










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkCiencias marinas
Print version ISSN 0185-3880
Cienc. mar vol.39 n.2 Ensenada Jun. 2013
https://doi.org/10.7773/cm.v39i2.2236
Artículos
Enriquecimiento invernal de nutrientes inorgánicos en el canal de Ballenas, golfo de California
Wintertime enrichment of inorganic nutrients in the Ballenas Channel, Gulf of California
Eunise Vanessa Torres-Delgado, Francisco Delgadillo-Hinojosa*, Víctor Froylán Camacho-Ibar, Miguel Angel Huerta-Díaz, José Antonio Segovia-Zavala, José Martín Hernández-Ayón, Salvador Galindo-Bect
Instituto de Investigaciones Oceanológicas, Universidad Autónoma de Baja California, Carretera Transpeninsular Tijuana-Ensenada No. 3917, Fraccionamiento Playitas, Ensenada 22860, Baja California, México.
* Corresponding author. E-mail: fdelgadillo@uabc.edu.mx
Received October 2012,
received in revised form February 2013,
accepted February 2013.
RESUMEN
Se realizaron tres cruceros oceanográficos en la región del canal de Ballenas (CB) durante el invierno de 2002, la primavera de 2003 y el verano de 2004, con el objetivo de estudiar el impacto de la mezcla vertical de la columna de agua (Φ ) sobre la concentración superficial de los nutrientes inorgánicos disueltos (NO3, PO4 y Si(OH)4). Las concentraciones promedio de nutrientes integrados en los primeros 100 m del CB mostraron una clara variación estacional, con valores altos en el invierno (18.2 ± 1.1 μM de NO3, 2.36 ± 0.08 μM de PO4 y 35.4 ± 3.2 μM de Si(OH)4), asociados a condiciones de mezcla intensa de la columna de agua (Φ = 0.7-16 J m-3), y valores bajos en el verano (7.1 + 1.0 μM de NO3 1.22 ± 0.06 μM de PO4 y 16.0 ± 2.1 μM de Si(OH)4), asociados a condiciones de estratificación de la columna de agua (Φ = 150-223 J m-3). En contraste, por debajo de los 200 m la variabilidad estacional de la concentración de los tres nutrientes fue muy pequeña. El análisis hidrográfico indicó que el enriquecimiento superficial de nutrientes en invierno se debió a la intensificación de la mezcla de la columna de agua, la cual transportó verticalmente el Agua Subsuperficial Subtropical (ASS, fría y rica en nutrientes) hacia la superficie del CB. En contraste, el empobrecimiento de nutrientes durante el verano se debió a que el consumo por la actividad biológica se combinó con la estratificación de la columna de agua, la cual redujo el flujo vertical del ASS y, en consecuencia, el aporte de nutrientes hacia la superficie de este sistema. Las razones N:P, Si:N y un exceso de PO4 > 0.65 μM en las aguas superficiales indicaron que el NO3 fue el nutriente potencialmente limitante para la producción primaria de esta provincia oceanográfica del golfo de California.
Palabras clave: nutrientes, enriquecimiento, mezcla vertical, canal de Ballenas.
ABSTRACT
Three oceanographic cruises were carried out in the region of the Ballenas Channel (BC) during winter 2002, spring 2003, and summer 2004, in order to study the impact of vertical mixing of the water column (Φ) on the surface concentration of dissolved inorganic nutrients (NO3, PO4, and Si(OH)4). Mean nutrient concentrations integrated over the upper 100 m of BC showed a clear seasonal variation, with higher values in winter (18.2 + 1.1 μM NO3, 2.36 + 0.08 μM PO4, and 35.4 + 3.2 μM Si(OH)4), associated with a well-mixed water column (Φ = 0.7-16 J m-3), and low values in summer (7.1 ± 1.0 μM NO3, 1.22 ± 0.06 μM PO4, and 16.0 ± 2.1 μM Si(OH)4), related to vertically stratified water column conditions (Φ = 150-223 J m-3). In contrast, below 200 m the seasonal variability of the three nutrient concentrations was reduced. The hydrographic analysis indicated that the wintertime surface nutrient enrichment was due to an intensification of the water column mixing, which vertically transported Subtropical Subsurface Water (SSW, cold and nutrient-enriched) to the surface of BC. In contrast, nutrient impoverishment in summertime was due to biological consumption combined with the stratification of the water column, which reduced the vertical flow of SSW and, consequently, the supply of nutrients to the surface of this system. The N:P and Si:N ratios and an excess of PO4 > 0.65 μM in surface waters indicated that NO3 was the potential limiting nutrient for primary production in this oceanographic province of the Gulf of California.
Key words: nutrients, enrichment, vertical mixing, Ballenas Channel.
INTRODUCCIÓN
El canal de Ballenas (CB), localizado en la región de las grandes islas del golfo de California (GC), se caracteriza por ser una zona con una dinámica particular y características fisicoquímicas distintas al resto del golfo, por lo que ha sido considerado como una provincia oceanográfica única (Lavín et al. 1997, Lavín y Marinone 2003). Por ejemplo, en el CB se ha registrado el agua superficial más fría (Soto-Mardones et al. 1999, Martínez-Díaz-de-León et al. 2006) y menos salina (Roden 1964, Beron-Vera y Ripa 2000) del GC. Además, en la capa profunda del CB el agua es más cálida y más salina que a la misma profundidad en la región central del GC (Roden 1964, Álvarez-Borrego et al. 1978). Estas características únicas del CB se han atribuido a la fuerte mezcla de la columna de agua asociada a la interacción de las vigorosas corrientes de marea con la compleja batimetría de la zona (Badan-Dangon et al. 1991).
Recientemente, un mecanismo alterno a la mezcla mareal para explicar porqué el CB tiene las aguas superficiales más frías del GC ha sido propuesto por López et al. (2006). Estos autores, por medio de observaciones directas con correntómetros instalados a lo largo de la columna de agua en los umbrales de San Lorenzo (400 m) y CB (600 m), dedujeron que el agua ingresa al CB por la parte profunda pasando sobre ambos umbrales y que la convergencia de estos flujos de entrada subsuperficiales produce una surgencia en el CB. Por otro lado, Marinone (2008) utilizó un modelo numérico tridimensional para simular la circulación profunda alrededor de la isla Ángel de la Guarda, incluyendo el CB, y reprodujo los flujos de agua profundos que ingresan y convergen dentro del CB y que divergen en la superficie, corroborando numéricamente el modelo conceptual propuesto por López et al. (2006). Así, ambas aproximaciones sugieren que el aporte vertical de agua subsuperficial podría ser la causa de la baja temperatura superficial en el CB a lo largo de todo el año.
Desde un punto de vista biológico, el CB es un sistema productivo que presenta altas tasas fotosintéticas a lo largo de todo el año, con valores hasta de 104 mg C m-2 h-1 (Gaxiola-Castro et al. 1995). Sin embargo, estos valores no corresponden a los valores de productividad más altos que se pueden observar en el GC y se ha especulado que esto se debe a que la turbulencia de la columna de agua en el CB causa que el fitoplancton sea transportado verticalmente por debajo de la zona eufótica, donde su desarrollo se ve limitado por la disponibilidad de luz (Santamaría-del-Ángel et al. 1994a). A pesar de las condiciones turbulentas predominantes en el CB, las concentraciones de clorofila a se mantienen elevadas (2.5-6.0 mg m-3) a lo largo de todo el año y no presentan cambios estacionales marcados como en el resto del GC (Santamaría-del-Ángel et al. 1994b).
En el CB, existen pocos estudios sobre la dinámica de los nutrientes. Se ha registrado que las concentraciones superficiales más altas de nutrientes de todo el GC se encuentran en esta zona, y se considera que la mezcla por mareas es el principal mecanismo responsable de este enriquecimiento (Álvarez-Borrego et al. 1978, Gaxiola-Castro et al. 1995, Lavín et al. 1995, Millán-Núñez y Yentsch 2000). En este trabajo se muestra que las concentraciones superficiales de nutrientes inorgánicos en el CB no son altas y constantes a largo de todo el año. El elevado enriquecimiento con nutrientes de la capa superficial del CB se registró principalmente en invierno y estuvo fuertemente asociado con la intensidad dela mezcla vertical de la columna de agua y el flujo vertical del Agua Subsuperficial Subtropical. Aquí presentamos evidencia de que la variabilidad temporal de las concentraciones de los nutrientes de las aguas superficiales del CB, en una escala anual, es una consecuencia de la variación estacional de la mezcla de la columna de agua, de la variabilidad del flujo vertical del Agua Subsuperficial Subtropical y de la actividad biológica que ocurre en esta capa.
MATERIALES Y MÉTODOS
Área de estudio
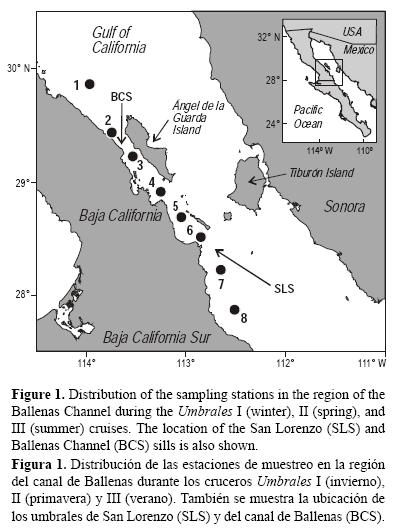
El CB se localiza en la región norte del GC, en la costa oriental de la península de Baja California (fig. 1), y se extiende desde el norte de la isla Ángel de la Guarda (29°34' N) hasta la isla San Lorenzo (28°23' N). El CB es angosto y profundo, mide aproximadamente 125 km de largo y 25 km de ancho, con una profundidad máxima de alrededor de 1600 m (Rusnak et al. 1964). El canal está delimitado al este por la isla Ángel de la Guarda, al oeste por la península de Baja California, al norte por el umbral de CB, que tiene una profundidad de 600 m, y al sur por el umbral de San Lorenzo, que tiene una profundidad de 400 m. La profundidad del umbral de San Lorenzo restringe el intercambio de agua profunda entre el CB y la región centro-sur del GC (López et al. 2006, 2008; Marinone 2007).
Recolección de muestras y análisis de NO3, PO4 y Si(OH)4
Con la intención de determinar la variación estacional de la concentración de los nutrientes inorgánicos disueltos y su relación con la estructura de la columna de agua, se realizaron tres cruceros oceanográficos en la región del CB a bordo del B/O Francisco de Ulloa: Umbrales I se realizó en el invierno de 2002 (17 a 22 de marzo), Umbrales II en la primavera de 2003 (12 a 16 de mayo) y Umbrales III en el verano de 2004 (2 a 6 de septiembre). La distribución de ocho estaciones se ubicó de acuerdo con el gradiente espacial de la intensidad de la mezcla vertical a lo largo del CB, partiendo del norte del umbral de CB hasta el sur del umbral de San Lorenzo (fig. 1). Durante el crucero de verano, no se recolectaron muestras en la estación 8 debido a la presencia de una tormenta tropical al sur de las grandes islas (fig. 1).
Las muestras de agua de mar se recolectaron en cada estación, desde la superficie hasta 1000 m (donde la profundidad lo permitió), utilizando una roseta equipada con seis botellas Niskin de 5 L de capacidad y un CTD modelo SeaBird 911 Plus. Las muestras para el análisis de los nutrientes se recolectaron directamente de las botellas Niskin y se pasaron a través de filtros de fibra de vidrio GF/F Whatman de 24 mm de diámetro con un poro nominal de 0.7 ^m. Se almacenaron en botellas de polietileno de 250 mL, se congelaron inmediatamente y se mantuvieron a -20 °C hasta el momento de su análisis en el laboratorio. Además, se tomaron muestras de agua para el análisis de salinidad en botellas de borosilicato. En el laboratorio, la salinidad se determinó a partir de la medición de la conductividad con un salinómetro Guildline modelo 3400B calibrado con agua de mar estándar de la IAPSO (por sus siglas en inglés, Asociación Internacional para las Ciencias Físicas del Océano). La cuantificación de los nitratos (NO3), fosfatos (PO4) y silicatos (Si(OH)4) se realizó por colorimetría (Gordon et al. 1993) utilizando un autoanalizador de flujo segmentado modelo Skalar SAN Plus. Los límites de detección del análisis de NO3, PO4 y Si(OH)4 fueron de 0.05, 0.07 y 0.20 μM, respectivamente.
Procesamiento de datos
Con la finalidad de evaluar la intensidad de la mezcla vertical de la columna de agua e identificar las zonas con mayor mezcla en el CB, se calculó el parámetro de estratificación (Φ) de acuerdo con la ecuación descrita por Simpson y Bowers (1981):

donde Φ es la cantidad de energía (J m-3) requerida para mezclar la columna de agua (el valor equivale a cero para una columna completamente mezclada y aumenta con la estratificación), h es la profundidad máxima (m) de integración de la columna de agua, p es la densidad del agua (kg m-3) a cada profundidad de recolecta, 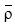 es la densidad promedio para el intervalo de profundidad de 0 a h, g es la aceleración (m s-2) debido a la fuerza de gravedad, z es la coordenada vertical (m) y dz es el incremento de profundidad. En este trabajo seleccionamos h = 100 m debido a que el desarrollo del fitoplancton está asociado con la disponibilidad de los nutrientes en esta capa. Adicionalmente, en el CB se han cuantificado niveles de clorofila a relativamente altos (0.2 a 0.5 mg m-3) desde la superficie hasta los 75-100 m de profundidad (Millán-Núñez y Yentsch 2000).
es la densidad promedio para el intervalo de profundidad de 0 a h, g es la aceleración (m s-2) debido a la fuerza de gravedad, z es la coordenada vertical (m) y dz es el incremento de profundidad. En este trabajo seleccionamos h = 100 m debido a que el desarrollo del fitoplancton está asociado con la disponibilidad de los nutrientes en esta capa. Adicionalmente, en el CB se han cuantificado niveles de clorofila a relativamente altos (0.2 a 0.5 mg m-3) desde la superficie hasta los 75-100 m de profundidad (Millán-Núñez y Yentsch 2000).
Para expresar de manera cuantitativa la forma de la distribución vertical de los nutrientes e identificar su variabilidad temporal, se calculó el gradiente vertical:

donde ΔN es la diferencia en concentración entre 0 y 100 m de profundidad, que es la capa donde la concentración de los nutrientes puede variar debido a la captación del fitoplancton (Millán-Núñez y Yentsch 2000). Así, los valores de ΔN que tiendan a cero indicarán perfiles de nutrientes verticalmente homogéneos, mientras que los valores de ΔN significativamente mayores que cero indicarán una distribución vertical más pronunciada (tipo nutriente), reflejando una columna agua más estratificada. Finalmente, con el propósito de estudiar la variabilidad estacional de la disponibilidad de los nutrientes en la capa superficial, se calculó la concentración promedio del nutriente integrada en los primeros 100 m de acuerdo con la siguiente ecuación:

donde  es la concentración (μM) promedio del nutriente integrada en la columna de agua, N es la concentración del nutriente medida a cada profundidad, dz es la diferencia de profundidad entre las concentraciones medidas y h es la profundidad hasta donde se realizó la integración (100 m).
es la concentración (μM) promedio del nutriente integrada en la columna de agua, N es la concentración del nutriente medida a cada profundidad, dz es la diferencia de profundidad entre las concentraciones medidas y h es la profundidad hasta donde se realizó la integración (100 m).
RESULTADOS
Distribución vertical de las masas de agua en el canal de Ballenas
El análisis hidrográfico de la región del CB indicó que en el invierno, el verano y la primavera se detectaron únicamente dos de las tres masas de agua principales registradas para los primeros 1000 m de la zona centro-norte del GC (Bray y Robles 1991, Torres-Orozco 1993). La isohalina de 35.0 indicó que el Agua del Golfo de California (AGC: S > 35.0 y T >12.0 °C) ocupó la capa superficial y que por debajo de ella se encontró el Agua Subsuperficial Subtropical (ASS: 34.5 < S < 35.0 y 9.0 °C < T < 18.0 °C) llenando la cuenca profunda (fig. 2). También se observó que el Agua Intermedia del Pacífico (AIP: 34.5 < S < 34.8 y 4.0 °C < T < 9.0 °C) no penetró en esta cuenca ya que quedó restringida al sur del CB por el umbral de San Lorenzo (fig. 2a-d).
La variabilidad estacional de la temperatura y de la salinidad durante los tres cruceros fue mayor en los primeros 200 m de profundidad (figs. 2, 3d-e). Las aguas superficiales más frías (15.50 ± 1.70 °C, promedio ± error estándar) y menos salinas (35.16 ± 0.13) se registraron durante el crucero de invierno, mientras que las aguas más cálidas (28.30 ± 1.00) y salinas (35.50 ± 0.21) se detectaron en el verano. Por debajo de esta capa superficial, entre los 400 y 1000 m de profundidad, la variación vertical y la variabilidad estacional de la temperatura y la salinidad fue muy pequeña (<1.10 °C y <0.10, respectivamente), indicando que la parte profunda del CB presentó una columna de agua mucho más homogénea (figs. 2, 3d-e). Este resultado contrastó con lo observado en la estación 8, ubicada al sur del umbral de San Lorenzo, la cual presentó una marcada estratificación de la columna de agua. A una profundidad de 400 m, por ejemplo, el agua fue más fría (~3.5 °C) y menos salina (~0.31) con respecto al promedio de las estaciones del interior del CB debido a la presencia del Agua Intermedia del Pacífico (fig. 2).
Variación temporal de la mezcla vertical en el canal de Ballenas
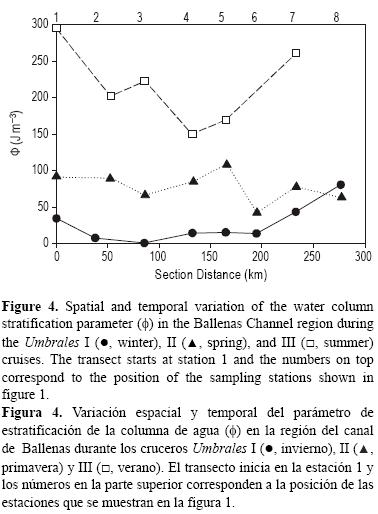
La variación espacial y temporal del parámetro de estratificación calculado para el CB se presenta en la figura 4. Durante el invierno, la capa de los 100 m superficiales del CB (estaciones 3 a 6) se encontró totalmente mezclada, con valores de Φ que variaron entre 0.7 y 15.5 J m-3. Sin embargo, en la primavera el Φ osciló entre 42 y 108 J m-3 debido a la ganancia de calor del agua desde la atmosfera en esta época de transición (Martínez-Díaz-de-León et al. 2013), lo que favoreció la formación de una termoclina más estable (Castro et al. 1994). Durante el verano, a pesar de que la mezcla asociada a las mareas varía con una frecuencia quincenal y se presenta en la región del CB a lo largo de todo el año (Marinone y Lavín 2003, Martínez-Díaz-de-León et al. 2006), la turbulencia del agua disminuyó sustancialmente y se obtuvieron los valores máximos (150-223 J m-3) del Φ, lo cual se reflejó en una columna de agua muy estable (fig. 4). Trabajos previos han mostrado que la estratificación de la columna de agua que se observa durante el verano en el GC se debe al predominio de las fuerzas de flotabilidad asociada principalmente a la ganancia de calor del agua desde la atmósfera (Lavín y Organista 1988, Castro et al.1994).
Distribución vertical de los nutrientes en el canal de Ballenas
Las distribuciones verticales de los tres nutrientes en el CB presentaron el perfil típico del océano abierto, con concentraciones bajas en la superficie y altas hacia la capa profunda, y al igual que la temperatura y la salinidad, los niveles de nutrientes en la capa superficial del CB mostraron una variación estacional muy marcada (fig. 3). Las concentraciones superficiales más altas se detectaron en el invierno (NO3: 17.0-20.8 μM; PO4: 2.0-2.4 μM; Si(OH)4: 35.6-40.2 μM) y las más bajas en el verano (NO3: 0.13-0.66 μM; PO4: 0.83-1.11 μM; Si(OH)4: 3.3-6.4 μM). En todas las campañas se presentó un gradiente superficial espacial definido, con valores superficiales más altos en el CB (estaciones 3 a 6) que los registrados al norte o al sur del canal (fig. 5). En invierno, por ejemplo, en el CB los niveles de NO3, PO4 y Si(OH)4 fueron de dos a tres veces mayores que los medidos al sur del umbral de San Lorenzo (fig. 5a-b). En contraste a la variación en la capa superficial, en la parte profunda por debajo de los 200 m la variabilidad estacional de la concentración de los tres nutrientes no mostró cambios significativos. Entre los 200 y 500 m de profundidad, en el interior del CB, por ejemplo, las concentraciones de los tres nutrientes variaron relativamente poco entre los tres cruceros: 22 μM < NO3 < 24 μM, 2.6 μM < PO4 < 2.8 μM y 50 μM < Si(OH)4 < 55 μM (fig. 3a-c).
DISCUSIÓN
Los resultados del presente trabajo muestran que existió una clara variabilidad estacional en la concentración de los nutrientes en los primeros 200 m del CB. A diferencia de trabajos previos (Álvarez-Borrego et al. 1978, Lavín et al. 1995, Millán y Yentsch 2000), este trabajo muestra que el enriquecimiento de la capa superficial en el CB fue más intenso durante el invierno, cuando la columna de agua se encontró verticalmente más mezclada. Como se argumenta a continuación, proponemos que este enriquecimiento se debe a que en el invierno existe un mayor flujo de ASS hacia el interior del GC y, en consecuencia, hacia el CB, además de la intensificación de la mezcla vertical de la columna de agua que promueve un aumento de la proporción de ASS que arriba a la capa superficial. En contraste, durante el verano, bajo condiciones mucho más cálidas y con una columna de agua muy estable, se produce una disminución en la concentración superficial de estos nutrientes debido a una combinación de consumo fitoplanctónico y un reducido y limitado flujo vertical de ASS.
Transporte horizontal de ASS en el Golfo de California y el flujo vertical de ASS en el canal de Ballenas
La variación estacional de la posición del límite superior del ASS con respecto a la superficie del CB depende del flujo estacional del ASS hacia el interior del GC y de la intensidad de la mezcla vertical de la columna de agua. Aunque no es el propósito de este trabajo demostrar cuantitativamente cuál es el mecanismo físico responsable del movimiento vertical del ASS en el CB, existen evidencias de este transporte vertical en esta provincia oceanográfica (ver también Hernández-Ayón et al. (2013) en este número). Varios reportes indican que el GC presenta una circulación termohalina muy peculiar (e.g., Lavín y Marinone 2003). Producto de la pérdida de agua y la ganancia de calor, el AGC se genera en la región norte del GC y se transporta hacia el sur en forma de una lengüeta de agua salina (>35.0). Para mantener la continuidad de volumen del GC y compensar este flujo de agua que se moviliza entre los 50 y 250 m superficiales desde el norte del GC, ocurre también un flujo hacia el norte de agua intermedia entre 250 y 500 m de profundidad (Bray 1988). Este transporte ocurre de manera sistemática a lo largo de todo el año; sin embargo, durante el invierno, al intensificarse los vientos del noroeste, el AGC se desplaza hacia la boca del golfo y el Agua Superficial Ecuatorial se retrae hacia el exterior del GC (Álvarez-Borrego y Schwartzlose 1979, Lavín et al. 1997). Esto produce una intensificación del flujo de ASS hacia el interior del GC e impacta la parte norte del mismo incluyendo la región de las grandes islas. Estimaciones recientes indican que este flujo horizontal subsuperficial de ASS produce un intercambio vertical anual promedio de aproximadamente 0.23 ± 0.02 Sv en el interior del GC (Álvarez-Borrego y Giles-Guzmán 2012).
El ascenso vertical de las isopicnas subsuperficiales es evidencia de la intensificación del flujo horizontal de ASS que ocurre durante el invierno en el GC y que afecta la región del CB. En este trabajo se define el límite superior de la ASS por la isopicna de 26.3 kg m-3 (temperatura y salinidad promedio ± error estándar de 13.50 ± 0.16 °C y 35.00 ± 0.04, respectivamente), lo cual es consistente con Torres-Orozco (1993) quien delimitó la frontera entre el ASS y el AGC por una salinidad de 35.00. Durante el verano, la isopicna de 26.3 kg m-3 se detectó a 305 ± 17 m, en la primavera a 176 ± 24 m y en el invierno mucho más somera a 80 ± 50 m de profundidad (figs. 3f, 5). Este desplazamiento vertical en una escala estacional de la isopicna de 26.3 kg m-3 lo interpretamos como un ascenso del límite superior de la ASS y, en consecuencia, como un incremento en el flujo vertical y de la proporción de esta masa de agua arribando a la capa superficial del CB. Una segunda evidencia de este ascenso es la disminución de la salinidad en la capa superficial de verano (35.51 ± 0.20) a invierno (35.05 ± 0.02; figs. 2a, 2e, 3d). Esta disminución no puede explicarse por aportes de agua dulce debido a que en la región del CB la lluvia es prácticamente nula y no existen aportes de agua dulce importantes desde la zona continental. Las salinidades superficiales bajas de invierno tampoco pueden explicarse por un proceso advectivo de las aguas superficiales desde el norte del GC debido a que las aguas en esa región son mucho más salinas que las del CB (fig. 2a). Por lo tanto, el agua con menor salinidad debe provenir de la parte subsuperficial del CB.
Todas estas evidencias apuntan hacia la existencia de un flujo vertical de ASS que tiene como resultado un incremento de la concentración de los nutrientes en la capa superficial del CB. En un estudio sobre la dinámica física del CB, López et al. (2006) realizaron mediciones de corrientes en la columna de agua del umbral de San Lorenzo y del umbral de CB por un periodo de seis a ocho meses y, para poder explicar el flujo medio, dedujeron que el ASS ingresa al interior del CB por la parte profunda pasando sobre ambos umbrales. Según estos autores, para compensar estos flujos de entrada subsuperficiales, se produce una surgencia en el CB (0.18 Sv) con una velocidad vertical media de 5.8 m d-1. Sin embargo, aún queda por resolver de manera inequívoca cuales son las causas responsables del aumento del flujo vertical de ASS que ocurre en el invierno en el CB.
Influencia de la mezcla vertical sobre la concentración y la distribución vertical de nutrientes en el canal de Ballenas
En la región central del GC, la mezcla vertical asociada a la marea juega un papel relevante en controlar la estructura de la columna de agua (e.g., Simpson et al. 1994, Argote et al. 1995), el contenido de calor (Paden et al. 1991, 1993), la intensidad de los procesos biológicos (e.g., Gaxiola-Castro et al. 1995) y la composición química de sus aguas (Álvarez-Borrego et al. 1978; Delgadillo-Hinojosa et al. 2001, 2006; Hernández-Ayón et al. 2007; Segovia-Zavala et al. 2010; 2011). La distribución vertical de los nutrientes registrados en este trabajo sugiere que la mezcla vertical de la columna de agua ejerce un efecto directo sobre la forma del perfil y controla la concentración promedio de los nutrientes en la capa superficial del CB. Por ejemplo, las distribuciones verticales fueron mucho más homogéneas (promedio ± error estándar de ΔN = 0.07 + 0.02 μM m-1; ΔP = 0.006 + 0.002 μM m-1; ΔSi = 0.16 + 0.04 μM m-1) bajo condiciones de mezcla en el invierno, mientras que durante el verano, cuando la columna de agua se estratificó, los perfiles de los tres nutrientes fueron más pronunciados (ΔN = 0.13 +0.02 μM m-1, ΔP = 0.009 + 0.002 μM m-1, ΔSi = 0.24 +0.05 μM m-1; fig. 3a-c). De igual manera, existió una relación inversa entre la intensidad de la mezcla vertical (Φ) y la concentración de NO3, PO4 y Si(OH)4 integrada en los primeros 100 m (fig. 6). Las concentraciones promedio integradas más altas (18.2 + 1.1 μM de NO3, 2.36 + 0.08 μM de PO4 y 35.4 + 3.2 μM de Si(OH)4) se detectaron en el invierno cuando la columna de agua estuvo bien mezclada, mientras que las más bajas (7.1 + 1.0 μM de NO3 1.22 + 0.06 μM de PO4 y 16.0 + 2.1 μM de Si(OH)4) se registraron en el verano asociadas a condiciones de estratificación (tabla 1, fig. 4). Mas del 66% de la variabilidad temporal de la concentración de los nitratos, fosfatos y silicatos se explicó por la variabilidad temporal de la mezcla vertical en el CB (fig. 6), sugiriendo que la agitación de la columna de agua en una escala de tiempo estacional (fig. 4) juega también un papel muy importante en controlar la disponibilidad de los nutrientes en la capa superficial del CB. Aunque desconocemos el mecanismo físico que promueve la intensificación de la mezcla vertical en invierno, una posible explicación podría estar dada por la mezcla vertical asociada a la pérdida de calor del agua hacia la atmosfera en la región centro norte del GC (Castro et al. 1994, Beron-Vera y Ripa 2000). Este proceso de pérdida de calor del agua hacia la atmósfera en la zona del CB podría provocar un proceso de convección de la columna de agua, lo que facilitaría el transporte vertical de agua y explicaría, a su vez, el enriquecimiento invernal de nutrientes en la capa superficial del CB. Evidencia preliminar para la zona del CB que apoya esta hipótesis ha sido dada por Martínez-Díaz-de-León et al. (2006, 2013), quienes instalaron termógrafos en seis bahías localizadas a lo largo del CB y estudiaron la variación estacional de la temperatura superficial por un periodo de un año. Los datos de estos autores indican que durante el otoño ocurre una intensa disminución de la temperatura del agua, la cual fue atribuida a la pérdida de calor hacia la atmosfera. En contraste, durante el verano, la ganancia de calor del agua del CB (Paden et al. 1993, Martínez-Díaz-de-León et al. 2013) genera una columna de agua muy estratificada, lo cual impone un control físico al flujo vertical de la ASS y, en consecuencia, representa una restricción en el aprovisionamiento de nutrientes a la capa superficial de CB.


Efecto de los procesos biológicos sobre la distribución de los nutrientes en el canal de Ballenas
Los principales procesos biológicos que afectan la concentración de NO3, PO4 y Si(OH)4 en el océano son la asimilación por parte del fitoplancton en la capa superficial y, en la capa profunda, la respiración de la materia orgánica en el caso de los NO3 y PO4 (Tyrrell y Law 1997) y la disolución del sílice biogénico en el caso de los Si(OH)4 (Van Cappellen et al. 2002). Una aproximación que utilizamos para estudiar el efecto de los procesos biológicos sobre la concentración de nutrientes fue realizar un análisis de regresión lineal entre los diferentes nutrientes y, a partir de las pendientes, inferir sus tasas de consumo y regeneración. Las relaciones N:P y Si:N estimadas para la columna de agua del CB se presentan en la tabla 2. Considerando todo el conjunto de datos de las tres épocas de muestreo, las tasas de consumo y regeneración de N:P y Si:N fueron de 12.1 (±0.4):1 y 2.0 (±0.1):1, respectivamente. Estos valores de las razones N:P y Si:N se encuentran en el intervalo de valores documentados previamente para el GC. Por ejemplo, Torres-Valdés (2000) estimó razones N:P que variaron de 12.7 en invierno a 9.3 en verano, mientras que Pichevin et al. (2012) registraron una razón Si:N >1.8 y la atribuyeron como característica del GC.
En general, las tasas de consumo y regeneración de Si:N en el CB variaron de 2.6 (±0.1):1 en invierno a 1.9 (±0.1):1 en verano (tabla 2), indicando que el Si(OH)4 se encontró en exceso en relación al N en las tres épocas del año. Sin embargo, cabe notar que los valores de la razón N:P en la región del CB de invierno y verano (tabla 2) son menores que la razón de 16:1 propuesta por Redfield et al. (1963). Las proporciones N:P bajas se han registrado para sistemas marinos asociados a ambientes subóxicos, donde se favorece la desnitrificación (Tyrrel y Lucas 2002) y/o donde el grupo predominante del fitoplancton es el de las diatomeas (Arrigo et al. 1999), las cuales se caracterizan por ser fuertes consumidoras de N (Arrigo et al. 2002). Los valores de oxígeno (>30 | M) registrados en la parte profunda de la columna de agua correspondiente al ASS en el CB (datos no presentados) sugieren que difícilmente el proceso de desnitrificación pueda estar llevándose a cabo en la columna de agua de esta región. Si el proceso de desnitrificación local puede ser descartado, lo más probable es que, en su viaje hacia la región de las grandes islas, el ASS llegue al CB con un exceso de PO4 (tabla 2) y una deficiencia de NO3 por haber sido utilizado en la región subóxica ubicada al sur del GC (Altabet et al. 1999, Delgadillo-Hinojosa et al. 2006). Por otro lado, el aporte continuo de nutrientes hacia la superficie del CB a través de la mezcla vertical, especialmente en invierno y primavera, favorece que se desarrollen organismos capaces de proliferar en ambientes con fuerte agitación del agua. Las diatomeas son un grupo de fitoplancton con tasas de división celular rápidas que, en presencia de un incremento de la concentración de nutrientes, se desarrollan y prosperan aún en aguas con fuerte agitación (Thunell et al. 1996). Diversos estudios han documentado que la composición de la comunidad fitoplanctónica juega un papel central en determinar la magnitud de la razón N:P del agua. Por ejemplo, en el mar de Ross se observó que cuando hubo un florecimiento de diatomeas la razón N:P fue baja (promedio ± error estándar = 9.69 ± 0.33), mientras que en presencia de otra especie de fitoplancton (el dinoflagelado Phaeocystis Antarctica) la razón N:P fue alta (19.2 ± 0.6) (Arrigo et al. 1999). Nuestros resultados indican que la concentración de los Si(OH)4 en la capa superficial del CB disminuyó significativamente de invierno a verano (fig. 3), sugiriendo que su decremento se debió al consumo por parte de las diatomeas. Además, tomando en cuenta que el GC (Sancetta 1995) y el CB en particular (Millán-Núñez y Yentsch 2000) se caracterizan por una gran abundancia de este grupo fitoplanctónico, la respuesta más probable a la baja razón N:P registrada en este trabajo es el efecto combinado del intenso consumo de N asociado a la alta abundancia de las diatomeas y a la presencia de agua desnitrificada proveniente de la región sur del GC.
El intercepto negativo en la razón N:P, el intercepto positivo en la razón Si:N y el exceso >0.65 μM de PO4 calculado para el conjunto de datos de los tres cruceros (tabla 2) indican también que el NO3 es el nutriente potencialmente limitante de la producción primaria para las especies eucariontes del fitoplancton que prosperan en la región del CB, lo cual es consistente con otros trabajos que han mostrado condiciones de limitación de N en el GC. Recientemente, White et al. (2007) documentaron que bajo condiciones de verano, se registran en el GC tasas relativamente altas de fijación de N2 (20 a 250 μmol m-2 d-1), asociadas a una capa de mezcla bien marcada y con concentraciones de NO3 extremadamente bajas. Estas tasas de fijación de N constituyen una fuente importante de N fijado para este sistema y, de acuerdo con White et al. (2007), pueden contribuir con el 35% al 48% de la demanda de nitrógeno del fitoplancton de la región central del GC. En el caso del CB, esta condición de limitación de N podría adquirir mayor relevancia durante el verano, cuando existe menor disponibildad de N (figs. 3a, 5e) en las aguas cálidas (T > 27 °C; figs. 2f, 3e) superficiales. Durante la campaña de verano, las concentraciones superficiales de NO3variaron de 0.13 a 0.66 μM 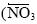 = 0.38 ± 0.12 μM) a lo largo del canal y se encontraron ligeramente por encima del límite de detección del método. Estos niveles de NO3 bajos combinado con el exceso de PO4 y las altas temperaturas registradas en el CB podrían favorecer la aparición, el desarrollo y la proliferación de especies fitoplanctónicas con la habilidad para fijar el N2 (e.g., Capone et al. 2005). Una implicación de estos resultados es que bajo condiciones de verano, la fijación del N2 en el GC podría estar ocurriendo de manera más extensa de lo que se había pensado antes, incluso en lugares caracterizados por una dinámica física muy intensa y altas concentraciones de nutrientes de manera cuasi permanentes, como es el caso del CB en la región centro-norte del GC.
= 0.38 ± 0.12 μM) a lo largo del canal y se encontraron ligeramente por encima del límite de detección del método. Estos niveles de NO3 bajos combinado con el exceso de PO4 y las altas temperaturas registradas en el CB podrían favorecer la aparición, el desarrollo y la proliferación de especies fitoplanctónicas con la habilidad para fijar el N2 (e.g., Capone et al. 2005). Una implicación de estos resultados es que bajo condiciones de verano, la fijación del N2 en el GC podría estar ocurriendo de manera más extensa de lo que se había pensado antes, incluso en lugares caracterizados por una dinámica física muy intensa y altas concentraciones de nutrientes de manera cuasi permanentes, como es el caso del CB en la región centro-norte del GC.
CONCLUSIONES
La concentración superficial y la distribución vertical de los nutrientes inorgánicos disueltos (PO4, NO3 y Si(OH)4) en el CB varía estacionalmente y está controlada por la variabilidad en la intensidad de la mezcla de la columna de agua y del aporte del ASS hacia la superficie asociada a la circulación termohalina del GC. El enriquecimiento superficial de nutrientes se registró bajo condiciones de invierno y primavera, cuando la columna de agua se encontró verticalmente mezclada y la proporción de ASS que arribó a la capa superficial fue mayor. Por otro lado, las concentraciones relativamente bajas de nutrientes que se registraron en verano se debieron a la fuerte actividad biológica y a un aporte limitado de ASS hacia la capa superficial del CB asociado a la intensa estratificación de la columna de agua. La razón N:P varió estacionalmente y fue menor que 16:1, indicando que el NO3 es el nutriente potencialmente limitante de la producción primaria que tiene lugar en el CB. Además, se estimó que las aguas superficiales poseen un exceso de PO4> 0.65 μM, indicando que esta agua está relativamente empobrecida en NO3, ya que proviene de la región al sur del GC donde ocurre la desnitrificación. Este aporte de agua desnitrificada pudiera permitir el desarrollo de especies fitoplanctónicas fijadoras de N durante el verano, y mantener así, a lo largo de todo el año, la alta productividad primaria del CB.
AGRADECIMIENTOS
Este trabajo fue financiado por el Consejo Nacional de Ciencia y Tecnología (CONACYT, proyectos G33464-T, 46523 y 46576) y la Universidad Autónoma de Baja California (UABC, proyecto 403/1/5/10). Agradecemos a los dos revisores anónimos sus comentarios y sugerencias, las cuales mejoraron significativamente la versión final del artículo.
REFERENCES
Altabet MA, Pilskaln C, Thunell R, Pride C, Sigman D, Chavez F,Francois R. 1999. The nitrogen isotope biogeochemistry of sinking particles from the margin of the Eastern North Pacific. Deep-Sea Res. 46: 655-679. http://dx.doi.org/10.1016/S0967-0637(98)00084-3 [ Links ]
Álvarez-Borrego S, Schwartzlose RA. 1979. Water masses of the Gulf of California. Cienc. Mar. 6: 43-63. [ Links ] Álvarez-Borrego S, Giles-Guzmán AD. 2012. Opal in Gulf of California sediments as a tool to estimate the vertical component of water exchange between the Gulf and the Pacific Ocean. Bot. Mar. 55: 161-168. http://dx.doi.org/10.1515/bot.2011.104 [ Links ]
Álvarez-Borrego S, Rivera JA, Gaxiola-Castro G, Acosta-Ruiz JM, Schwartzlose RA. 1978. Nutrientes en el Golfo de California. Cienc. Mar. 5: 53-71. [ Links ]
Argote ML, Amador A, Lavín MF, Hunter JR. 1995. Tidal dissipation and stratification in the Gulf of California. J. Geophys. Res. 100(C8): 16103-16118.http://dx.doi.org/10.1029/95JC01500 [ Links ]
Arrigo KR, Robinson DH, Worthen DL, Dunbar RB, DiTullio GR, Van Woert M, Lizotte MP. 1999. Phytoplankton community structure and the drawdown of nutrients and CO2 in the Southern Ocean. Science 283: 365-367. http://dx.doi.org/10.1126/science.283.5400.365 [ Links ]
Arrigo KR, Dunbar RB, Lizotte MP, Robinson DH. 2002. Taxon-specific differences in C/P and N/P drawdown for phytoplankton in the Ross Sea, Antarctica. Geophys. Res. Lett. 29:(44-1)-(44-4).http://dx.doi.org/10.1029/2002GL015277 [ Links ]
Badan-Dangon A, Hendershott MC, Lavín MF. 1991. Underway Doppler current profiles in the Gulf of California. Eos Trans. AGU 72: 209-218. [ Links ]
Beron-Vera FJ, Ripa P. 2000. Three-dimensional aspects of the seasonal heat balance in the Gulf of California. J. Geophys. Res. 105 (C5), 11441-11457.http://dx.doi.org/10.1029/2000JC900038 [ Links ]
Bray NA. 1988. Thermohaline circulation in the Gulf of California. J. Geophys. Res. 93(C5): 4993-5020. http://dx.doi.org/10.1029/JC093iC05p04993 [ Links ]
Bray N, Robles JM. 1991. Physical oceanography of the Gulf of California. In: Dauphin JP, Simoneit BR (eds.), The Gulf and Peninsular Province of the Californias. Mem. Am. Assoc. Pet. Geol. 47: 511-553. [ Links ]
Capone DG, Burns JA, Montoya JP, Subramaniam A, Mahaffey C, Gunderson T, Michaels AF, Carpenter EJ. 2005. Nitrogen fixation by Trichodesmium spp.: An important source of new nitrogen to the tropical and subtropical North Atlantic Ocean. Global Biogeochem. Cycles 19, GB2024http://dx.doi.org/10.1029/2004GB002331 [ Links ]
Castro R, Lavín MF, Ripa P. 1994. Seasonal heat balance in the Gulf of California. J. Geophys. Res. 99: 3249-3261 http://dx.doi.org/10.1029/93JC02861 [ Links ]
Delgadillo-Hinojosa F, Macías-Zamora JV, Segovia-Zavala JA, Torres-Valdés S. 2001. Cadmium enrichment in the Gulf of California. Mar. Chem. 75: 109-122. http://dx.doi.org/10.1016/S0304-4203(01)00028-7 [ Links ]
Delgadillo-Hinojosa F, Segovia-Zavala JA, Huerta-Díaz MA, Atilano-Silva H. 2006. Influence of geochemical and physical processes on the vertical distribution of manganese in Gulf of California waters. Deep-Sea Res. I 53: 1301-1319. http://dx.doi.org/10.1016/j.dsr.2006.06.002 [ Links ]
Gaxiola-Castro G, García-Córdova J, Valdez-Holguín JE, Botello-Ruvalcaba M. 1995. Spatial distribution of chlorophyll a and primary productivity in relation to winter physical structure in the Gulf of California. Cont. Shelf Res. 15: 1043-1059. http://dx.doi.org/10.1016/0278-4343(94)00071-T [ Links ]
Gordon LI, Jennings JC Jr, Ross AA, Krest JM. 1993. A suggested protocol for continuous flow automated analysis of seawater nutrients (phosphate, nitrate, nitrite and silicic acid) in the WOCE Hydrographic Program and the Joint Global Ocean Fluxes Study. WOCE Hydrographic Program Office, Methods Manual WHPO 91-1, 55 pp. [ Links ]
Hernández-Ayón JM, Delgadillo-Hinojosa F, Camiro-Vargas T, López-Mariscal M. 2007. Estudio de las variables del CO2 en la región de las grandes islas en el Golfo de California en condiciones de invierno. In: Hernández-de-la-Torre B, Gaxiola-Castro G (eds.), Carbono en Ecosistemas Acuáticos de México. Instituto Nacional de Ecología, México, pp. 59-73. ISBN: 978968-817-855-3. [ Links ]
Hernández-Ayón JM, Chapa-Balcorta C, Delgadillo-Hinojosa F, Camacho-Ibar VF, Huerta-Díaz MA, Santamaria-del-Angel E, Galindo-Bect S, Segovia-Zavala JA. 2013. Dynamics of dissolved inorganic carbon in the Midriff Islands region of the Gulf of California: Influence of water masses. Cienc. Mar. 39: 65-83.http://dx.doi.org/10.7773/cm.v39i2.2243 [ Links ]
Lavín MF, Organista S. 1988. Surface heat flux in the northern Gulf of California. J. Geophys. Res. 93: 14033-14038. http://dx.doi.org/10.1029/JC093iC11p14033 [ Links ]
Lavín MF, Marinone SG. 2003. An overview of the physical oceanography of the Gulf of California. In: Velasco-Fuentes OU et al. (eds.), Nonlinear Processes in Geophysical Fluid Dynamics. Kluwer Academic Publisher, Netherlands, pp. 173-204.http://dx.doi.org/10.1007/978-94-010-0074-1_11 [ Links ]
Lavín MF, Gaxiola-Castro G, Robles JM, Richter K. 1995. Winter water masses and nutrients in the northern Gulf of California. J. Geophys. Res. 100(5): 8587-8605. http://dx.doi.org/10.1029/95JC00138 [ Links ]
Lavin MF, Beier EJ, Badan-Dangon ARF. 1997. Estructura hidrográfica y circulación del Golfo de California: escalas estacional e interanual. In: Lavín MF (ed.), Contribuciones a la oceanografía física en México. Monografía #3. Unión Geofísica Mexicana. 139-169 p. [ Links ]
López M, Candela J, Argote ML. 2006. Why does the Ballenas Channel have the coldest SST in the Gulf of California? Geophys. Res. Lett. 33: L11603http://dx.doi.org/10.1029/2006GL025908. [ Links ]
López M, Candela J, García J. 2008. Two overflows in the northern Gulf of California. J. Geophys. Res. 113.http://dx.doi.org/10.1029/2007JC004575. [ Links ]
Marinone SG. 2007. A note on "Why does the Ballenas Channel have the coldest SST in the Gulf of California?" Geophys. Res. Lett. 34: L02607http://dx.doi.org/10.1029/2006GL028589. [ Links ]
Marinone SG. 2008. On the three-dimensional numerical modeling of the deep circulation around Angel de la Guarda Island in the Gulf of California. Estuar. Coast. Shelf Sci. 80: 430-434. [ Links ]
Marinone SG, Lavín MF. 2003. Residual flux and mixing in the large islands region of the central Gulf of California. In: Velasco-Fuentes OU et al. (eds.), Nonlinear Processes in Geophysical Fluid Dynamics. Kluwer Academic Publisher, Netherlands, pp. 213-236. http://dx.doi.org/10.1007/978-94-010-0074-1_13 [ Links ]
Martínez-Díaz-de-León A, Pacheco-Ruíz I, Delgadillo-Hinojosa F, Zertuche-González JA, Chee-Barragán A, Blanco-Betancourt R, Guzmán-Calderón JM, Gálvez-Telles A. 2006. Spatial and temporal variability of the sea surface temperature in the Ballenas-Salsipuedes Channel (central Gulf of California). J. Geophys. Res. 111: C02008/1-7. http://dx.doi.org/10.1029/2005JC002940 [ Links ]
Martínez-Díaz-de-León A, Castro R, Santamaría-del-Ángel E, Pacheco-Ruíz I, Blanco-Betancourt R. 2013. Sea surface heat fluxes and fortnightly modulation of the surface temperature within the Ballenas Channel, Gulf of California. J. Coast. Res. (in press).http://dx.doi.org/10.2112/JCOASTRES-D-12-00189.1 [ Links ]
Millán-Núñez E, Yentsch C. 2000. El Canal de Ballenas, Baja California, como ambiente favorable para el desarrollo delfitoplancton. Hidrobiológica 10: 91-100. [ Links ]
Paden CA, Abbott MR, Winant CD. 1991. Tidal and atmospheric forcing of the upper ocean in the Gulf of California. 1. Seasurface temperature variability. J. Geophys. Res. 96:18337-18359.http://dx.doi.org/10.1029/91JC01597 [ Links ]
Paden CA, Winant CD, Abbott MR. 1993. Tidal and atmospheric forcing of the upper ocean in the Gulf of California. 2. Surfaceheat flux. J. Geophys. Res. 98: 20091-20103.http://dx.doi.org/10.1029/93JC02190 [ Links ]
Pichevin L, Ganeshram RS, Reynolds BC, Prahl F, Pedersen TF, Thunell R, McClymont EL. 2012. Silicic acid biogeochemistry in the Gulf of California: Insights from sedimentary Si isotopes. Paleoceanography 27, PA2201.http://dx.doi.org/10.1029/2011PA002237 [ Links ]
Redfield AC, Ketchum BH, Richards FA. 1963. The influence of organisms on the composition of sea-water. In: Hill MN (ed.), The Sea. Wiley, New York, Vol. 2, pp. 26-77. [ Links ]
Roden GI. 1964. Oceanographic aspects of the Gulf of California. In: Van Andel TH, Shor GG Jr (eds.), Marine Geology of the Gulf of California: A symposium. Mem. Am. Assoc. Pet. Geol. 3: 30-58. [ Links ]
Rusnak GA, Fisher RL, Shepard FP. 1964. Bathymetry and faults of the Gulf of California. In: Van Andel TH, Shor GG Jr (eds.), Marine Geology of the Gulf of California. Mem. Am. Assoc. Pet. Geol. 3: 59-75. [ Links ]
Sancetta C. 1995. Diatoms in the Gulf of California: Seasonal flux patterns and the sediment record for the last 15,000 years. Paleoceanography 10: 67-84.http://dx.doi.org/10.1029/94PA02796 [ Links ]
Santamaría-del-Ángel E, Álvarez-Borrego S, Müller-Karger FE. 1994a. The 1982-1984 El Niño in the Gulf of California as seenin coastal zone color scanner imagery. J. Geophys. Res. 99:7423-7431.http://dx.doi.org/10.1029/93JC02147 [ Links ]
Santamaría-del-Ángel E, Álvarez-Borrego S, Müller-Karger FE. 1994b. Gulf of California biogeographic regions based on coastal color scanner imagery. J. Geophys. Res. 99: 7411-7421.http://dx.doi.org/10.1029/93JC02154 [ Links ]
Segovia-Zavala JA, Lares-Reyes ML, Delgadillo-Hinojosa F, Tovar-Sánchez A, Sañudo-Wilhelmy SA. 2010. Dissolved iron distributions in the central region of the Gulf of California. Deep-Sea Res. I 57: 53-64.http://dx.doi.org/10.1016/j.dsr.2009.10.007 [ Links ]
Segovia-Zavala JA, Delgadillo-Hinojosa F, Lares-Reyes ML, Huerta-Díaz MA, Muñoz-Barbosa A, Santamaría-del-Ángel E, Torres-Delgado EV, Sañudo-Wilhelmy SA. 2011. Vertical distribution of dissolved iron, copper, and cadmium in Ballenas Channel, Gulf of California. Cienc. Mar. 37: 457-469.http://dx.doi.org/10.7773/cm.v37i4A.1967 [ Links ]
Simpson JH, Bowers D. 1981. Models of stratification and frontal movement. Deep-Sea Res. 28: 727-738.http://dx.doi.org/10.1016/0198-0149(81)90132-1 [ Links ]
Simpson JH, Souza AJ, Lavín MF. 1994. Tidal mixing in the Gulf of California. In: Beven KJ, Chatwin PC, Millbank JH (eds.), Mixing and Transport in the Environment. Wiley, London,pp. 169-182. [ Links ]
Soto-Mardones L, Marinone SG, Parés-Sierra A. 1999. Time and spatial variability of sea surface temperature in the Gulf of California. Cienc. Mar. 25: 1-30. [ Links ]
Thunell R, Pride C, Ziveri P, Muller-Karger F, Sancetta C, Murray D. 1996. Plankton response to physical forcing in the Gulf ofCalifornia. J. Plankton Res. 18: 2017-2026.http://dx.doi.org/10.1093/plankt/18.11.2017 [ Links ]
Torres-Orozco E. 1993. Análisis volumétrico de las masas de agua del Golfo de California. MSc thesis, Centro de Investigación Científica y de Educación Superior de Ensenada, México, 80 pp. [ Links ]
Torres-Valdés S. 2000. Distribución espacial y variabilidad temporal de nutrientes en el Golfo de California. MSc thesis, Facultad de Ciencias Marinas, Universidad Autónoma de Baja California,Ensenada, México, 62 pp. [ Links ]
Tyrrell T, Law CS. 1997. Low nitrate: phosphate ratios in the globalocean. Nature 387: 793-796.http://dx.doi.org/10.1038/42915 [ Links ]
Tyrrell T, Lucas MI. 2002. Geochemical evidence of denitrification in the Benguela upwelling system. Cont. Shelf Res. 22:2497-2511.http://dx.doi.org/10.1016/S0278-4343(02)00077-8 [ Links ]
Van Cappellen P, Dixit S, van Beusekom J. 2002. Biogenic silica dissolution in the oceans: Reconciling experimental and field-based dissolution rates. Global Biogeochem. Cycles 16: 231-23-10.http://dx.doi.org/10.1029/2001GB001431 [ Links ]
White AE, Prahl FG, Letelier RM, Popp BN. 2007. Summer surface waters in the Gulf of California: Prime habitat for biological N2 fixation. Global Biogeochem. Cycle 21, GB2017. http://dx.doi.org/10.1029/2006GB002779. [ Links ]

