











 text new page (beta)
text new page (beta) Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO 
 Permalink
PermalinkIntroducción
La estimación de las propiedades ópticas de las hojas a partir de mediciones del follaje de diferentes tipos de vegetación, es importante para relacionarlo con las concentraciones de los constituyentes bioquímicos de las hojas, particularmente en el ámbito de la aplicación de la tecnología de sensores remotos. Esta tecnología, a nivel de campo o a bordo de satélites, permite la utilización de métodos no destructivos para la estimación de parámetros biofísicos o bioquímicos, asociados a los datos espectrales medidos de la vegetación u hojas. No obstante su atractivo, la aplicación operacional de los sensores remotos ha sido poco exitosa y plagada de aproximaciones empíricas que requieren la tarea constante de calibrar los métodos de cálculo para las estimaciones. La estimación de los constituyentes bioquímicos de las hojas ha sido explorada a través de métodos estadísticos (Jacquemoud et al., 1995b); los resultados obtenidos muestran correlaciones buenas entre los datos espectrales y la bioquímica, sin embargo, estas no son congruentes con las propiedades radiativas de las hojas, además de ser poco estables (Grossman et al., 1996). Ante esta situación se ha planteado el desarrollo de modelos radiativos semi-teóricos que asocien las mediciones ópticas de las hojas con la bioquímica. El modelo PROSPECT (Jacquemoud y Baret, 1990; Jacquemoud et al., 1996) ha sido desarrollado para estos fines y es actualmente uno de los más usados. El modelo PROSPECT utiliza la concentración de la clorofila a y b, del agua (espesor equivalente) y de la biomasa seca (proteína + celulosa + hemicelulosa + azucares + almidones + lignina, principalmente); además de un parámetro relativo a la estructura del mesófilo foliar.
Las mediciones de reflectancias por sensores remotos ha sido utilizada para el desarrollo de numerosos índices espectrales de vegetación o IV, tanto a nivel de hojas (le Maire et al., 2004) como de follaje (Broge y Leblanc, 2000). Los resultados de este enfoque han sido poco satisfactorios y poco estables fuera del conjunto de calibración. En el caso particular de la determinación de la bioquímica foliar en el follaje, la inversión de modelos radiativos acoplados de las hojas y del follaje también ha sido poco exitosa (Jaquemoud et al., 1995a), algo similar ocurre en el caso del uso de los IV (Broge y Leblanc, 2000).
Recientemente, usando el marco teórico de los invariantes espectrales (Huang et al., 2007), Lewis y Disney (2007) han demostrado en términos generales que dado el acoplamiento de las propiedades estructurales del follaje (índice de área foliar o IAF y cobertura o fv) con la bioquímica foliar no es posible realizar estimaciones robustas de ambas propiedades, sin importar lo angosto de las bandas espectrales usadas o el número de ellas. Cualquier estimación que se realice de ambas propiedades contendrá un error muy grande y representará un problema matemáticamente indeterminado (muchas soluciones al mismo problema). Así, la perspectiva de la estimación de las propiedades bioquímicas de las hojas usando datos espectrales del follaje de la vegetación parece tener una limitación fundamental e intrínseca.
El entendimiento de los patrones de los espacios espectrales asociados a las mediciones de reflectancias de la vegetación, sensores remotos o radiómetros de campo, permite el desarrollo de aplicaciones prácticas y robustas de esta tecnología en la estimación de la biomasa aérea y rendimientos de los cultivos (Pascual et al., 2012), cobertura de la vegetación o índice de área foliar (Paz et al., 2007 y 2011; Reyes et al., 2011), estrés en cultivos (Solorio et al., 2008; Reyes et al., 2011), entre otras aplicaciones. Para que el uso de la información espectral sea útil, dado que lo que se mide es función de una gran cantidad de variables, es necesario el desarrollo de esquemas de modelación genéricos, pero simples, que permitan obtener datos biofísicos con el mínimo de información (Paz et al., 2009c). Las bases de esta aproximación son discutidas en este trabajo, bajo diferentes perspectivas.
El objetivo principal de este trabajo fue demostrar que la limitación definida por Lewis y Disney (2007), puede ser evitada usando el concepto de medio equivalente y espacios n-paramétricos, que hacen operacional el problema de convertir un medio heterogéneo (problema de acoplamiento estructura- bioquímica) a uno homogéneo (medio turbio en términos radiativos), donde el factor estructural es desacoplado del bioquímico. La propuesta, desarrollada a partir del planteamiento realizado por Paz et al. (2009a y c), es ejemplificada con dos experimentos clásicos (maíz y algodón) de mediciones radiativas en cultivos, además de simulaciones radiativas. La presentación de simulaciones y datos de campo de experimentos controlados permite revisar la congruencia del marco teórico desarrollado, además de posicionar los problemas experimentales asociados a las mediciones en campo.
Medio Radiativos Equivalentes
Dejando fuera el problema asociado a la geometría sol-sensor (función de distribución bidireccional de la reflectancia y su contraparte geométrica de la distribución de las plantas en una parcela o píxel) se introduce el concepto de medio radiativo equivalente, como un esquema operacional a la limitación asociada a la imposibilidad de conocer, en primera aproximación, los elementos constituyentes de un píxel en una imagen satelital, sin recurrir a hipótesis que no pueden ser validadas directamente.
En una parcela o rodal, el IAF estimado experimentalmente representa uno global:
Sin considerar efectos de borde, el IAF1 o local (plantas individuales) esta relacionado con el global como:
donde fv es la fracción de vegetación y está dado en proporción de 0 a 1. Así, la fracción de cobertura del suelo es fs = 1-fv.
En la Figura 1 se muestra el IAF1, donde la fracción de huecos (fh) dentro del límite del follaje (proyección horizontal) estará dado por la Ley de Beer-Lambert, para hojas no negras (Goudriaan y van Laar, 1994):
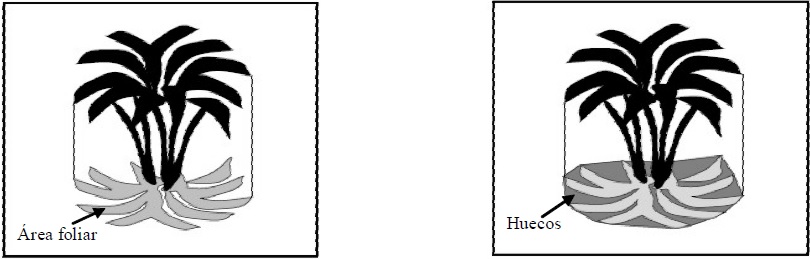
Figura 1 Área foliar y huecos para el caso de un medio local o planta individual (adaptada de BLM, 1999).
donde: Tl es la transmitancia (local) de la radianza directa solar, ω = ρ + τ es el albedo foliar (suma de la reflectancia y transmitancia foliar, respectivamente) y K es un coeficiente de extinción. Nótese que fv ≠ fh, ya que fv se refiere a la fracción de vegetación fuera de los límites de las plantas individuales; aunque en la práctica (fotografías digitales) fv incorpora a fh.
En la Figura 2 se muestra el caso de un cultivo sembrado en surcos, donde al principio de la etapa de crecimiento se observa un patrón de plantas individuales, donde las plantas no se tocan entre sí (caso 3-D). Al avanzar el crecimiento, las plantas en un mismo surco se tocan entre si y forman un cultivo en hileras o surcos (caso 2-D). Cuando las plantas de diferentes surcos se tocan, tenemos la situación de cobertura total del suelo (caso 1-D). Esta dinámica de crecimiento requiere de una estrategia genérica de modelación que tome en cuenta, razonablemente, la complejidad geométrica de los cultivos en parcelas y la vegetación natural.
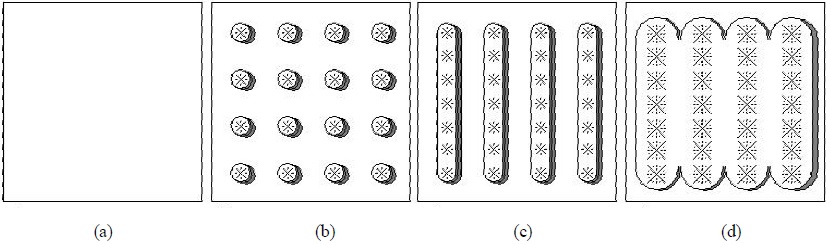
Figura 2 Geometría asociada a un cultivo sembrado en surcos. (a) suelo desnudo; (b) plantas individuales sin traslape; (c) plantas traslapadas en surcos; (d) vegetación densa traslapada.
De la discusión anterior, un medio equivalente (términos físicos) es aquel que cumple:
Es decir, para una medición de IAFG hay un número prácticamente infinito de combinaciones equivalentes de IAFl y fv que satisfacen la relación (4). En términos ópticos, un medio radiativo equivalente cumple la relación (4), dado que el IAFl y fv están en función de las propiedades ópticas del suelo y los fitoelementos de la vegetación (dejando lo demás fijo). En la Figura 3 se muestra en forma esquemática el concepto de medio equivalente, donde las reflectancias obtenidas para todos los medios mostrados son iguales, generando un problema de indeterminación a nivel sub-píxel. El medio que muestra una distribución al azar de los fitoelementos se le denomina medio turbio en términos radiativos (Ross, 1981).

Los medios radiativos equivalentes deben cumplir la condición de que sus dimensiones 3-D (voxéles) deben ser tales, que las interacciones horizontales de fotones entre pixeles contiguos sean mínimas o nulas (Pinty et al., 2004). Esto se cumple generalmente en cultivos agrícolas, matorrales y pastizales y requiere de evaluarse en el caso de bosques y selvas.
Dinamica espectral y espacios n-paramétricos
Para analizar la dinámica espectral del crecimiento de los cultivos en diferentes espacios n-paramétricos, se utilizaron simulaciones radiativas de un modelo de medio turbio uni-dimensional (Verhoef, 1984), detalladas en Paz et al. (2005), y dos experimentos de campo con cultivos con arquetipos contrastantes: los datos fueron tomados de Bausch (1993) para maíz y Huete et al. (1985) para algodón. En ambos experimentos, se realizaron mediciones de reflectancia durante la etapa de emergencia y vegetativa hasta reproductiva, en cultivos sin estrés, utilizando charolas deslizantes con diferentes suelos debajo de los cultivos, para simular suelos debajo de los cultivos con propiedades ópticas diferentes, debido a textura, materia orgánica, humedad, rugosidad y óxidos de fierro, principalmente. Estos experimentos, así como las mediciones radiométricas y biofísicas asociadas están detalladas en Paz et al. (2007 y 2011). En el experimento de maíz el ángulo cenital solar durante toda la campaña de muestreo varió de 17.2 a 24.2°. En el caso del experimento de algodón, las variaciones fueron de 22 a 31.7°. Considerando que las variaciones de la geometría sol-sensor fueron mínimas y que ambos experimentos muestran una ventana de condiciones de iluminación más o menos similares, no se hizo ningún intento de estandarizar la geometría sol-sensor de éstos.
Espacio R-IRC (n = 0-paramétrico)
En la Figura 4 se muestra el espacio n = 0-paramétrico, de la banda espectral del rojo (R) e infrarrojo cercano (IRC) asociado a la dinámica del crecimiento de la vegetación, representada por el índice de área foliar, donde se usaron cuatro suelos con propiedades ópticas (reflectancias) diferentes. En esta figura las líneas iso-IAF (IRC = a0 + b0R) representan la misma cantidad de vegetación, pero con suelos diferentes; donde el caso de IAF = 0 representa la línea del suelo desnudo (IRC = aS + bSR). Si el suelo (reflectancia) permanece constante, entonces el crecimiento de la vegetación sigue una curva iso-Suelo. El atrayente de la dinámica del crecimiento de la vegetación para la banda del R es el punto de convergencia de todas las curvas iso-Suelo. En este punto la banda del R se satura (no cambia de valor al incrementarse el IAF) y representa a un medio ópticamente denso o infinito (Ross, 1981), por lo que se le denomina como R∞. El valor del IRC asociado al R de saturación es IRCR∞. La banda del IRC sigue creciendo hasta alcanzar su propio punto de saturación definido por IRC∞.

Las reflectancias en el infinito están en función de las propiedades ópticas de las hojas y su distribución angular, y la geometría sol-sensor, principalmente (Ross, 1981; Zarco et al., 2008; Paz et al., 2009a). Las concentraciones de los constituyentes bioquímicos foliares están en función de las propiedades ópticas de las hojas (reflectancia y transmitancia) (Jacquemoud et al., 1996), por lo que determinadas éstas es posible realizar estimaciones desacopladas de los parámetros estructurales del follaje, puesto que la reflectancia en el infinito es invariante por efecto del IAF o fv (los valores superiores de estos parámetros no cambian las reflectancias en el infinito).
Los patrones lineales iso-IAF mostrados para el caso uni-dimensional también son válidos para el caso de simulaciones radiativas tridimensionales (Gao et al., 2000) y cultivos reales con mediciones de campo (Paz et al., 2007 y 2011). El rompimiento de la linealidad (polinomio de segundo grado) se presenta cuando los valores de las reflectancias de los suelos son altos (tipo desiertos). En el caso de los espacios del espectro visible (azul o A, verde o V y rojo) con el del IRC tiene patrones similares a los mostrados en la Figura 4 (Paz et al., 2005), producto de la linealidad de los patrones de las reflectancias de los suelos que impone una condición de frontera lineal en la solución de la ecuación de transferencia de fotones en el sistema suelo-vegetación. Así, los resultados mostrados en lo siguiente son generalizables para los espacios A-IRC y V-IRC. En el caso de los infrarrojos medios, los resultados son aplicables pero con una simetría no lineal, producto de que los patrones de las reflectancias de los suelos son no lineales (Paz et al., 2009b).
En la misma Figura 4 se muestran la proyección de las líneas iso-IAF hasta la intersección con la línea del suelo (Ris, IRCis), definida por:
Espacio a0-b0 (n = 1-paramétrico)
Los parámetros de las líneas iso-IAF del espacio R-IRC, Figura 4, forman un espacio n = 1-paramétrico con patrones específicos. La Figura 5a muestra el espacio a0-b0 para las simulaciones radiativas de la Figura 4, donde se observa un patrón exponencial para valores bajos del IAF y para valores altos el patrón es lineal. Entre ambos patrones hay valores asociados a un IAF de transición, que implica un cambio en las pendientes de la curva exponencial a la lineal. Esto queda más claro si utilizamos la transformación 1/b0 y la analizamos en relación a a0 (Figura 5b). En este caso se observan dos patrones aproximadamente lineales. Cuando b0 tiende al valor de la pendiente asociada a la línea recta del ápice de la Figura 4 (reflectancias en el infinito), entonces 1/b0 tiende a cero (Figura 5b).

El patrón de la Figura 5b ha sido usado para el diseño del índice espectral NDVIcp, donde los casos del maíz y algodón discutidos se conforman muy bien al patrón observado para las simulaciones radiativas (Paz et al., 2007).
Es conveniente analizar los patrones temporales del IAF asociado a los dos cultivos bajo análisis. La Figura 6 muestra la evolución del IAF en el tiempo para el maíz y algodón, donde se observa al inicio de la emergencia del cultivo un patrón exponencial, seguido por uno lineal, dentro de la etapa vegetativa. Posteriormente, al inicio de la etapa reproductiva se presenta un patrón tipo exponencial que termina hasta el IAF máximo. La fase exponencial y lineal de la etapa vegetativa puede caracterizarse con un modelo expo-lineal (Ferrandino, 1989; Goudriaan y Monteith, 1990) y generalizarse a uno expo-lineal-exponencial (Goudriaan, 1994) para modelar en forma completa los patrones mostrados en la Figura 6. Estos modelos tienen bases biofísicas relacionados con la Ley de Beer-Lambert (hojas negras, es decir, hojas que absorben todos los fotones que chocan contra ellas) y la eficiencia de conversión de radiación solar a área foliar o biomasa aérea fueron desarrollados bajo la consideración de medios turbios. En esta situación idealizada, los patrones exponencial y lineal de la Figura 5a y 6 se corresponden, por lo que se observa que un espacio espectral puede proyectarse a un espacio temporal. No obstante esto, para cultivos sin competencia extrema entre sí, la fase expo-lineal en tiempo del IAF aproxima la fase exponencial hasta un poco más de la transición a la lineal del espacio a0-b0 y la fase exponencial de la etapa de reproducción del IAF se aproxima a los inicios de la fase lineal del espacio a0-b0; aunque los patrones espectrales entre un medio turbio y uno heterogéneo o tri-dimensional son similares, las tasas de cambio son diferentes. Esto se discute más adelante, particularmente para valores más allá del punto de transición del patrón exponencial al lineal.

Espacio dIRC-R (n = 0-paramétrico)
Las reflectancias en el infinito están relacionadas con las propiedades ópticas foliares de interés, por lo que es necesario estimar primero sus valores. Para lograr este objetivo es necesario hacer lineales las curvas iso-suelo (Figura 4) usando el espacio transformado dIRC-IRC (Paz et al., 2006), donde dIRC = IRC- (aS + bSR). En este espacio las líneas iso-IAF (IRC = a1 + b1dIRC) están dadas por (Paz et al., 2005 y 2006):
El problema del espacio R-IRC es que al aproximar las reflectancias del infinito, las líneas iso-IAF tienen pendientes que tienden a infinito. En el espacio dIRC-IRC, cuando b0 tiende a infinito, b1 tiende a 1 (línea de 45°), aunque permanece el problema de pendiente con valor infinito para la línea del suelo (Figura 7).

Se observa en la Figura 7 que después de un punto de transición t (transición de la etapa exponencial a la lineal mostrada en la Figura 5a), las pendientes de las líneas iso-IAF son indistinguibles del valor 1, tanto a nivel de simulaciones (Figura 7a), como de mediciones en campo (Figura 7b). En la fase exponencial del crecimiento de los cultivos, en el espacio espectral dIRC-IRC, podemos aproximar las curvas iso-Suelo con líneas rectas (IRC = r1 + s1dIRC) (Figura 7). Las líneas iso-suelo se interceptan en el punto (dIRCt, IRCt) y después de este punto hasta el punto de saturación del R (dIRCR∞, IRCR∞), región de la transición exponencial-lineal de la Figura 5a, las pendientes de las líneas iso-Suelo se aproximan a 1, por lo que resultan difícil de distinguir de esta pendiente. Esto es consecuencia de los valores relativos entre dIRC e IRC. En el caso de mediciones en campo, de reflectancias de los cultivos, donde la fase lineal es muy corta (espectralmente hablando), se puede utilizar la aproximación: dIRCt = dIRCR∞ y IRCt = IRCR∞ (Paz et al., 2008a) (Figura 7b).
El problema del espacio dIRC-IRC es que el valor de IRC∞ no es accesible antes o después de la fase exponencial (Figura 7), ya que no se conoce cuando será el valor máximo de la línea a 45 grados después de esta fase.
Espacio a1-ß (n = 1-paramétrico)
La ventaja del espacio dIRC-IRC es que permite plantear la relación entre los parámetros a1 y b1 de las líneas de iso-IAF en un formato donde la pendiente va de un valor de 90° (suelo desnudo) a 45° (vegetación ópticamente densa). Así, Paz et al. (2005 y 2006) propusieron la transformación de b1 a un formato estandarizado:
En la Figura 8 se muestran los patrones del espacio n = 1-paramétrico a1-ß, tanto para las simulaciones radiativas como para las mediciones en campo.
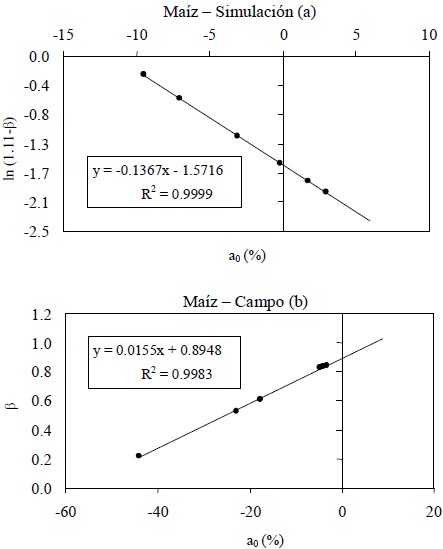
En el caso de las simulaciones radiativas, la relación entre a1-ß es del tipo exponencial, Figura 8a, donde el valor 1.11 usado fue optimizado para el modelo radiativo aplicado (Paz et al., 2005). En el caso de las mediciones en campo, el patrón es lineal (Figura 8b). Esta diferencia de patrones es producto del desarrollo corto o largo de la fase lineal en la etapa vegetativa, tal como se discutió anteriormente. En el caso de las simulaciones radiativas, el patrón exponencial puede aproximarse por dos patrones lineales que coinciden, aproximadamente, en el punto de transición de la fase exponencial a la lineal (Romero et al., 2009). No obstante esta situación, el punto critico es que las tasas de cambio espectrales tienen diferencias entre el caso de medios homogéneos (turbios) y heterogéneos (campo). Esto explica algunas inconsistencias discutidas más adelante.
Problemática asociada a las mediciones espectrales de cultivos en campo
Las mediciones de reflectancias en los cultivos de maíz y algodón, usando fondos de suelos deslizables, dentro del error experimental, presentan algunos problemas. En la Figura 9a para el maíz (similar al caso del algodón), las curvas iso-Suelo muestran un inicio con valores menores del R e IRC a los esperados, provocando que los patrones similares a los mostrados en la Figura 4 tengan valores de reflectancia mayores para el suelo, por lo que al desechar los datos del suelo desnudo (como se hizo) se recuperan los patrones. No está clara una explicación de esta situación, aunque probablemente sea efecto de sombreado y rugosidad del suelo.
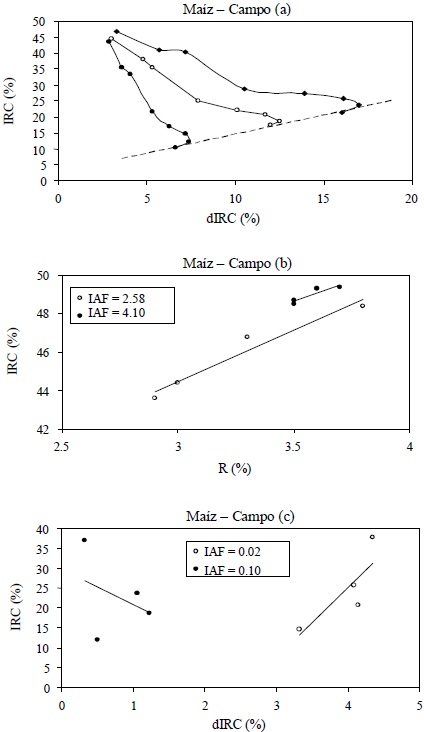
En el caso del espacio R-IRC, las líneas iso-IAF para valores grandes del IAF muestran errores importantes (Figura 9b) y para el caso de IAF para los valores pequeños los ajustes son buenos (no mostrados). Para el espacio dIRC-IRC la situación se revierte, donde los líneas iso-IAF para valores grandes del IAF tienen buenos ajustes (no mostrados), pero para valores pequeños del IAF los ajustes son malos (Figura 9c). Estas tendencias en los errores se propagan en los análisis de los espacios n = 1-paramétricos de estos espacios espectrales.
De la discusión anterior, los parámetros a0 y b0 (y otros) de los cultivos agrícolas analizados fueron estimados usando una combinación de los espacios espectrales discutidos y sus transformaciones, para contar con información completa.
Espacio IVPP-R (n = 0-paramétrico)
Como se comentó anteriormente, la estimación de IRC∞, así como las propiedades ópticas y concentraciones bioquímicas asociadas, son imposibles en el espacio del dIRC-IRC. Otra alternativa para hacer lineales las curvas iso-Suelo es usar el espacio espectral IVPP-R (Paz et al., 2008a), donde IVPP = dIRC/IRC (Figura 10).
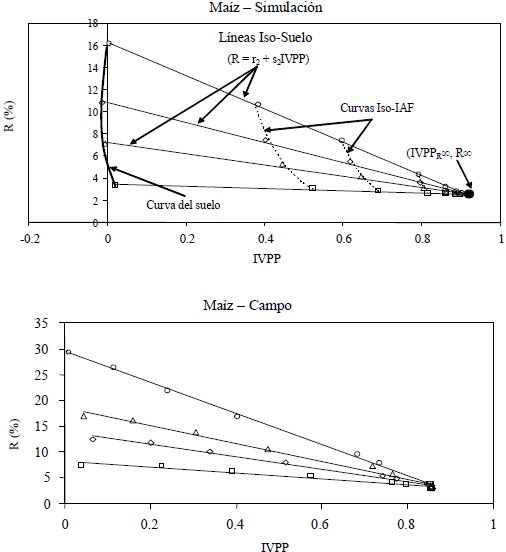
La ventaja del espacio IVPP-R es que solo considera la perspectiva de la banda del R y está acotado por el límite de los patrones iso-Suelo del espacio R-IRC hasta el punto (R∞, IRCR∞) de la Figura 4, ya que el formato del IVPP reduce los efectos de las contribuciones arriba de este valor (hacia la saturación de la banda del IRC).
En el espacio IVPP-R, las líneas iso-IAF son curvas no lineales (Figura 10) contrastando con la linealidad de las curvas iso-Suelo (R = r2 + s2IVPP). Las curvas iso-Suelos representan, en el espacio del R-IRC, un modelo lineal con interacciones:
Definido (IVPPR∞, R∞) entonces podemos estimar los valores del IRC asociados:
Las relaciones (10) evitan el problema de la hipótesis dIRCt = dIRCR∞ y IRCt = IRCR∞ del uso del espacio del dIRC-IRC y permiten una estimación directa del punto (R∞, IRCR∞) de la Figura 4.
Espacio s1-r1 (n = 1-paramétrico)
En la Figura 11 se muestra el patrón entre los parámetros s1-r1 de las líneas del suelo del espacio dIRC-IRC.

La relación mostrada en la Figura 11 está definida por:
La cual es consecuencia de la propiedad de un conjunto de líneas rectas que tienen un punto de intersección común.
En la Figura 12 se muestra el patrón entre los parámetros s2-r2 de las líneas del suelo del espacio IVPP-R. La relación mostrada en la Figura 12 está definida por:
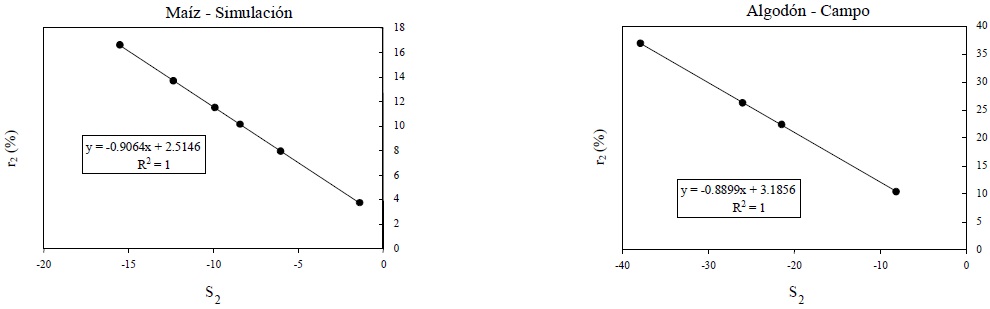
la cual es también consecuencia de la propiedad de un conjunto de líneas rectas que tienen un punto de intersección común.
Espacio Rs-Rsv e IRCs-IRCsv (n = 0-paramétrico)
Para medios homogéneos (medios turbios), la relación entre la reflectancia de la mezcla suelo-vegetación, puede aproximarse considerando solo interacciones de primer orden de los fotones (chocan contra el suelo una sola vez) (Huete, 1987):
donde: T se refiere a la transmitancia en la banda espectral particular, Rs e IRCs son las reflectancias del suelo (huecos en la vegetación), Rv e IRCv son las reflectancia de solo la vegetación (Rs = 0 o condición de frontera de suelo negro) y Rsv e IRCsv son las reflectancias del sistema suelo-vegetación (medio turbio).
En el caso de medios heterogéneos (no turbios), podemos usar la aproximación de una mezcla lineal entre las reflectancias del suelo desnudo y de la mezcla suelo-vegetación (medio turbio):
donde Rv e IRCv se refiere a la reflectancia de la mezcla suelo desnudo-vegetación (medio turbio, con una fracción fh de suelo en los huecos de la vegetación, relación (3).
La determinación de las reflectancias de la mezcla suelo-vegetación (medio turbio) pueden aproximarse usando la solución de un sistema de ecuaciones tipo Kubelka-Munk (Bonhomme y Varlet, 1977; Goudriaan, 1977; Ross, 1981) para radiación difusa (valida también para radiación total), bajo la condición de frontera de suelo negro:
De las relaciones (12) y (13) se puede establecer (Yoshiaka et al., 2000):
que puede plantearse en función de las reflectancias en el infinito usando las relaciones (14):
Para el caso de medios turbios, las relaciones (16) se revierten a las (13), fv = 1.
Usando la relación IRCsv = as + bsRsv (mezcla suelo desnudo y suelo-vegetación) de la línea del suelo, podemos sustituir las relaciones (15) en esta relación (Yoshiaka et al., 2000) para obtener la relación que define los parámetros de las líneas iso-IAF del espacio R-IRC y utilizar las relaciones (14) para plantearlas en función de las reflectancias en el infinito (accesibles en términos experimentales y de sensores remotos):
Las Ecuaciones (5), (6), (7), (8), (16) y (17) definen las relaciones entre los diferentes espacios analizados en este trabajo, las cuales pueden usarse en forma simultánea para evaluar la congruencia y estimaciones de los diferentes espacios. La ventaja de la parametrización de las relaciones (17) es que los parámetros de las líneas iso-IAF, espacio del R-IRC, están definidas en función de los parámetros de las líneas del suelo (no de los valores específicos de las reflectancias) y de las transmitancias solo de la vegetación (el suelo desnudo está incorporado en el término fv), accesibles a nivel experimental (además de las reflectancias en el infinito determinadas de acuerdo a los procedimientos experimentales introducidos en Paz et al., 2008a).
En la Figura 13 se muestran los patrones del espacio Rs-Rsv y en la Figura 14 los del espacio IRCs-IRCsv, los cuales implican relaciones lineales de acuerdo a las ecuaciones (12), (15) o (16) definidas previamente.


La hipótesis de interacciones de orden uno de las reflectancias y el modelo lineal de las ecuaciones (13) explican los comportamientos observados (medios equivalentes).
En el caso del espacio Rs-Rsv, las líneas iso-IAF de la Figura 13 pueden ser representadas usando la relación Rsv = t1 + u1Rs y para el espacio IRCs-IRCsv (Figura 14) por IRCsv = t2 + u2IRCs.
Espacio u1-t1 (n = 1-paramétrico)
La Figura 15 muestra el patrón entre los parámetros u1 y t1 de la relación lineal entre Rs-Rsv, para las simulaciones radiativas.

De la relación (16) para Rsv, los parámetros u1 y t1 pueden definirse en términos de fv y TR, por lo que podemos ponerlos en términos de fv e igualar los resultados para obtener:
El patrón de la Figura 15 es congruente con esta parametrización, que aunque este espacio nos permite obtener R∞ en forma alternativa al espacio IVPP-R, este último es más estable.
En la Figura 16 se presentan los patrones del espacio u1-t1 para los dos experimentos de campo analizados.
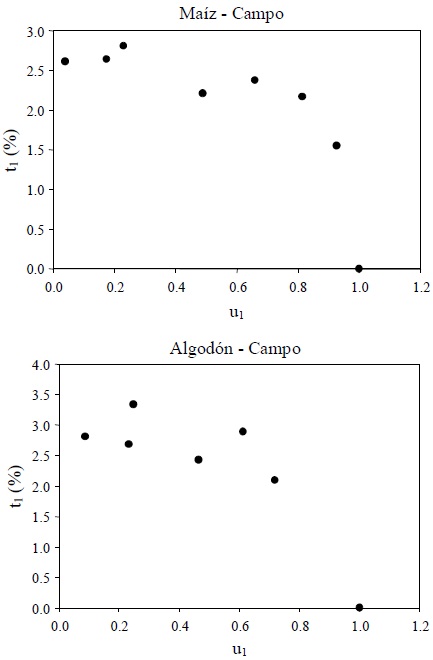
Los patrones observados en la Figura 16 son consecuencia de los problemas asociados a las mediciones de las reflectancias del R (poca variación) y pueden aproximarse por un modelo polinómico de segundo orden.
Espacio u2-t2 (n = 1-paramétrico)
En la Figura 17 se muestra el patrón del espacio u2-t2, asociado al modelo lineal entre IRCs e IRCsv, para las simulaciones radiativas.

El modelo polinómico de segundo orden de la Figura 17 puede parametrizarse usando las condiciones de frontera u2=1, t2=0 y u2=0, t2=IRC∞, para quedar como:
que es consecuencia de la fase lineal larga de la etapa vegetativa modelada en las simulaciones radiativas.
En el caso de las mediciones de campo de reflectancias, fase lineal corta, la Figura 18 muestra los patrones entre u2-t2.

En la Figura 18 se ha ajustado un modelo lineal para la etapa vegetativa y otro para la reproductiva con buenos ajustes, que es congruente con el modelo lineal implicado por la relación (18):
No obstante esto, en el caso del maíz, el IRC∞ asociado a la etapa vegetativa no es congruente con los resultados del espacio dIRC-IRC. Así, en la Figura 19 se muestra el resultado del ajuste de un solo modelo lineal para los datos de la etapa vegetativa y reproductiva, después de eliminar el dato para IAF = 0.58 del maíz, dato que rompe patrones en varios espacios analizados y que probablemente tenga un error experimental grande de medición; algo similar sucede con el dato de IAF = 0.31.
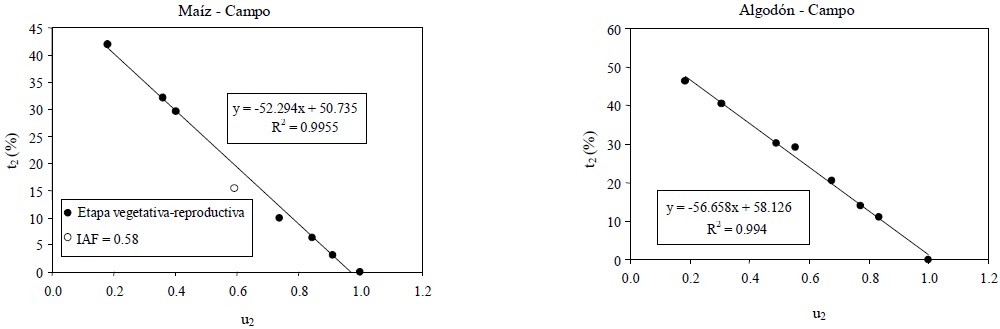
Figura 19 Patrones del espacio u2-t2 (n = 1-paramétrico), experimentos de campo y un solo modelo para las etapas vegetativa y reproductiva.
Las implicaciones de usar un solo modelo lineal para la etapa vegetativa y reproductiva, considerando que las reflectancias son función de las propiedades ópticas y angulares de las hojas (ver sección siguiente), son que los cultivos mantienen un patrón invariante de ambas propiedades durante el crecimiento hasta el valor del IAF máximo (Casiano et al. 2012), donde la etapa reproductiva con propiedades ópticas diferentes a la vegetativa conserva el patrón global al cambiar sus propiedades angulares; esto en términos de medios equivalentes.
Espacio IRCis-t2 (n = 1-paramétrico)
En el caso del patrón polinómico entre u2 y t2 para las simulaciones radiativas, podemos generar un espacio alternativo que lo plantee en forma lineal: IRCis-t2 (Figura 20).
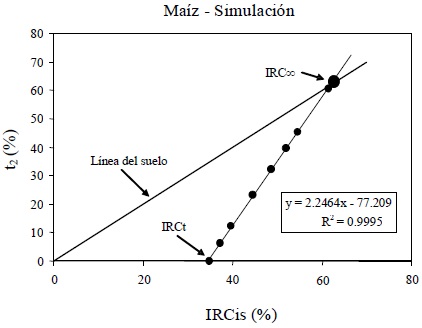
El modelo lineal de la Figura 20 está dado por:
por lo que considerando que la línea del suelo mostrada en la Figura 20 tiene pendiente unitaria e intersección cero, la intersección de ambas líneas definirá el punto IRC∞:
Se observa en la Figura 21 los problemas asociados a la propagación de errores de las mediciones de campo, discutidos anteriormente.

Espacios Supra-Paramétricos (n = 2-paramétrico) y mayores
Los espacios meta-paramétricos espectrales (n = 1-paramétrico) fueron introducidos por Paz et al. (2005 y 2006) para el espacio a1-β (A-B). En lo general, si consideramos cualquier espacio meta-paramétrico entre los parámetros A y B, podemos ajustar un modelo lineal, en forma directa o transformando el espacio, del tipo:
Los parámetros AA y BB del espacio meta- paramétrico definen un solo punto en el espacio supra-paramétrico AA-BB y representa la dinámica del crecimiento asociado a un medio ambiente y manejo particular (Paz et al., 2009c). En el caso de los espacios espectrales, representan la trayectoria espectral del crecimiento bajo la consideración de un número infinito de condiciones iniciales, representadas por distintas propiedades ópticas del fondo de la vegetación.
Los patrones en el espacio AA-BB (diferentes medios ambientes y manejos) definen un patrón genérico para cada cultivo o vegetación, representando en forma muy compacta una especie de “genotipo” que tiene asociado un número infinito de “fenotipos” (dinámicas del crecimiento asociadas a los espacios meta-paramétricos para un medio ambiente y manejo particular) (Solorio et al., 2008; Paz et al., 2009c; Reyes et al., 2011). La Figura 22 esquematiza el espacio supra-paramétrico discutido.

En un espacio superior (n = 3-paramétrico), AA = AAA + BBB (BB), podemos comparar diferentes tipos de cultivo o vegetación en forma extremadamente compacta, permitiendo su caracterización asociada al espacio de análisis particular (espectral, temporal, angular, etc.) (Solorio et al., 2008; Paz et al., 2009c; Reyes et al., 2011).
Conclusiones
En este trabajo se introdujo un marco metodológico para la estimación de las reflectancias en el infinito usando diferentes espacios n-paramétricos, conversión de medios heterogéneos a homogéneos, bajo la restricción de medios equivalentes que usan parámetros efectivos de las propiedades biofísicas y bioquímicas asociadas a la geometría 3-D de la vegetación. Este marco teórico (aproximado) permite resolver el problema de acoplamiento entre las propiedades estructurales y bioquímicas de la vegetación planteado por Lewis y Disney (2007), que imposibilita la estimación conjunta de ambas propiedades. La solución planteada es totalmente operacional.
Con la estimación de las reflectancias en el infinito, atrayentes de la dinámica espectral asociada al crecimiento de la vegetación, podemos usar la teoría de transferencia radiativa de medios turbios u homogéneos para estimar las propiedades ópticas (equivalentes) del follaje de la vegetación y poder hacer estimaciones de las concentraciones de los componentes bioquímicos, usando una aproximación semi-teórica (Jacquemoud y Baret, 1990; Jacquemoud et al., 1996) o semi-empírica (Gitelson et al., 2003).

