Dobbelaere et al., 2002).
Table 2 shows that plants inoculated with Pseudomonas luteola 136, Aeromonas salmonicida N264, Stenotrophomonas maltophilia IMP289 and Stenotrophomonas maltophilia 79 were statistically different and superior in terms of plant height, relative to the other treatments 30, 45, 60 and 75 DAI. These species had a total indole production of 9.09, 30.26, 8.79 and 11.98 μg mL-1, respectively, and for phosphate solubilization, values of 5.38, 14.83, 0.13 y 216.38 μg mL-1 of soluble phosphate, respectively. The results show that plant height is not directly related to the quantity of indoles produced or phosphate solubilized by the bacterial species. Nevertheless, it was observed that the bacteria that promoted greater plant height were those that synthesized less than 30 μg mL-1 total indoles.
Table 2 Height and diameter of sugarcane seedlings inoculated with 24 bacterial species.
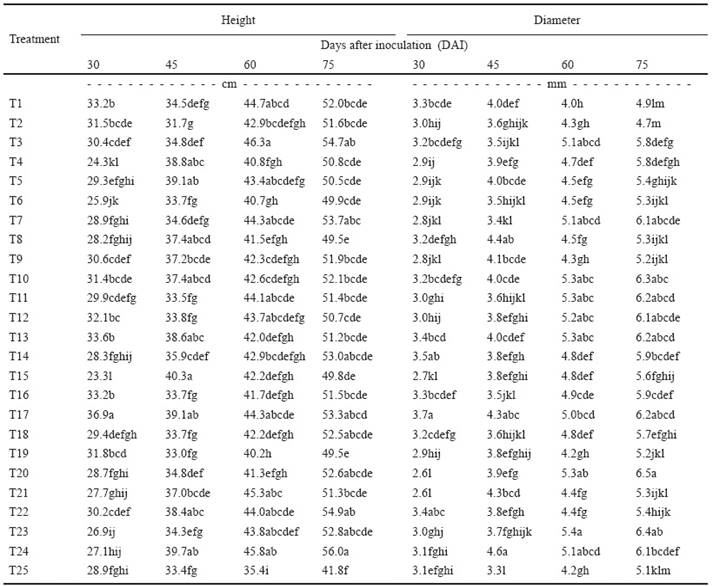
Different letters in the same column are statistically different (LCD; P ≤ 0.05). T1-T24: Bacterial species inoculated (See Table 1). T25: Control (not inoculated).
Like the results for plant height, Pseudomonas luteola 136 and Stenotrophomonas maltophilia 79 promoted significantly superior plantlet stem diameter in comparison to the rest of the treatments at 30 and 45 DAI. On days 60 and 75 after inoculation, the species Sphingomonas paucimobilis IMPCA190 and Stenotrophomonas maltophilia 68C had higher values for stem diameter than the other species; they synthesized 27.36 y 12.63 μg mL-1 total indoles, respectively. Regarding soluble phosphate, the same species had values of 1.18 and 1.69 μg mL-1, respectively. Again, it is observed that it is not necessarily the bacteria with the highest values of total indole synthesis and phosphate solubilization that induce larger stem diameter in seedlings.
The number of tillers of sugarcane plantlets (Table 3) is an important variable since in the field this datum is essential for crop yield projections. The species Aeromonas salmonicida N264 promoted the largest number of tillers 30 and 45 DAI, with 30.26 total indoles and 14.83 μg mL-1 phosphate solubilization. At 60 and 75 DAI the species Burkholderia cepacia N172 and Ochrobactrum anthropi IMP311 induced a larger number of tillers than the other species, with values of 11.39 and 115.701 μg mL-1 total indoles and 1.69 and 24.75 μg mL-1 soluble phosphate, respectively.
Table 3 Number of tillers and leaf area of sugarcane seedlings inoculated with 24 bacterial species.
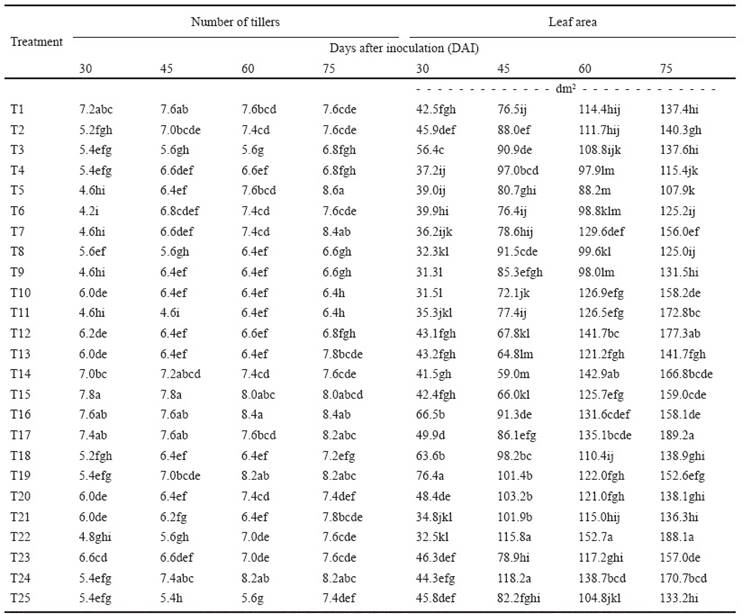
Different letters in the same column are statistically different (LCD; P ≤ 0.05). T1-T24: Bacterial species inoculated (See Table 1). T25: Control (not inoculated).
Leaf area is another important parameter (Table 3) since the size ofthe photosynthetic apparatus determines crop yield. The species Ochrobactrum anthropi N208 promoted the largest leaf area 15 DAI, with total indoles and phosphate solubilization values of 116.69 and indoles and 56.79 μg mL-1 soluble phosphate. This species was the most consistent in inducing the largest leaf area during the bioassay, relative to the other bacterial species and had high values of total indole synthesis and phosphate solubilization.
Aerial dry matter weight (sum of leaf area weight plus main stem and tiller weight) is highly relevant since it reflects the effect of PGPR on development of sugarcane plants.
Table 4 shows that Shewanella putrefaciens group 20 promoted more aerial dry matter 30 DAI with total indole values of 74.955 μg mL-1 and 1.75 μg mL-1 of soluble phosphate. At 45 DAI, Stenotrophomonas maltophilia 79 was statistically superior in aerial dry matter induction with values of total indoles and soluble phosphates of 11.989 and 216.38 μg mL-1, respectively.
Table 4 Aerial and root dry matter of sugarcame inoculated with 24 bacteral species.
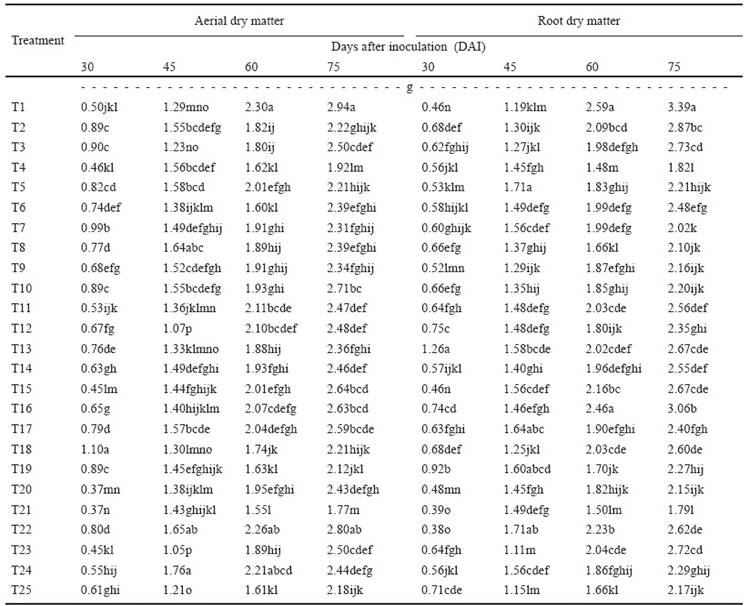
Different letters in the same column are statistically different (LCD; P ≤ 0.05). T1-T24: Bacterial species inoculated (See Table 1). T25: Control (not inoculated).
The dry weight of the root is an important indicator, reflecting the quantity of effective root for plant nutrient and water absorption. For this reason, indoles and soluble phosphates are of vital importance for plant development. Table 4 shows that Burkholderia cepacia 7 promoted a greater effect on root growth of the sugarcane seedlings at 30 DAI, with values of total indoles and soluble phosphates of 13.53 and 1.57 μg mL-1, respectively. Ochrobactrum anthropi IMP311, induced better results in root dry weight 46 DAI, with synthesis of total indoles and soluble phosphates at values of 115.70 and 24.75 μg mL-1, respectively. Finally, Pseudomonas luteola IMPCA244 promoted higher values of root dry weight in comparison with the rest of bacterial strains at 60 and 74 DAI. It is important to emphasize that some Burkholderia cepacia strains have been recognized as a human opportunistic pathogen in hospitals and associated with cystic fibrosis (Ibarguren et al., 2011).
The leaf area index (LAI) revealed significant permanent increases on the sampling dates (Table 5). However, the PGPR inoculated treatments had variable increases on different dates, while the control showed constant increases with no large variations.
Table 5 Leaf Area Index (LAI) of sugarcane seedlings inoculated with five strains of bacterial species.

T1, T17, T19, T22 y T24 = strains of four bacterial species (See Table 1). T25 = non inoculated plants.
In the case of net assimilation rate (NAR) and relative growth rate (RGR), the effect of inoculation with Pseudomonas luteola IMPCA244 was the highest and most constant, although on the early dates the effect of Stenotrophomonas maltophilia 79 is outstanding, but it declines later on. Absolute growth rate (AGR) had behavior similar to NAR and RGR, although the effect of inoculation with Pseudomonas fluorescens N50 showed a constant trend, as observed in Figure 1.
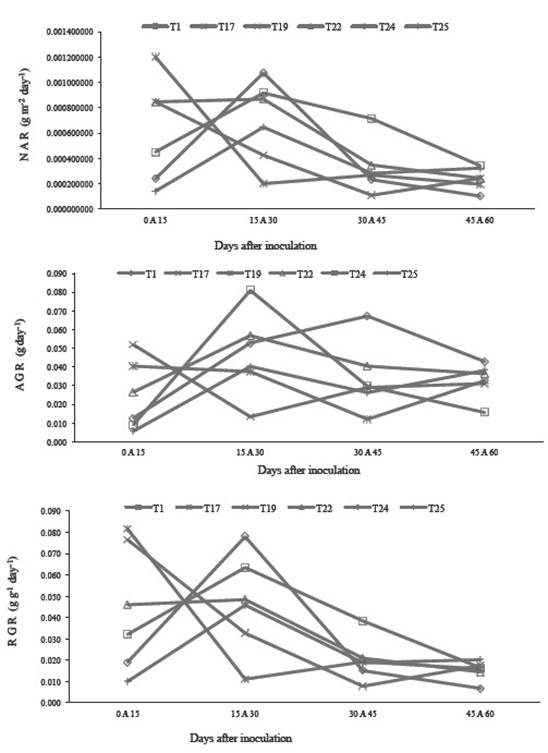
Figure 1 Dynamics of the Net Assimilation Rate (NAR), Absolute Growth Rate (AGR) and Relative Growth Rate (RGR) in sugarcane plants inoculated with five bacterial strains: T1, T17, T19, T22 and 24 = strains of four bacterial species (See Table 1). T25 = control, non-inoculated plants.
Conclusions
The species Ochrobactrum anthropi (strains IMP311 and N208) and Pseudomonas luteola IMPCA244 were statistically superior in total indole production. The species Stenotrophomonas maltophilia 79 was that with the highest values of phosphate solubilization.
The species Pseudomonas luteola IMPCA244, Ochrobactrum anthropi IMP311, Aeromonas salmonicida N264, Burkholderia cepacia N172, Pseudomonas fluorescens N50 and Stenotrophomonas maltophilia 79 outstandingly promoted different response variables in sugarcane throughout the bioassay.
The sugarcane plants inoculated with the species Pseudomonas luteola (strains IMPCA244 and 136), P. fluorescens N50, Ochrobactrum anthropi N208 and Stenotrophomonas maltophilia 79 had the highest leaf area indexes and growth rates.
The results of this study indicate that the gains in sugarcane plant height are not necessarily related to the amount of indoles synthesized or to phosphates solubilized by the bacterial species used for inoculation. Nevertheless, it is observed that the bacteria that promoted greater plant height synthesized less than 30 μg mL-1 total indoles.
Before using some of the species evaluated in this research in the production of biofertilizers for sugarcane, the absence of human pathogenic genes in their genomes must be corroborated. In the same way, antibiosis studies are required to determine the feasibility of using them individually or in consortiums. Particularly, the use of Burkholderia cepacia N172 is not recommended unless it can be proved that it does not carry in its genome genes that are pathogenic for humans.

