










Services on Demand
Journal
Article
 text in
text in  English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkCiencias marinas
Print version ISSN 0185-3880
Cienc. mar vol.49 Ensenada Jan./Dec. 2023 Epub Dec 08, 2023
https://doi.org/10.7773/cm.y2023.3362
Artículos
Estado de explotación y perspectivas de manejo pesquero del marlin rayado (Kajikia audax) en el océano Pacífico mexicano
1
 http://orcid.org/0009-0008-1410-0586
http://orcid.org/0009-0008-1410-0586
2
*
http://orcid.org/0000-0001-6705-1636
3
http://orcid.org/0000-0002-9846-0915
4
http://orcid.org/0000-0003-2753-6561
1Instituto Nacional de Pesca y Acuacultura, Centro Regional de Investigación Acuícola y Pesquera, 23020 La Paz, Baja California Sur, Mexico.
2Centro de Investigaciones Biológicas del Noroeste, 23096 La Paz, Baja California Sur, Mexico.
3Instituto Nacional de Pesca y Acuacultura, Centro Regional de Investigación Acuícola y Pesquera, 82010 Mazatlán, Sinaloa, Mexico.
4Fideicomiso de Investigación para el desarrollo del Programa Nacional de Aprovechamiento del Atún y Protección de Delfines y otros en torno a especies Acuáticas Protegidas (FIDEMAR), Centro de Investigación Científica y de Educación Superior de Ensenada, 22860 Ensenada, Baja California, Mexico.
5Instituto Nacional de Pesca y Acuacultura, Centro Regional de Investigación Acuícola y Pesquera, 28200 Colima, Colima, Mexico.
Se evaluó el estado de explotación del marlin rayado en el Pacífico mexicano con datos históricos (1963-2014) de captura y esfuerzo de flotas comerciales y deportivas, incluso datos de captura incidental. Se estimaron 8 indicadores de abundancia relativa independientes para ajustar un modelo dinámico de biomasa por máxima verosimilitud. Para el periodo más reciente (2015-2019), se evaluó el estado de explotación a través de simulaciones hechas con el modelo ajustado, y se tomó como referencia los datos disponibles de captura por unidad de esfuerzo de las flotas deportivas. Los resultados indicaron que hasta 2014, la abundancia del marlin rayado ha pasado por 3 etapas diferentes. La primera fue una tendencia decreciente de 1964 a 1977, donde las capturas y el esfuerzo de las flotas comerciales excedieron, respectivamente, el máximo rendimiento sostenible (MRS) y el esfuerzo para alcanzar el MRS. La segunda ocurrió de 1977 a 1990, y fue una tendencia estable de baja abundancia, inferior a la abundancia cuando se alcanza el MRS; este cambio se relacionó con la reducción del esfuerzo de las flotas comerciales, aunque con capturas superiores al MRS. La tercera ocurrió de 1990 a 2014, y mostró una tendencia al incremento de la abundancia, debido al cese de operaciones de las flotas comerciales. Este incremento ha sido lento debido al desarrollo de la pesca deportiva y al incremento de las capturas incidentales. Después de 2014, el indicador para las flotas deportivas sugirió un cambio en la abundancia; de acuerdo con las simulaciones, la mortalidad por pesca fue insuficiente para explicar dicho cambio. Se plantea como hipótesis que la mortalidad natural del marlin rayado ha sido alta desde la década de 2010. Se discuten las condiciones que pudieron influir en esta situación y sus implicaciones en el manejo pesquero.
Palabras clave: marlin rayado; Kajikia audax; modelo dinámico de biomasa; múltiples indicadores
Stock assessment of the striped marlin from the Mexican Pacific coast was done with historical (1963-2014) catch and effort data provided by commercial and recreational fleets, including bycatch data. Eight fishery-independent indices of relative abundance were used to fit a biomass dynamic model by maximum likelihood. For the most recent period (2015-2019), stock status was evaluated through simulations with the fitted model and available catch-per-unit-effort data of the recreational fleets as reference framework. Results indicated that up until 2014, striped marlin abundance showed 3 different trends. The first one was a decreasing trend from 1964 to 1977, where the catch and effort of commercial fleets exceeded the maximum sustainable yield (MSY) and the level of effort to reach MSY, respectively. The second one occurred from 1977 to 1990, and it was a stable trend with low abundance, less than the abundance when MSY is reached; this change was related to the reduction of commercial effort, although catches greater than the MSY were observed. The third one occurred from 1990 to 2014, and it showed increasing abundance due to the cessation of commercial fleets. This increase, however, has become slower because of the development of the recreational fishery and the increased bycatch. After 2014, the indicator estimated with the data from the recreational fleets suggested a shift in abundance; according to simulations, the fishing mortality was insufficient to explain this change. It is hypothesized that the natural mortality of striped marlin has been high since the mid-2010s. Conditions that could influence this situation and their implications on fishery management are discussed.
Key words: striped marlin; Kajikia audax; dynamic biomass model; multiple indicators
INTRODUCCIÓN
Entre las especies de picudos (familia Istiophoridae), el marlin rayado, Kajikia audax (Philippi, 1887), tiene la más amplia distribución latitudinal (entre 45° N y 45° S) en las regiones subtropicales y templadas (de 20 a 25 °C) de los océanos Índico y Pacífico (Howard y Ueyanagi 1965, Squire y Suzuki 1990, Domeier et al. 2003, McDowell y Graves 2008). En el océano Pacífico, las áreas de distribución del marlin rayado se configuran en forma de herradura, con 2 regiones transoceánicas en cada hemisferio (norte y sur) unidas en su extremo oriental a lo largo de la costa del continente americano (Squire y Suzuki 1990). El marlin rayado es particularmente abundante frente a las costas de la península de Baja California y en la boca del golfo de California, zona relevante que se considera centro de concentración poblacional de marlin rayado, tanto para su explotación como para su manejo pesquero (Squire 1987). En esta región, esta especie ha sido explotada por más de 50 años, tanto por flotas comerciales como por flotas deportivas.
La pesca deportiva tuvo su origen hacia 1930, cuando pescadores deportivos de Estados Unidos viajaban en sus propias embarcaciones al sur de la península de Baja California, entre cabo San Lucas y La Paz. En la década de 1960, cabo San Lucas seguía reportándose como un lugar con buenos sitios de pesca, e inició el desarrollo de servicios hoteleros para pescadores deportivos (Talbot y Wares 1975). Actualmente, las principales flotas deportivas se concentran en la boca del golfo de California, cerca del centro de mayor disponibilidad de marlin rayado en el Pacífico oriental (Ortiz et al. 2003, Domeier 2006), por lo que dependen de la abundancia de esta especie en la región. A pesar de su larga historia, la pesca deportiva en México tuvo su mayor crecimiento hasta principios de la década de 1990, sin que, hasta la fecha, se haya evaluado el efecto de esta pesquería en la abundancia del marlin rayado.
El aprovechamiento comercial empezó a finales de la década de 1950, cuando flotas atuneras de Japón expandieron sus áreas de pesca hacia el Pacífico oriental. En 1963 alcanzaron la región frente a la península de Baja California, donde las capturas de marlin rayado eran grandes e igualaban a las de atún (Talbot y Wares 1975). En la década de 1970, con la declaración de la zona económica exclusiva (ZEE) de México, se dio la transición a flotas mexicanas centradas en la pesca de atún, tiburón y escama, donde el marlin rayado formaba parte importante de las capturas incidentales. En 1984, se emitieron los primeros permisos de pesca comercial específicos para diversas especies de picudos, de las cuales el marlin rayado constituía la mayor parte de estas capturas. En 1991, se dejaron de otorgar permisos para la pesca de marlin rayado, y las flotas se orientaron a la pesca de pez espada. Eventualmente, gran parte de estas embarcaciones se dirigieron a la pesca de tiburón, por lo que, desde la década de 1990, estas flotas capturan marlin rayado de manera incidental. El estatus del marlin rayado en el Pacífico mexicano fue analizado por Klett-Traulsen y Aguilar-Ibarra (2001) con base en la serie histórica de capturas comerciales disponible en ese momento, es decir, de 1963 a 1990, que representa un periodo de intensa pesca por flotas palangreras tanto japonesas como mexicanas. Klett-Traulsen y Aguilar-Ibarra (2001) no incluyeron en su modelo la pesca deportiva, y para la época que abarcó el estudio, las capturas incidentales se consideraban insignificantes. Otras evaluaciones de marlin rayado que tienen una mayor cobertura geográfica en el océano Pacífico oriental se encuentran en los trabajos de Hinton y Bayliff (2002), Hinton y Maunder (2004), Hinton (2009) y Hinton y Maunder (2009) (Fig. 1a).

Figura 1 Área de estudio. (a) Área de estudio (área sombreada) con relación a las áreas de evaluación que se manejan en otras evaluaciones de marlin rayado (A1-A3) (e.g., Hinton y Maunder 2009). (b) Área de influencia (área sombreada) de las flotas comerciales y atuneras. (c) Área de influencia (área punteada) de las flotas deportivas. (d) Área de influencia (área con patrón de diamantes) de las flotas espadera y tiburoneras. I: Ensenada, Baja California; II: La Paz, Baja California Sur; III: Guaymas, Sonora; IV: Mazatlán, Sinaloa; V: Manzanillo, Colima; y VI: Acapulco, Guerrero.
Actualmente, el manejo de marlin rayado en México se centra en la pesca deportiva y en las capturas incidentales que se generan en las pesquerías de pez espada y tiburón. Los lineamientos de manejo se encuentran en distintos documentos legales (DOF 2007, 2012, 2013) y varios acuerdos vigentes que datan desde la década de 1980 (DOF 1987, 1994, 2008). Dada la importancia de la especie, en este trabajo se presenta una evaluación del estado de explotación del marlin rayado que se distribuye frente a las costas del Pacífico mexicano con una serie histórica de más de 50 años, en la que se incluyen capturas comerciales, deportivas e incidentales.
MATERIALES Y MÉTODOS
Captura y esfuerzo
Los datos de captura y esfuerzo usados en este estudio provinieron de flotas comerciales y deportivas que han operado en la ZEE del Pacífico mexicano entre 1963 y 2014 (Fig. 1b-d). Todas las capturas se expresaron en número de individuos, mientras que el esfuerzo se cuantificó en distintas unidades desglosadas en la Tabla 1. La información de la pesca comercial dirigida al marlin rayado provino de flotas palangreras tanto japonesas como mexicanas (1963-1990), las cuales se consideraron como una sola flota comercial (f 1) con un indicador de abundancia relativa, la cual ya estaba estandarizada (Klett-Traulsen y Aguilar-Ibarra 2001). La información de la pesca deportiva provino de 2 fuentes que abarcaron distintos periodos. La primera fuente se tomó del trabajo de Squire (1987), con información de 1969 a 1984, de la cual se seleccionaron datos de Baja California; Guaymas, bahía Kino y Puerto Peñasco, Sonora; Mazatlán, Sinaloa; y Acapulco, Zihuatanejo e Ixtapa, Guerrero; y estos datos se manejaron como provenientes de una sola flota (f 2). La segunda fuente abarcó información de 1985 a 2014, la cual provino de muestreos mensuales y reportes diarios de captura proporcionados por las flotas deportivas de 3 puertos clave en la boca del golfo de California: cabo San Lucas, Buenavista y Mazatlán. Esta información provino del Programa de Monitoreo de Pesca Deportiva del Instituto Nacional de Pesca y Acuacultura (PMPD-INAPESCA), y se supuso como información de otra flota (f 3). Las capturas de estas flotas se refieren a la captura retenida y desembarcada más el 26% de los individuos reportados como liberados, bajo el supuesto de que no lograron sobrevivir (Domeier et al. 2003), por lo que se incluyeron como parte de la mortalidad causada por esta flota. También se incluyeron otras flotas comerciales que capturan incidentalmente marlin rayado: una flota atunera con red de cerco (f4) (1993-2014) cuya información provino del Programa Nacional de Aprovechamiento del Atún y Protección del Delfín que opera el Fideicomiso de Investigación para el Desarrollo del Programa Nacional de Aprovechamiento del Atún y Protección de Delfines y otros en torno a Especies Acuáticas Protegidas (PNAAPD-FIDEMAR); una flota espadera de palangre (f 5) (1983-2002); y 3 flotas tiburoneras de palangre con diferentes puertos base (Manzanillo [f 6, 2003-2014], Ensenada [f 7, 2006-2014] y Mazatlán [f 8, 2006-2014]). La información de las flotas f 5 y f 6 provino del Programa de Pelágicos Mayores del Pacífico Central Mexicano del INAPESCA (PPMPC-INAPESCA), mientras que la información de las flotas f 7 y f 8 provino del Programa de Observadores a Bordo de Embarcaciones Mayores en la Pesca de Tiburón del Pacífico Mexicano operado por FIDEMAR (POBPT-FIDEMAR). Dado que para la mayor parte de las flotas no se contaba con datos estandarizados de captura y esfuerzo (Tabla 1), se utilizaron indicadores independientes de abundancia relativa en términos de captura por unidad de esfuerzo (CPUE), bajo el supuesto de que cada flota era lo suficientemente homogénea con una constante de capturabilidad específica (q), lo que permitió realizar las estimaciones que se describen en la siguiente subsección.
Tabla 1 Flotas que capturan marlin rayado (f 1-f 8) en la zona económica exclusiva del Pacífico mexicano y que fueron incluidas en el análisis de este trabajo. PMPD-INAPESCA, Programa de Monitoreo de Pesca Deportiva del Instituto Nacional de Pesca y Acuacultura; PNAAPD-FIDEMAR, Programa Nacional de Aprovechamiento del Atún y Protección del Delfín que opera el Fideicomiso de Investigación para el Desarrollo del Programa Nacional de Aprovechamiento del Atún y Protección de Delfines y otros en torno a Especies Acuáticas Protegidas; PPMPC-INAPESCA, Programa de Pelágicos Mayores del Pacífico Central Mexicano del INAPESCA; POBPT-FIDEMAR, Programa de Observadores a Bordo de Embarcaciones Mayores en la Pesca de Tiburón del Pacífico Mexicano operado por FIDEMAR.
| Fleet | Period | Type of catch | Fishing gear | Effort unit | Standarized data | Source1 |
| f 1 Commercial (Japan-Mexico) | 1963-1990 | Objective | Longline | Hooks × 103 | Yes | Klett and Aguilar (2001) |
| f 2 Sportfishing | 1969-1984 | Objective | Hook and line | Angler-days | No | Squire (1987) |
| f 3 Sportfishing | 1985-2014 | Objective | Hook and line | Trips | No | PMPD-INAPESCA |
| f 4 Tuna | 1993-2013 | Incidental | Purse seine | Sets | No | PNAAPD-FIDEMAR |
| f 5 Deep-sea billfish | 1983-2002 | Incidental | Longline | Hooks × 103 | No | PPMPC-INAPESCA |
| f 6 Billfish-shark | 2003-2014 | Incidental | Longline | Hooks × 103 | No | PPMPC-INAPESCA |
| f 7 Shark (Ensenada) | 2006-2014 | Incidental | Longline | Trips | No | POBPT-FIDEMAR |
| f 8 Shark (Mazatlán) | 2006-2014 | Incidental | Longline | Trips | No | POBPT-FIDEMAR |
1 See Materials and methods section for more on the source.
Análisis cuantitativo
Los datos de captura (comercial, deportiva e incidental) fueron incorporados a un modelo dinámico de abundancia (Haddon 2011):
donde N t es la abundancia del marlin rayado en el año t, N t - 1 es la abundancia en el año anterior (t - 1), ΣC t-1,f es la suma de las capturas de las distintas flotas (f 1, f 2, … f 8) en el año t - 1, r es la tasa máxima de crecimiento poblacional cuando N → 0, y K es la capacidad de carga en la zona de estudio.
Con los valores de N t obtenidos de la ecuación 1, se estimó la CPUE anual de cada flota a partir de:
donde Î t,f es la CPUE estimada de la flota f en el año t, q f es la capturabilidad específica de la flota f, y e ε es el error de observación con ε ~ N(0,σ 2). Se probaron distintos valores de K y r en la ecuación 1 para aproximar los valores de CPUE estimados (Î t,f , ecuación 2) a los valores de CPUE observados de cada flota (I t,f = C t,f /E t,f , donde C t,f y E t,f son la captura y el esfuerzo registrados de la flota f en el año t). Para tal propósito, se utilizó la función de verosimilitud (Haddon 2011):
donde L
f
(I
t,f
│r,K,q
f
) es la verosimilitud de I
t,f
dados los parámetros r, K y q
f
;
donde L T {I t,1 , I t,2 ,…I t,7 , I t,8 } es la verosimilitud conjunta de los 8 indicadores independientes de abundancia ponderados por el factor Ω f , que expresa el peso del indicador de cada flota y que se estimó como Ω f = λ f 2/Σ[λ f 2], donde λ 2 es el valor de varianza explicada de cada indicador (Deriso et al. 1985, Methot 1989, Punt y Hilborn 1996). El procedimiento de maximización se realizó mediante la rutina Solver de Excel con el método de resolución gradiente reducido generalizado no lineal. Con el fin de evitar la posible convergencia en un máximo local, se probaron distintos valores semilla de r y K, lo que permitió tener un intervalo amplio de valores de verosimilitud y construir la superficie de respuesta que garantizara la obtención de la máxima verosimilitud. Los intervalos de confianza de K, de r y de cada q f se estimaron con base en los perfiles de verosimilitud suponiendo que L(θ)max/L(θ) ~ χ2 y seleccionando aquellos valores de r, K y q f que cumplieran la condición 2 × [L(θ)max - L(θ)] ≤ χ2 2,α, para la cual se consideraron parámetros covariantes con valor crítico de χ2 2,0.05 = 5.99 (Venzon y Moolgavkar 1988, Haddon 2011). Finalmente, se estimaron los principales puntos de referencia: máximo rendimiento sostenible (MRS = rK/4), la abundancia cuando se alcanza el MRS (N MRS = K/2) y el esfuerzo para alcanzar el MRS (E MRS = r/2q f ) (Haddon 2011).
Simulaciones
Después de 2014, los únicos datos disponibles eran aquellos de las flotas deportivas (f 3). Suponiendo que la CPUE de f 3 continuaba siendo un indicador de abundancia, se realizaron simulaciones con el modelo de abundancia (ecuación 1) bajo 2 escenarios diferentes para evaluar el nivel de mortalidad que debió ocurrir para ajustar Î t,3 a los datos observados en los años más recientes (2015-2019). Para el escenario 1, se planteó la hipótesis de que solo con agregar una mortalidad equivalente a las capturas incidentales esperadas se logra el ajuste. Las capturas incidentales esperadas se estimaron como la proporción promedio de las capturas incidentales con respecto a la pesca deportiva durante el periodo 2010-2014. Para el escenario 2, se planteó la hipótesis de que para lograr el ajuste, es necesario agregar una mayor mortalidad que la esperada con solo las capturas incidentales. En este caso, la mortalidad se incrementó hasta alcanzar la máxima verosimilitud (ecuación 3) en el ajuste de Î t,3 a los datos observados. Los valores de K, r y q f se mantuvieron fijos según el ajuste original, por lo cual la verosimilitud se maximizó asignando distintos valores de captura total (ΣC t-1,f ).
RESULTADOS
Captura y esfuerzo
La pesca comercial de marlin rayado en el Pacífico mexicano se desarrolló rápidamente, y entre 1964 y 1971, se registraron las mayores capturas promedio, con 117,000 ind·año-1, y la máxima captura histórica de 212,000 individuos en 1968 (Fig. 2a). Entre 1972 y 1978, se observó una tendencia negativa en las capturas, con un promedio de 50,000 ind·año-1. Entre 1979 y 1990, se registró la mayor variabilidad interanual con el menor promedio de capturas (37,000 ind·año-1). Después de 1990, las capturas solo provinieron de la pesca deportiva (67.5%) o de la captura incidental (32.5%).
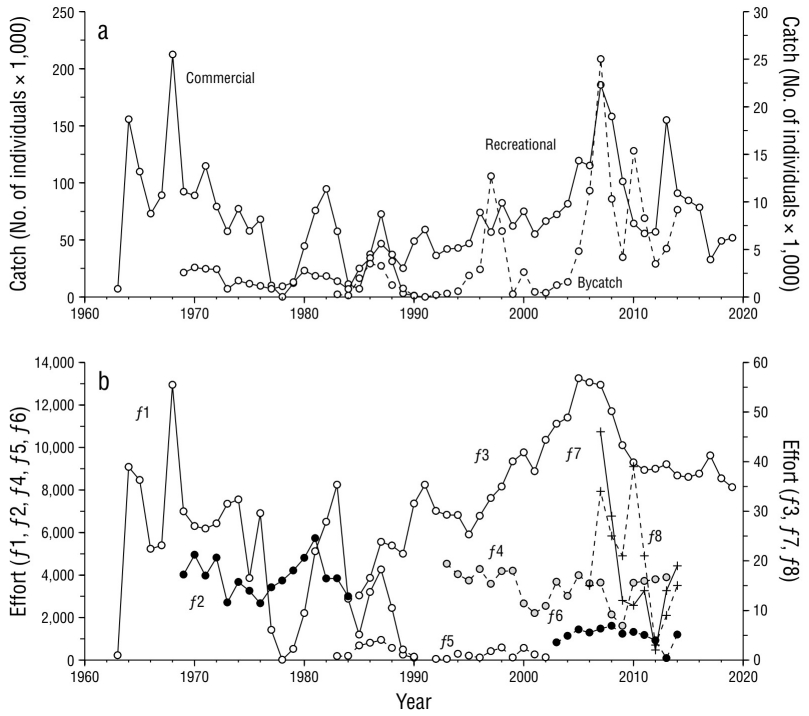
Figura 2 Captura y esfuerzo en la pesca de marlin rayado en el Pacífico mexicano durante el periodo de 1963 a 2014. (a) Capturas comerciales (eje izquierdo) y capturas deportivas e incidentales (eje derecho). (b) Esfuerzo por flota (f 1-f 8). Solo para la flota deportiva f 3, se tienen registros hasta 2019.
Las capturas de la pesca deportiva fueron menores que 2,000 ind·año-1, en promedio, hasta 1985. Después, inició un periodo de capturas ascendentes hasta 2007, cuando se alcanzó la máxima captura de 22,000 individuos. Posteriormente, las capturas de la pesca deportiva mostraron una amplia variación, desde 6,600 individuos en 2011 hasta 18,600 individuos en 2013. Después de 2013, las capturas disminuyeron hasta alcanzar un promedio de 5,300 ind·año-1 entre 2017 y 2019, lo que delinea una tendencia descendente que parece iniciarse unos años antes de 2010. Las capturas incidentales, por su parte, presentaron amplias oscilaciones. Las primeras ocurrieron en el periodo 1986-1987 y en 1997, con picos de captura de 3,200 y 12,600 individuos, respectivamente. Igual que con la pesca deportiva, en 2007 se dio la máxima captura con 25,000 individuos. Posteriormente, se registraron oscilaciones, con picos de captura incidental de 15,300 y 9,000 individuos en 2010 y 2014, respectivamente, lo que delinea una tendencia a disminuir.
El esfuerzo aplicado a la pesca de marlin rayado presentó un comportamiento similar al que mostraron las capturas (Fig. 2b). El esfuerzo de las flotas comerciales (f 1) se desarrolló rápidamente hasta alcanzar, en 1968, el máximo esfuerzo de 12.9 × 106 anzuelos. En los siguientes 10 años, la tendencia fue a disminuir el esfuerzo hasta casi desaparecer en 1979. En la década de 1980, se retomó la pesquería hasta alcanzar 8.2 × 106 anzuelos en 1983 y luego concluir con 109,000 anzuelos en 1990, último año en que se otorgaron permisos de pesca comercial para marlin rayado. A pesar de que el esfuerzo de la pesca deportiva provino de diferentes periodos de tiempo y distintas unidades de esfuerzo, se observó un patrón acorde con el desarrollo de las capturas de pesca deportiva. Entre 1969 y 1984, el esfuerzo de f 2 osciló de 2,600 a 5,700 días-pescador sin una tendencia aparente. A partir de 1985, el esfuerzo de f 3 inició con 12,900 viajes y, progresivamente, fue incrementando hasta 2005, cuando se alcanzó el mayor esfuerzo con 56,700 viajes. En los siguientes 6 años, el esfuerzo presentó una tendencia descendente, y a partir de 2012, el esfuerzo se estabilizó en un promedio de 37,800 viajes·año-1. El esfuerzo relacionado a las capturas incidentales de f4 a f 8 provino de distintas flotas que pescaban bajo distintas circunstancias (especies objetivo, distancia desde la costa, artes de pesca, entre otras), por lo que no fue posible hacer una comparación directa entre flotas y tampoco se pudo identificar una tendencia en su comportamiento.
Análisis cuantitativo
A pesar de la complejidad que significa trabajar con múltiples indicadores de abundancia (Î t,f ), el ajuste del modelo logró convergir en valores de r, K y q f cuyas verosimilitudes permitieron estimar sus intervalos de confianza (Fig. 3) y los correspondientes puntos de referencia (Tabla 2). Dada la variabilidad de las CPUE observadas, el ajuste mostró significación (α < 0.05) con la flota comercial f1 y con las flotas deportivas f 2 y f 3, cuyas r 2 oscilaron entre 0.33 y 0.44 (Tabla 3), y fue posible identificar algunos patrones similares en los indicadores de estas flotas (Fig. 4). Las flotas f 1 y f 2 mostraron una tendencia decreciente desde el inicio de sus registros hasta 1977-1978. En los siguientes 6 o 7 años, los indicadores de ambas flotas mostraron una oscilación con su punto máximo entre 1980 y 1982. El indicador de f 1 mostró una segunda oscilación con pico en 1987 y cese de operaciones en 1990, lo que coincide con una oscilación del indicador de f 3, entre 1985 y 1989 (Fig. 4). La abundancia reconstruida a partir de estos indicadores (Fig. 5) sugiere que, desde el inicio de la pesca comercial, la población de marlin rayado disminuyó continuamente hasta 1977, a una tasa promedio de 6.7% anual. En este periodo, tanto las capturas como el esfuerzo de las flotas comerciales (f 1) duplicaron prácticamente el MRS y el E MRS. Entre 1978 y 1990, según las estimaciones, la abundancia alcanzó su nivel más bajo, menor que la N MRS (40% de K). A partir de 1990, la abundancia mostró una tendencia a incrementarse debido al cese de las operaciones comerciales (f 1), y las flotas deportivas presentaron esfuerzo y capturas menores que el E MRS y el MRS. Sin embargo, el incremento no fue constante: entre 1990 y 1997, la tasa de incremento anual estimada fue de 5.3%, pero entre 1998 y 2007 disminuyó a 2.9%, y de 2008 a 2014, a 1.0%. En parte, esta reducción está asociada al desarrollo de la pesca deportiva con el cual alcanzó el máximo esfuerzo cercano al 66% del E MRS y a que las capturas totales, incluidas las incidentales, superaron el MRS con 47,000 individuos en 2007. Sin embargo, después de 2007, el esfuerzo de las flotas deportivas (f 3) disminuyó al 49% del E MRS, y las capturas totales oscilaron alrededor del 50% del MRS, por lo que, aparentemente, la pesca no es el único factor que ha influido en la reducción de la tendencia de incremento poblacional después de 2007.
Tabla 2 Parámetros del modelo dinámico de abundancia ajustados a los datos observados y puntos de referencia estimados a partir del modelo, así como los límites de sus intervalos de confianza con α = 0.05. Capturabilidad específica de cada flota, qf 1-qf 8; máximo rendimiento sostenible, MSY (por sus siglas en inglés); abundancia cuando se alcanza el MSY, N MSY; esfuerzo para alcanzar el MSY, E MSY; y flotas que capturan marlin rayado, f 1-f 8.
| Confidence interval | ||||
| Parameter | Mean | Lower | Upper | Units |
| K | 1,423 × 103 | 1,081 × 103 | 1,750 × 103 | Individuals |
| r | 0.1125 | 0.0659 | 0.2237 | y-1 |
| qf 1 | 1.660 × 10-5 | 1.082 × 10-5 | 2.576 × 10-5 | (Hooks × 1,000)-1 |
| qf 2 | 7.037 × 10-7 | 5.026 × 10-7 | 9.748 × 10-7 | Angler-days |
| qf 3 | 6.550 × 10-7 | 4.783 × 10-7 | 9.113 × 10-7 | Trip-1 |
| qf 4 | 2.631 × 10-8 | 0.642 × 10-8 | 10.701 × 10-8 | (Set × 1,000)-1 |
| qf 5 | 5.702 × 10-6 | 0.403 × 10-6 | 66.937 × 10-5 | (Hooks × 1,000)-1 |
| qf 6 | 2.301 × 10-6 | No convergence | No convergence | (Hooks × 1,000)-1 |
| qf 7 | 5.469 × 10-6 | 0.366 × 10-6 | 191.337 × 10-4 | Trip-1 |
| qf 8 | 2.240 × 10-5 | No convergence | No convergence | Trip-1 |
| Confidence interval | ||||
| Reference point | Mean | Lower | Upper | Units |
| MSY | 40,014 | 28,832 | 60,453 | Ind × y-1 |
| NMSY | 711,648 | 540,448 | 874,787 | Individuals |
| EMSY - f 1 | 3,388 | 3,045 | 4,343 | Hooks × 1,000 |
| EMSY - f 2 | 79,896 | 65,578 | 114,754 | Angler-days |
| EMSY - f 3 | 85,848 | 68,913 | 122,747 | Trips |
| EMSY - f 4 | 2,137 | 1,045 | 5,133 | Sets × 1,000 |
| EMSY - f 5 | 9,860 | 167 | 81,684 | Hooks × 1,000 |
| EMSY - f 6 | 24,440 | Undefined | Undefined | Hooks × 1,000 |
| EMSY - f 7 | 10,280 | 6 | 90,159 | Trips |
| EMSY - f 8 | 2,510 | Undefined | Undefined | Trips |
Tabla 3 Resultados de los análisis de varianza aplicados para evaluar la bondad de ajuste del modelo dinámico de abundancia en cada serie de datos por flota (f 1, f 2,…, f 8).
| Sum of squares | d.f. | Mean square | |||||||||||
| Fleet | Model | Residual | Model | Residual | Model | Residual | F | a | r2 | ||||
| f 1 | 653.7300 | 822.4100 | 1.0000 | 26 | 653.7300 | 31.6300 | 20.6675 | 0.0001 | ** | 0.4429 | |||
| f 2 | 0.0885 | 0.1687 | 1.0000 | 14 | 0.0885 | 0.0120 | 7.3448 | 0.0169 | * | 0.3441 | |||
| f 3 | 0.4878 | 0.9849 | 1.0000 | 28 | 0.4878 | 0.0352 | 13.8672 | 0.0009 | ** | 0.3312 | |||
| f 4 | 0.0003 | 0.0025 | 1.0000 | 19 | 0.0003 | 0.0001 | 2.3417 | 0.1424 | NS | 0.1097 | |||
| f 5 | 139.2900 | 1,237.9800 | 1.0000 | 17 | 139.2900 | 72.8200 | 1.9128 | 0.1846 | NS | 0.1011 | |||
| f 6 | 0.4807 | 9.7066 | 1.0000 | 10 | 0.4807 | 0.9707 | 0.4952 | 0.4977 | NS | 0.0472 | |||
| f 7 | 159.7800 | 635.7700 | 1.0000 | 6 | 159.7800 | 105.9600 | 1.5079 | 0.2654 | NS | 0.2008 | |||
| f 8 | 165.4300 | 1,001.6300 | 1.0000 | 7 | 165.4300 | 143.0900 | 1.1561 | 0.3179 | NS | 0.1417 | |||
*Significant with α < 0.05;
**highly significant with α < 0.01; NS: not significant.
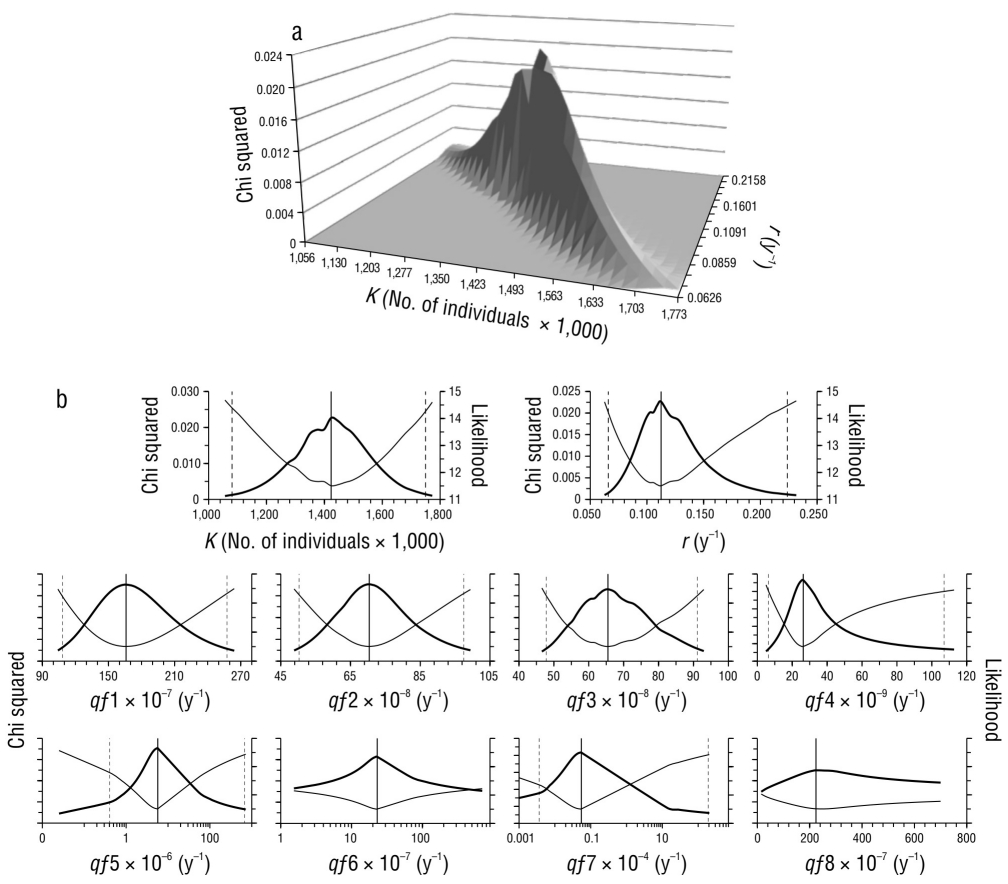
Figura 3 Bondad de ajuste al modelo dinámico de abundancia. (a) Superficie de respuesta de la verosimilitud frente a combinaciones de capacidad de carga (K) y tasa intrínseca de crecimiento (r). (b) Perfil de verosimilitud (línea delgada) y chi cuadrada (línea gruesa) de K, r y capturabilidad específica de cada flota (qf1-qf8). Las líneas verticales representan la máxima verosimilitud (línea continua) y los límites de su intervalo de confianza (α = 0.05, líneas discontinuas).
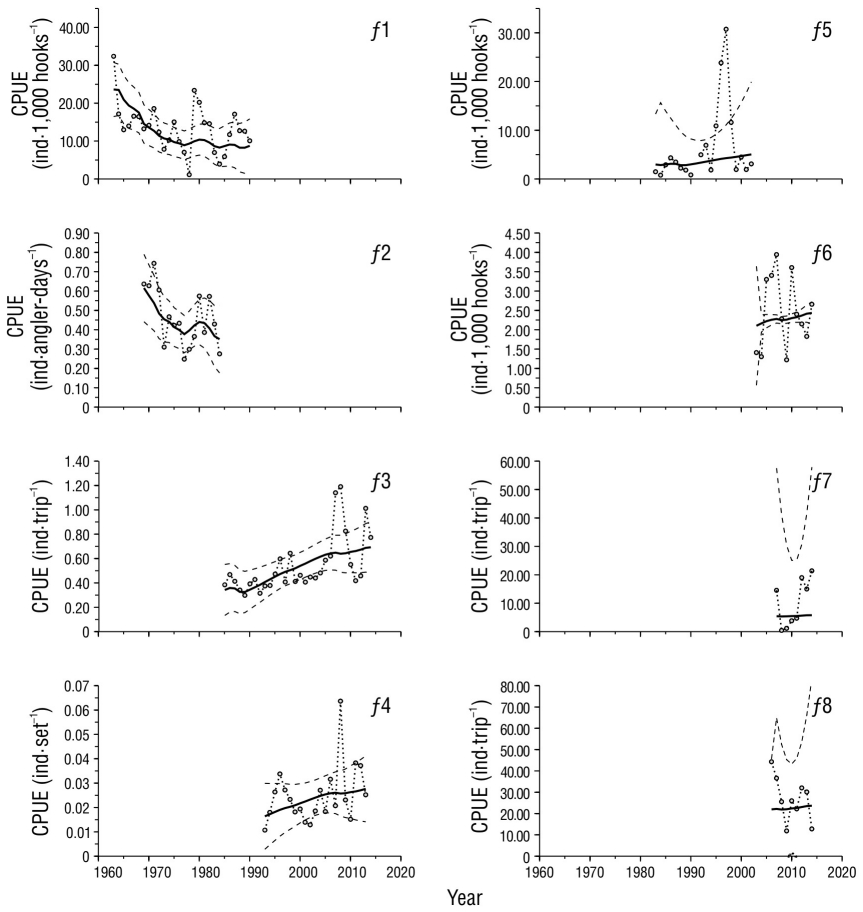
Figura 4 Captura por unidad de esfuerzo (CPUE) por flota (f 1-f 8). Se presentan los valores observados (línea punteada con círculos) y los valores estimados (línea gruesa continua) con sus intervalos de confianza (α = 0.05: líneas discontinuas). Los intervalos inferiores de las flotas f 5, f 7 y f 8 solo contienen valores negativos y no se muestran en las gráficas.

Figura 5 Salidas del modelo poblacional para el marlin rayado. (a) Reconstrucción de la abundancia por año. (b) Capturas totales registradas por año. Las líneas horizontales en (a) representan la abundancia cuando se alcanza el máximo rendimiento sostenible, y aquellas en (b), el máximo rendimiento sostenible. Se presentan los límites de confianza (α = 0.05) con líneas discontinuas.
Simulaciones
Después de 2014, las flotas deportivas (f 3) mostraron las CPUE más bajas de la serie histórica y acentuaron una tendencia negativa que inició con las oscilaciones registradas a partir de 2007 (Fig. 6a). Entre 2010 y 2014, las capturas incidentales representaron el 81.5% de las capturas deportivas, proporción que se tomó para estimar las capturas esperadas entre 2015 y 2019 y correr el primer escenario de simulación. Bajo este escenario, la CPUE estimada continuaría con su tendencia ascendente, lo cual difiere notablemente con respecto a la CPUE observada (Fig. 6a). El segundo escenario mostró que el mejor ajuste de la CPUE se logró cuando se agregó una captura 27 veces mayor que las capturas de la pesca deportiva, equivalente a 189 × 103 ind·año-1, en promedio (Fig. 6b). Este escenario implicó que la abundancia se habría reducido por debajo de la N MRS, similar a la abundancia que se alcanzó con la explotación de las flotas comerciales (f1) entre 1963 y 1990 (Fig. 6c).
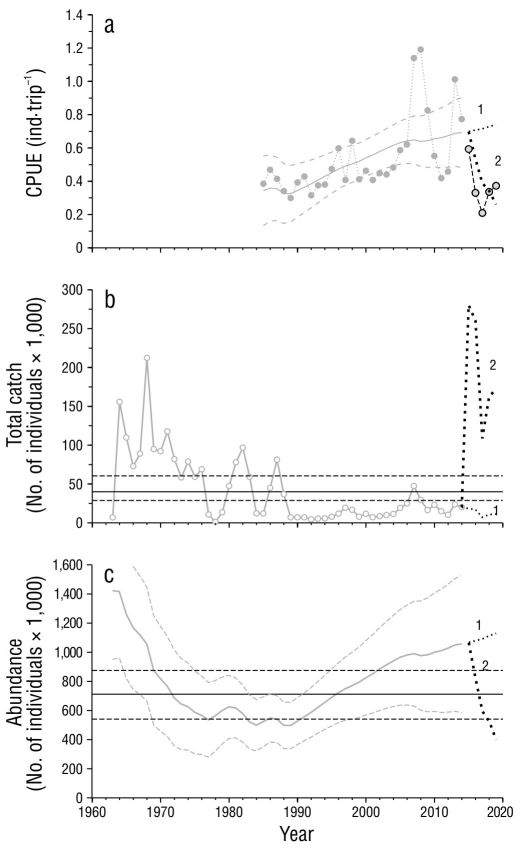
Figura 6 Simulaciones de los escenarios 1 y 2 definidos en la sección de Materiales y métodos. (a) Captura por unidad de esfuerzo (CPUE) observada de las flotas deportivas (f 3) con énfasis en el periodo 2015-2019 (línea discontinua delgada con círculos) y curvas ajustadas (líneas punteadas mediana y gruesa) según el escenario corrido. (b) Capturas totales requeridas para correr los escenarios de simulación (líneas punteadas mediana y gruesa). (c) Abundancia estimada según el escenario corrido. Las líneas horizontales en (b) representan el máximo rendimiento sostenible, y aquellas en (c), la abundancia cuando se alcanza el máximo rendimiento sostenible. Se presentan los límites de confianza (α = 0.05) con líneas discontinuas.
DISCUSIÓN
El modelo dinámico de abundancia en este estudio representó una alternativa, ya que la información de la pesquería de K. audax es limitada y dispersa, dada la amplia zona de distribución de la especie y de las áreas de operación de las flotas. Una ventaja de la información usada en este estudio es que provino de la región reconocida como centro poblacional de esta especie en el Pacífico oriental (Domeier 2006, McDowell y Graves 2008), por lo que se maximizan las probabilidades de registrar cambios significativos en la población. Un supuesto del modelo se refiere a que la abundancia, en número de individuos, resulta del balance entre reclutamiento e inmigración versus muertes y emigración. Generalmente, se supone que el efecto de la migración es despreciable, ya sea porque es baja la proporción de individuos que migran o porque la inmigración es igual a la emigración (Hilborn y Walters 1992, Quinn y Deriso 1999, Haddon 2011). En el caso del marlin rayado, hay evidencias basadas en ADN mitocondrial de que existen al menos 4 grupos poblacionales en el océano Pacífico (Australia, Ecuador, México y Pacífico Norte) entre los cuales hay poco intercambio genético (McDowell y Graves 2008). Esto concuerda con estudios de marcas satelitales que demuestran la fidelidad regional del marlin rayado (Domeier 2006) y con los sitios de desove, entre los que se encuentran las aguas cercanas a la península de Baja California y la boca del golfo de California (González-Armas et al. 2006), lo que enfatiza la escasa mezcla entre regiones. Se tienen evidencias de un grupo poblacional de marlin rayado en el Pacífico mexicano para el cual la migración no es un factor importante, por lo que se puede suponer que su abundancia se debe, principalmente, al balance entre reclutamiento y mortalidad. Dentro de la mortalidad, se considera la pesca y, con ella, otro supuesto clave de que la capturabilidad es constante. La capturabilidad es un parámetro ligado al comportamiento de las flotas de pesca y es una de las fuentes de variación más importantes al evaluar un stock (Hilborn y Walters 1992). Klett-Traulsen y Aguilar-Ibarra (2001) resaltaron la amplia variación del índice de abundancia (Î t ) de las flotas comerciales hacia la última década en que operaron (1980-1990), lo que coincide con la aplicación de cambios regulatorios que pudieron haber afectado su capturabilidad, como el establecimiento de la ZEE de México (Squire 1987) y 2 zonas de exclusión a la pesca comercial de picudos en la boca del golfo de California y el golfo de Tehuantepec (DOF 1987). Con estas medidas, hubo cambios en la composición de las flotas y en las zonas de pesca, por lo que no fue posible identificar si la variación en el indicador se debió a cambios en la capturabilidad o a cambios efectivos en la abundancia. La incorporación de las flotas deportivas f 2 y f 3 en este estudio muestra que sus indicadores (Î t,f ) tuvieron tendencias y oscilaciones similares a las de las flotas comerciales f 1 durante el periodo en que operaron simultáneamente. Squire (1987) también encontró correlación entre las CPUE de las flotas comerciales y las de las flotas deportivas entre 1969 y 1976, por lo que consideró que el indicador de las flotas deportivas podría representar la abundancia del marlin rayado, incluso en un área más grande que su área de influencia. Dada la consistencia en las oscilaciones y las tendencias de estas flotas, se puede afirmar que los indicadores independientes de cada flota (Î t,f ) parecen responder a una señal común. Considerando que las flotas deportivas operan en áreas más restringidas que las comerciales, las medidas regulatorias que pudieron afectar la capturabilidad de las flotas comerciales difícilmente pudieron afectar en la misma forma a las flotas deportivas. Por ello, es más probable que la señal de respuesta común esté asociada a cambios efectivos en la abundancia que a la capturabilidad.
Aunque el modelo explica solo una proporción de la variabilidad en los indicadores de abundancia, dicha proporción es estadísticamente significativa, al menos para las flotas comerciales y deportivas. Esto implica que el modelo tiene baja resolución para explicar los cambios de corto plazo, pero se supone que tiene la capacidad de registrar los cambios de tendencia en la abundancia de la población. En este sentido, se puede plantear que, entre 1963 y 2014, la abundancia de marlin rayado ha pasado por 3 etapas con diferentes tendencias: una tendencia decreciente de 1964 a 1977, asociada al esfuerzo de las flotas comerciales (f 1) que excedieron el E MRS y que produjeron capturas por encima del MRS durante todo ese periodo; una tendencia estable de 1977 a 1990 con baja abundancia, inferior a la N MRS, asociada a la reducción del esfuerzo de las flotas comerciales y de las capturas en general, aunque todavía con años en que se registraron capturas mayores que el MRS; y una tendencia al incremento después de 1990, motivado principalmente por el cese de operaciones de las flotas comerciales (f 1). Sin embargo, de acuerdo con el modelo ajustado, este incremento en la abundancia poblacional ha sido cada vez más lento y, en parte, está correlacionado con el desarrollo de la pesca deportiva entre 1995 y 2007, cuando el número de viajes y las capturas fueron cada vez mayores. Además, en el mismo periodo, las capturas incidentales mostraron una tendencia ascendente, lo cual parece haber contribuido también a que el crecimiento de la población se haya hecho más lento en los últimos años de registro (2008-2014). Estos resultados coinciden, en términos generales, con otras evaluaciones de marlin rayado que tienen una mayor cobertura geográfica (Fig. 1a), la cual abarca el océano Pacífico oriental (Hinton y Bayliff 2002, Hinton y Maunder 2004, Hinton 2009). Hinton y Maunder (2009) presentaron la evaluación más reciente, en la cual integraron información de una gran área que se extiende al este del meridiano 145° W, entre los paralelos 5° S y 45° N; estos autores emplearon el modelo estadístico Stock Synthesis (Methot 2009), y sus resultados confirmaron lo que se reportó en evaluaciones anteriores: la abundancia de marlin rayado en el océano Pacífico oriental se encuentra en o por encima del nivel esperado para obtener el MRS, bajo un esfuerzo de pesca menor que el que se esperaría para obtener el MRS y sin indicios de que el esfuerzo pueda incrementarse, por lo que se considera que el marlin rayado en el océano Pacífico oriental mantiene una condición poblacional biológicamente aceptable (Hinton y Bayliff 2002, Hinton y Maunder 2004, Hinton 2009).
De acuerdo con nuestro estudio, el estado de producción y abundancia parece ser cierto solo hasta 2014. Después de este año y hasta 2019, la CPUE de las flotas deportivas (f 3) mostró una tendencia decreciente, y las inferencias hechas a partir de estas flotas indicaron que la mortalidad requerida para explicar este cambio de tendencia equivale a 26 veces las capturas registradas en el mismo periodo, es decir, una mortalidad promedio de 196,000 ind·año-1. Esta cifra solo es comparable con el periodo de mayor explotación de marlin rayado (1963-1990), cuando se registraron las mayores capturas de la serie histórica. En ese periodo, el mayor promedio de capturas durante 5 años consecutivos sucedió entre 1964 y 1968, cuando se registraron capturas promedio de 128,000 ind·año-1. Es posible considerar que una mortalidad tan elevada como la estimada para los años más recientes sea evidencia de que se tiene un registro incompleto de las capturas. Este es un problema frecuente en el análisis de pesquerías debido, entre otras cosas, a problemas logísticos, inaccesibilidad a los sitios de desembarco o prácticas deficientes de manejo pesquero (Cisneros-Montemayor et al. 2013), y el caso de la pesca de marlín rayado no es la excepción (Hinton y Maunder 2009). Se trata de una fuente de incertidumbre importante en toda evaluación pesquera, que en el caso de México, se estima que llega a ser hasta el doble de los registros oficiales de desembarco (Cisneros-Montemayor et al. 2013). En este contexto, la mortalidad estimada a través de las simulaciones es notablemente mayor que el doble de los desembarcos registrados de marlin rayado, por lo que parece una mortalidad muy grande para atribuirla exclusivamente a capturas incidentales no actualizadas o, incluso, a capturas no reportadas, ilegales o no reguladas.
Para distintas especies de depredadores tope en el océano Pacífico, se ha encontrado que la variabilidad de la biomasa en los stocks no se puede atribuir completamente a la pesca (Sibert et al. 2006). Existe un caso particular con otra especie pelágica, el calamar gigante (Dosidicus gigas) en el golfo de California. Después de un periodo de alta abundancia (1996-2010) en el cual se capturaban alrededor de 75,000 t·año-1, en promedio, con picos que rebasaban las 100,000 t, se presentó un declive entre 2011 y 2014, con capturas promedio de 34,000 t·año-1, y desde 2015 se registran capturas promedio de 1,500 t·año-1 (SEMARNAP 1999; CONAPESCA 2010, 2018). Aunque no se puede descartar el efecto de la pesca, hay evidencias de que el decremento del calamar está también asociado a un hábitat pelágico cada vez más caliente, con menos clorofila a y surgencias considerablemente bajas en la región central del golfo de California cercana al puerto de Guaymas (Robinson et al. 2013). Este caso del calamar es importante para el análisis que se presenta en este trabajo por al menos 2 razones. La primera es que, así como el calamar gigante, el marlin rayado se encuentra entre las especies de mayor tasa de crecimiento corporal (Markaida et al. 2004, Kopf et al. 2011), por lo que la dinámica poblacional de estas especies depende, en gran medida, de las condiciones tróficas del ambiente que habitan y que además comparten (Abitia-Cárdnas et al. 2002, Bazzino et al. 2010, Robinson et al. 2013). La segunda razón tiene que ver con el hecho de que el propio calamar gigante es un componente importante en la dieta del marlin rayado (Abitia-Cárdenas et al. 1997, 2002, 2011). Aunque el análisis de Robinson et al. (2013) se centra en la región central del golfo de California, existen evidencias que sugieren que las condiciones que afectaron las surgencias no se restringieron solamente a la región de Guaymas, sino que pudo haber sido un proceso de mayor alcance geográfico, el cual abarcó incluso todo el golfo de California. De acuerdo con la serie histórica del índice de surgencias costeras (ISC) (Bakun 1973, 1975), estimado para el punto ubicado en la boca del golfo de California (21° N, 107° W) (Fig. 7)1, se observó un patrón de variación muy similar al que registraron Robinson et al. (2013) para la región central del golfo de California. Hasta antes de 2003, predominaron los valores positivos del ISC, a pesar de presentar valores con amplias oscilaciones. En contraste, entre octubre de 2003 y julio de 2004, el ISC presentó valores negativos, con promedio cercano a -300 m3·s-1 por 100 m de línea de costa, y posteriormente, los valores se mantuvieron negativos, con promedio cercano a los -30 m3·s-1 por 100 m de línea de costa hasta 2010. Sin duda, se requieren análisis más profundos para comprender los mecanismos a través de los cuales las surgencias pueden afectar a una especie en particular dentro de la trama trófica; sin embargo, la variación del ISC antes descrito coincide con el periodo en que el indicador de abundancia de marlin rayado presentó grandes oscilaciones (2007-2011) y parece marcar el cambio en la tendencia del mismo indicador (Fig. 6). La correspondencia temporal de estos resultados nos ofrece la base para hipotetizar que la población de marlin rayado podría estar bajo condiciones tróficas desfavorables que han favorecido una mayor mortalidad natural, lo cual podría ser un factor, adicional a las capturas, que ayuda a explicar la reciente tendencia negativa en la abundancia de marlin rayado.

Figura 7 Índice de surgencias costeras para la posición ubicada en la boca del golfo de California (21° N, 107° W). Datos estimados por la División de Investigación Ambiental (NOAA-SWFSC) y disponibles en: https://oceanview.pfeg.noaa.gov/products/upwelling/bakun.
En términos de manejo, esto implica que, aunque las distintas flotas analizadas operan por debajo de los niveles del MRS y del E MRS, la abundancia de marlin rayado podría estar por debajo de la N MRS. De acuerdo con los análisis, esta situación parece estar fuertemente influenciada por las limitantes condiciones tróficas a las que está sujeto el marlin rayado, por lo cual difícilmente se puede pensar en una medida de manejo que pueda incidir directamente. Sin embargo, se pueden tomar decisiones de manejo que permitan mitigar la presión ambiental sobre esta especie. Cualquier medida de manejo que ayude a reducir significativamente la mortalidad por pesca ayudaría a este propósito. Entre otras alternativas, el fortalecimiento de la práctica de captura-liberación en la pesca deportiva y la reducción de las cuotas de captura incidental que se encuentran actualmente vigentes en la legislación mexicana son medidas de manejo que podrían aplicarse de manera simultánea a las 2 principales fuentes de mortalidad en las que se puede incidir a través del manejo pesquero del marlin rayado.
AGRADECIMIENTOS
Los autores reconocen el apoyo que el Instituto Nacional de Pesca y Acuacultura (INAPESCA) ha otorgado tanto al Programa de Monitoreo de Pesca Deportiva (PMPD-INAPESCA) como al Programa de Pelágicos Mayores del Pacífico Central mexicano del INAPESCA (PPMPC-INAPESCA) desde hace más de 40 años y con los cuales se generaron parte de los datos que se analizaron para este estudio. Otras organizaciones también proporcionaron información relevante que fue analizada en este estudio: Programa Nacional de Aprovechamiento del Atún y Protección del Delfín que opera el Fideicomiso de Investigación para el Desarrollo del Programa Nacional de Aprovechamiento del Atún y Protección de Delfines, y otros en torno a Especies Acuáticas Protegidas (PNAAPD-FIDEMAR); Programa de Pelágicos Mayores del Pacífico Central mexicano del INAPESCA (PPMPC-INAPESCA); y Programa de Observadores a Bordo de Embarcaciones Mayores en la Pesca de Tiburón del Pacífico mexicano operado por FIDEMAR (POBPT-FIDEMAR). A todas estas organizaciones y sus representantes que nos facilitaron el acceso a la información, nuestro más amplio reconocimiento. EMB agradece el apoyo financiero recibido a través del proyecto Myers del Centro de Investigaciones Biológicas del Noroeste para la publicación de esta investigación.
REFERENCIAS
Abitia-Cárdenas, L.A, Galván-Magaña, F., Cruz-Escalona, V., Peterson, M.S, Rodríguez-Romero, J. 2011. Daily food intake of Kajikia audax (Philippi, 1887) off Cabo San Lucas, Gulf of California, Mexico. Lat Am J Aquat Res. 39(3):449-460. https://doi.org/10.3856/vol39-issue3-fulltext-6 [ Links ]
Abitia-Cárdenas, LA, Galván-Magaña, F, Rodríguez-Romero, J 1997. Food habits and energy values of prey of striped marlin, Tetrapturus audax, off the coast of Mexico. Fish Bull. 95:360-368. [ Links ]
Abitia-Cárdenas, LA, Muhlia-Melo, A, Cruz-Escalona, V, Galván-Magaña, F. 2002. Trophic dynamics and seasonal energetics of striped marlin Tetrapturus audax in the southern Gulf of California, Mexico. Fish Res. 57(3):287-295. http://doi.org/10.1016/S0165-7836(01)00350-2 [ Links ]
Bakun, A. 1973. Coastal upwelling indices, west coast of North America. US Department of Commerce. NOAA Tech Rep. Seattle (WA): NMFS SSRF-671. 112 p. [ Links ]
Bakun, A. 1975. Daily and weekly upwelling indices, west coast of North America, 1967-73. NOAA Tech Rep: NMFS SSRF-693. p. 16. [ Links ]
Bazzino, G, Gilly, WF, Markaida, U, Salinas-Zavala, CA, Ramos-Castillejo, J. 2010. Horizontal movements, vertical hábitat utilization and diet of the jumbo squid (Dosidicus gigas) in the Pacific Ocean off Baja California Sur, Mexico. Prog Oceanogr. 86(1-2):59-71. https://doi.org/10.1016/j.pocean.2010.04.017 [ Links ]
Cisneros-Montemayor, AM, Cisneros-Mata, MA, Harper, S, Pauly, D. 2013. Extent and implications of IUU catch in Mexico’s marine fisheries. Mar Pol. 39:283-288. https://doi.org/10.1016/j.marpol.2012.12.003 [ Links ]
[CONAPESCA] Comisión Nacional de Acuacultura y Pesca. 2010. Anuario estadístico de acuacultura y pesca 2008. Mazatlán (Sinaloa, Mexico): Secretaría de Agricultura, Ganadería, Desarrollo Rural, Pesca y Alimentación. 196 p. [ Links ]
[CONAPESCA] Comisión Nacional de Acuacultura y Pesca. 2018. Anuario estadístico de acuacultura y pesca 2018. Mazatlán (Sinaloa, Mexico): Secretaría de Agricultura y Desarrollo Rural. 293 p. [ Links ]
Deriso, RB, Quinn, TJ II, Neal, PR. 1985. Catch-age analysis with auxiliary information. Can J Fish Aquat Sci. 42:815-824. http://doi.org/10.1139/f85-104 [ Links ]
[DOF] Diario Oficial de la Federación. 1987 Aug 28. Acuerdo que regula el aprovechamiento de las especies marlin, pez vela y pez espada, conocidas comúnmente como picudos, en la zona económica exclusiva del litoral del Océano Pacífico. Mexico: Secretaría de Pesca. [ Links ]
[DOF] Diario Oficial de la Federación. 1994 Mar 16. Aviso por el que se da a conocer el establecimiento de épocas y zonas de veda para la pesca de diferentes especies de la fauna acuática en aguas de jurisdicción federal de los Estados Unidos Mexicanos. Mexico: Secretaría de Pesca. [ Links ]
[DOF] Diario Oficial de la Federación. 2007 Jul 24. Ley General de Pesca y Acuacultura Sustentables. Mexico: Secretaría de Gobernación. [ Links ]
[DOF] Diario Oficial de la Federación. 2008 Sep 12. Acuerdo mediante el cual se establece el volumen de captura incidental permitido en operaciones de pesca de tiburón y rayas en aguas de jurisdicción federal de los Estados Unidos Mexicanos ubicados en el Océano Pacífico. Mexico: Secretaría de Agricultura, Ganadería Desarrollo Rural, Pesca y Alimentación. [ Links ]
[DOF] Diario Oficial de la Federación. 2012 Aug 24. Acuerdo por el que se da a conocer la actualización de la Carta Nacional Pesquera. Mexico: Secretaría de Agricultura, Ganadería Desarrollo Rural, Pesca y Alimentación. [ Links ]
[DOF] Diario Oficial de la Federación. 2013 Nov 25. Modificación a la Norma Oficial Mexicana NOM-017-PESC-1994, para regular las actividades de pesca deportivo-recreativa en las aguas de jurisdicción federal de los Estados Unidos Mexicanos, publicada el 9 de mayo de 1995. Mexico: Secretaría de Agricultura, Ganadería Desarrollo Rural, Pesca y Alimentación. [ Links ]
Domeier, M.L. 2006. An analysis of Pacific striped marlin (Tetrapturus audax) horizontal movement patterns using pop-up satellite archival tags. Bull Mar Sci. 79(3):811-825. [ Links ]
Domeier, M.L., Dewar, H., Nasby-Lucas, N. 2003. Mortality rate of striped marlin (Tetrapturus audax) caught with recreational tackle. Mar Freshw Res. 54(4):435-445. https://doi.org/10.1071/MF01270 [ Links ]
González-Armas, R., Klett-Traulsen, A., Hernández-Herrera, A. 2006. Evidence of billfish reproduction in the southern Gulf of California, Mexico. Bull Mar Sci. 79(3):705-717. [ Links ]
Haddon, M. 2011. Modelling and quantitative methods in fisheries. 2nd ed. New York (NY): Chapman & Hall/CRC. 433 p. [ Links ]
Hilborn, R, Walters, CJ. 1992. Quantitative fisheries stock assessment: choice, dynamics and uncertainty. London (UK): Routledge, Chapman & Hall. 570 p. [ Links ]
Hinton, MG. 2009. Assessment of striped marlin in the Eastern Pacific Ocean in 2008 and Outlook for the future. Inter-American Tropical Tuna Commission. 10th Stock Assessment Review Meeting, 12-15 May 2009. Doc. SARM-10-08. La Jolla (CA). 18 p. [ Links ]
Hinton, MG, Bayliff, WH. 2002. Status of striped marlin in the eastern Pacific Ocean in 2001 and outlook for 2002. Stock Assess. Rep. 3. La Jolla (CA): Inter-Amer Trop Tuna Comm. p. 328-364. [ Links ]
Hinton, MG, Maunder, MN. 2004. Status of striped marlin in the eastern Pacific Ocean in 2002 and outlook for 2003-2004. In: Bayliff, WH, SAR, Inter-American Tropical Tuna Commission Rep 4. La Jolla (CA): Inter-American Tropical Tuna Commission p. 287-310. [ Links ]
Hinton, MG, Maunder, MN. 2009. Status and trends of striped marlin in the northeast Pacific Ocean in 2009. Stock Assess. Rep. 11: Inter-American Tropical Tuna Commission. p. 163-218. [ Links ]
Howard, JK, Ueyanagi, S. 1965. Distribution and relative abundance of billfishes (Istiophoridae) of the Pacific Ocean. Stud Trop Oceanogr. 2:1-134. [ Links ]
Klett-Traulsen, A, Aguilar-Ibarra, A. 2001. Marlin. In: Cisneros-Mata, MA, Beléndez-Moreno, LF, Zárate-Becerra, E, Gaspar-Dillanes, MT, López-González, LC, Saucedo-Ruiz, C, Tovar-Ávila, J. (eds.), Sustentabilidad y Pesca Responsable en México: Evaluación y Manejo 1999-2000. Mexico City (Mexico): Instituto Nacional de la Pesca. p. 437-466. [ Links ]
Kopf, RK, Davie, PS, Bromhead, D, Pepperell, JG. 2011. Age and growth of striped marlin (Kajikia audax) in the Southwest Pacific Ocean. ICES J Mar Sci. 68(9):1884-1895. https://doi.org/10.1093/icesjms/fsr110 [ Links ]
Markaida, U, Quiñonez-Velázquez, C, Sosa-Nishizaki, O. 2004. Age, growth and maturation of jumbo squid Dosidicus gigas (Cephalopoda:Ommastrephidae) from the Gulf of California, Mexico. Fish Res. 66(1):31-47. https://doi.org/10.1016/S0165-7836(03)00184-X [ Links ]
McDowell, JR, Graves, J.E. 2008. Population structure of striped marlin (Kajikia audax) in the Pacific Ocean based on analysis of microsatellite and mitocondrial DNA. Can J Fish Aquat Sci. 65:1307-1320. https://doi.org/10.1139/F08-054 [ Links ]
Methot, RD. 1989. Synthetic estimates of historical abundance and mortality for northern anchovy. Am Fish Soc Symp. 6:66-82. [ Links ]
Methot, R.D. 2009. Stock assessment: operational models in support of fisheries management. In: Beamish, RJ, Rothschild, BJ (eds.), Future of Fishery Science - Proceedings of the 50th Anniversary Symposium of the American Institute of Fishery Research Biologists. Fish & Fisheries Series, vol. 31. Seattle (WA): Springer. p. 137-165. [ Links ]
Ortiz, M, Prince, E.D., Serafy, JE, Holts, DB, Davy, KB, Pepperell, JG, Lowry, MB, Holdsworth, JC. 2003. Global overview of the major constituent-based billfish tagging programs and their results since 1954. Mar Freshwater Res. 54(4):489-507. https://doi.org/10.1071/MF02028 [ Links ]
Punt, AE, Hilborn, R. 1996. Biomass dynamic models. User’s manual. FAO Computerized Information Series. Fisheries. 10:62. [ Links ]
Quinn, TJ-II, Deriso, RB. 1999. Quantitative fish dynamics. Oxford: Oxford University Press. 542 p. [ Links ]
Robinson, CJ, Gómez-Gutiérrez, J, Salas de León, D.A. 2013. Jumbo squid (Dosidicus gigas) landings in the Gulf of California related to remotely sensed SST and concentrations of chloropyll a (1998-2012). Fish Res. 137:97-103. https://doi.org/10.1016/j.fishres.2012.09.006 [ Links ]
[SEMARNAP] Secretaría de Medio Ambiente Recursos Naturales y Pesca. 1999. Anuario estadístico de pesca 1998. Tlalpan (Mexico City): Comisión Nacional de Acuacultura y Pesca. 244 p. [ Links ]
Sibert, J., Hampton, J., Kleiber, P., Maunder, M. 2006. Biomass, size, and trophic status of top predators in the Pacific Ocean. Science. 314(5806):1773-1776. [ Links ]
Squire, JL. 1987. Pacific billfish angler catch rates for key area stock assessments. Mar Fish Rev. 49(2):15-25. [ Links ]
Squire, JL, Suzuki, Z. 1990. Migration trends of striped marlin (Tetmpturus audax) in the Pacific Ocean. In: Stroud, RH (ed.) Planning the future of billfishes: research and management in the 90s and beyond. Savannah (GA): National Coalition for Marine Conservation, Inc. p. 67-86. [ Links ]
Talbot, G.B., Wares, P.G. 1975. Fishery for Pacific billfish off southern California and Mexico, 1903-69. Trans Am Fish Soc. 104(1):1-12 https://doi.org/10.1577/1548-8659(1975)104<1:FFPBOS>2.0.CO;2 [ Links ]
Venzon, D.J., Moolgavkar, S.H. 1988. A method for computing profile-likelihood-based confidence intervals. J Royal App Statistic Soc. 37(1):87-94. https://doi.org/10.2307/2347496 [ Links ]
1 La División de Investigación Ambiental del Centro de Ciencia Pesquera del Suroeste de la Oficina Nacional de Administración Oceánica y Atmosférica (NOAA - Southwest Fisheries Science Center - Environmental Research Division) calcula este índice de surgencias costeras en una red de puntos geográficos, a lo largo de la costa del océano Pacífico oriental y los datos están disponibles en https://oceanview.pfeg.noaa.gov/products/upwelling/bakun.
Recibido: 16 de Mayo de 2022; Aprobado: 14 de Abril de 2023
 This is an open-access article distributed under the terms of the Creative Commons Attribution License
This is an open-access article distributed under the terms of the Creative Commons Attribution License
