










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkCiencias marinas
Print version ISSN 0185-3880
Cienc. mar vol.32 n.4 Ensenada Dec. 2006
Nota de Investigación
Comunidad íctica de una franja arrecifal del Parque Nacional Mochima, Venezuela
Fish community of a fringing reef at Mochima National Park, Venezuela
E Méndez de E1, LJ Ruiz1, A Prieto A1, A Torres de J1, A Fariña1, S Sant1, J Barrio2, B Marín2
1 Departamento de Biología, Escuela de Ciencias, Universidad de Oriente, Apartado 245, Cumaná, Estado Sucre, Venezuela. E- mail: lyeelguezabl@cantv.net
2 Instituto Oceanográfico de Venezuela, Universidad de Oriente, Cumaná, Venezuela.
Recibido en agosto de 2005
Aceptado en agosto de 2006.
Resumen
Se analizó la comunidad de peces asociada a una franja arrecifal en Cautaro, Parque Nacional Mochima, Venezuela. Las observaciones y conteo de las especies se realizaron mensualmente, desde diciembre de 2000 hasta mayo de 2002, mediante censos visuales y a una profundidad media de 2 m. Se identificaron 86 especies, pertenecientes a 31 familias. Las familias con mayor número de especies fueron Scaridae (10), Haemulidae y Labridae (9), así como Pomacentridae (7) y Serranidae (6), las cuales, conjuntamente con la única especie de Sparidae, acumularon 77.52% de la abundancia total. Las especies Diplodus argenteus (13.15%), Microspathodon chrysurus (13.00%) y Thalassoma bifasciatum (10.79%) fueron las más abundantes y frecuentes (>50%), por lo que pueden considerarse típicas y características del área estudiada. La diversidad total fue de 4.54 bits ind-1 y osciló entre 3.33 y 4.25 bits ind-1 con un promedio de 3.86 bits ind-1. La equitabilidad varió entre 0.74 y 0.89. El número de individuos por especie, en relación al rango, se ajustó con una regresión lineal altamente significativa (r = 0.976) y un índice de diversidad de la serie logarítmica de a = 14.19. El análisis trófico de la comunidad indicó que las especies carnívoras constituyeron el grupo dominante (63.10%), seguido por las herbívoras (21.40%) y omnívoras (15.50%). No se detectaron diferencias significativas entre el número de especies observadas y el esperado de acuerdo con la distribución Log-normal, lo cual indica una tendencia a la estabilización en relación al número de especies.
Palabras clave: franja arrecifal, estructura comunitaria de peces, Parque Nacional Mochima.
Abstract
The fish community associated with a fringing reef was studied at Cautaro, Mochima National Park, Venezuela. A systematic sampling design based on visual censuses was carried out from December 2000 to May 2002. A total of 86 species belonging to 31 families were identified, dominated by Scaridae (10), Haemulidae and Labridae (9), Pomacentridae (7) and Serranidae (6), which accumulated, together with the sole species of Sparidae, 77.52% of the total abundance. Diplodus argenteus (13.15%), Microspathodon chrysurus (13.00%) and Thalassoma bifasciatum (10.79%) were the most abundant and frequent (>50%) species, and can therefore be considered typical and characteristic of the study area. The total diversity was 4.54 bits ind-1 and oscillated between 3.33 and 4.25 bits ind-1, with an average of 3.86. Equitability varied between 0.74 and 0.89. The relationship between species abundance and rank was significant (r = 0.976) and the diversity index of the log-series based on the number of species was a = 14.19. The trophic analysis of the community indicated that the carnivorous species constituted the dominant group (63.10%), followed by the herbivores (21.40%) and omnivores (15.50%). The normal distribution fitted the species abundance data, indicating a tendency towards stabilization in relation to the number of species.
Key words: fringing reefs, fish community structure, Mochima National Park.
Introducción
Los arrecifes sostienen las comunidades más complejas que existen en el mar, como resultado de su heterogeneidad espacial y temporal, así como de factores físicos y biológicos (Glynn, 1976). En el Parque Nacional Mochima se encuentran las formaciones arrecifales más importantes de la región oriental venezolana, constituidas en un alto porcentaje por arrecifes franjeantes (Pauls 1982, Sant 1999). Estos ecosistemas son muy particulares en su composición, ya que las surgencias periódicas que caracterizan la zona generan altas tasas de turbidez y de sedimentación en la columna de agua, que conllevan al establecimiento de especies de corales distintas a las de arrecifes típicos, y un predominio de organismos capaces de tolerar bajas temperaturas y condiciones de turbidez (Antonius 1980). En tal sentido, son relevantes los estudios que ayuden a discernir los efectos de tales cambios anuales en la composición de las especies.
En Venezuela, las investigaciones sobre peces de ambientes coralinos son recientes. Para el occidente del país Rodríguez y Villamizar (2000) aportaron información sobre la comunidad de peces arrecifales en Playa Mero (Parque Nacional Morrocoy), y Alvarado (2000) trabajó en diferentes zonas arrecifales del mencionado parque. Kurten et al. (2003) y Lasso-Alcalá et al. (2003) analizaron las comunidades ícticas en los fondos coralinos de la Bahía de Turiamo, en la región centro-occidental. En la zona insular oceánica destacan los trabajos de Posada et al. (2003) en Los Roques, y de Fariña et al. (2005) en Los Monjes. En el caso particular del Parque Nacional Mochima, Méndez et al. (2001) y Ruiz et al. (2003) han realizado la evaluación ictiológica de algunos sistemas arrecifales escasamente perturbados, a fin de tener referencia de su diversidad íctica utilizando diferentes métodos de muestreo.
Los peces constituyen uno de los componentes biológicos más importantes dentro de la estructura de esas comunidades, ya que crean una fuerte asociación con el ambiente que los rodea. Dada la importancia de los ecosistemas mencionados, su gran interés biológico y el alto dinamismo y variabilidad que estacionalmente manifiestan, se analizó la estructura de la comunidad de peces asociada a un arrecife del sector Cautaro, del Parque Nacional Mochima.
Materiales y métodos
Este estudio se realizó, entre diciembre de 2000 y mayo de 2002, en una franja arrecifal ubicada en el Parque Nacional Mochima (10°24'30" N, 64°22'30" O), Estado Sucre, Venezuela (fig. 1). El arrecife de franja está ubicado en la localidad de Cautaro, en la parte externa de la Bahía de Mochima, en una ensenada semi-expuesta a la acción directa del viento. Presenta un lecho rocoso en aproximadamente 40% de su superficie y una cubierta de corales en 50%. El área total es de aproximadamente 2000 m2 y se extiende desde 0.5 m hasta 8 m de profundidad, con una pendiente suave (5°). La riqueza de especies coralinas actualmente es de 22 especies, ya que en la marejada de 1999 se perdieron 9 especies de corales (Sant et al. 2003). Los corales mejor representados son Míllepora alcicornis, Colpophyllia natans y Montastrea annularis (Sant 1999).

Para caracterizar la comunidad de peces arrecifales se realizaron censos visuales mensuales, sobre transeptos de 50 m de largo por 2 m de ancho, los cuales estaban ubicados a una profundidad media de 2 m. Éstos se llevaron a cabo siempre a la misma hora y durante una extensión de 50 min, desarrollándose en paralelo a la línea de costa. Las especies se identificaron empleando las descripciones de Humann (1997).
En algunos casos particulares se capturaron ejemplares y fueron identificados mediante las claves de Cervigón (1994, 1996).
Se determinaron los siguientes parámetros comunitarios (Krebs 1989): diversidad mensual y total [utilizando la expresión de Shannon (H') y Equitabilidad (J' = H' /Hmax)], especies dominantes (basado en la abundancia relativa mensual) y especies constantes (considerando las especies frecuentes en más del 50% de los censos). Los datos del número de individuos por especie se relacionaron entre sí usando el logaritmo natural de la abundancia.
Igualmente, se determinó la diversidad a usando los logaritmos naturales de la abundancia de las especies y aplicando el modelo Log-series de Taylor et al. (1976), usando la ecuación ST = α ln (1 + N/α), donde ST es el número de especies y N el número de individuos, parámetro que indica el número de especies importantes en la comunidad.
Las especies identificadas fueron agrupadas según sus hábitos tróficos, con base en los estudios previos de Randall (1967), Sierra et al. (1994), Martínez-Orsatti et al. (2001), Gauthier (2003) y Méndez et al. (2003). Con el objeto de analizar los resultados de abundancia y diversidad obtenidos, se empleó el modelo estadístico de la Distribución Log-normal (Preston 1948), la prueba de Chi-cuadrada para comparar los valores observados con los esperados y el método de la diversidad acumulada de Pielou (1975). En cada salida de campo se determinó la salinidad con un refractómetro (%o), así como la temperatura (°C).
Resultados
Composición y abundancia de especies
Se contabilizó un total de 4929 individuos pertenecientes a 86 especies de peces, incluidas en 31 familias y 10 órdenes (tabla 1). En términos de número de especies, las familias mejor representadas fueron Scaridae (11), Haemulidae (9), Labridae (8), Pomacentridae (7) y Serranidae (6). Los representantes de estas familias, conjuntamente con Sparidae acumularon 77.52% de la abundancia total (fig. 2).

En la familia Scaridae las especies mejor representadas fueron Sparisoma viride (5.54%), Scarus iseri (4.20%) y Scarus vetula (4.10%); en la familia Labridae fueron Thalassoma bifasciatum (10.79%) y Halichoeres bivittatus (3.46%); mientras que en Haemulidae, las especies con mayor importancia numérica fueron Haemulon aurolineatum (7.22%) y H. chrysargyreum (4.30%). En Pomacentridae destacaron Microspatodon chrysurus (13.00%) y Abudeduf saxatilis (5.90%). No obstante, la especie más abundante durante todos los censos fue Diplodus argenteus (13.15%) (fig. 3).

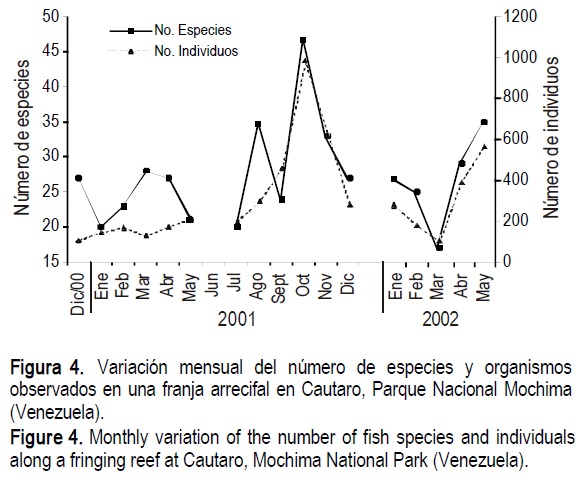
Los números de especies e individuos presentaron fluctuaciones mensuales marcadas, con valores máximos en agosto, octubre y noviembre de 2001 y mayo de 2002. El mayor número de especies observadas y censadas fue de 47 (octubre de 2001) y el mínimo de 17 (marzo de 2002), para un promedio de 27.50 especies. El número de individuos osciló entre 108 en diciembre de 2000 y 985 en octubre de 2001 (fig. 4).
Diversidad de la comunidad
La diversidad de la ictiofauna osciló ente 3.33 y 4.21 bits ind-1 con un promedio de 3.86 bits ind-1. La máxima diversidad se observó en diciembre de 2000 y la mínima en marzo de 2002. La equitabilidad media fue 0.82, observándose su máximo valor (0.89) en diciembre de 2000 y el mínimo (0.74) en noviembre de 2001 (fig. 5). Los datos del número de individuos por especie, en orden decreciente, se ajustaron satisfactoriamente a una regresión lineal altamente significativa (y = -0.033x + 2.433; r = 0.976), con un parámetro a (índice de diversidad) de la serie logarítmica = 14.19, que indica el número de especies importantes.

Especies dominantes y constantes
Con base en la información obtenida de los censos visuales, la abundancia relativa mensual y la frecuencia en los muestreos, se demostró que tres especies aparecieron en más de 50% de las observaciones mensuales realizadas: Thalassoma bifasciatum estuvo presente en todos los meses (100.0%), con una abundancia relativa que osciló entre 12.5% y 32.7%; Microspathodon chrysurus con una frecuencia de 88.9% y abundancia relativa que varió entre 12.2% y 43.4%; y Diplodus argenteus, con una frecuencia de 83.3% y una abundancia relativa mensual entre 16.2% y 31.9%. Estas especies pueden considerarse como características y típicas de la franja arrecifal en Cautaro. Numéricamente, T. bifasciatum fue la especie más abundante desde diciembre de 2000 hasta mayo de 2001, desde julio hasta octubre de 2001 dominó M. chrysurus, mientras que desde octubre a noviembre de 2001 dominó H. aurolineatum. Finalmente, D. argenteus fue la especie dominante de diciembre de 2001 hasta mayo de 2002.
Distribución log-normal de las especies
Tomando en consideración el total del número de individuos observados de cada especie, la distribución log-normal indica una gran relación entre el número de especies en cada octava (st) y los valores teóricos. En este sentido no se observaron diferencias significativas entre ambos (X2 = 13.199; P > 0.05), con una media estimada de 0.69 ± 1.07, donde los máximos números de especies ocurrieron en la primera y tercera octavas (fig. 6).

Análisis trófico de la ictiofauna
Agrupando las especies censadas de acuerdo a la alimentación se observó que las carnívoras presentaron el mayor número de especies (63.1%), seguido de las herbívoras (21.4%) y las omnívoras (15.5%). No obstante, en términos de número total de individuos los carnívoros presentan el mayor valor (41.3%), le siguen los omnívoros (36.67%) y luego los herbívoros (22.03%). Dentro del grupo de las especies carnívoras, se encuentran miembros de las familias Labridae, Chaetodontidae, Haemulidae, Scianidae, Serranidae, Diodonti-dae, Muraenidae y Synodontidae, entre otros. La mayoría de las especies de estas familias (33.96% del total de las carnívoras) se alimentan de presas pequeñas como gasterópodos, pequeños crustáceos, erizos, esponjas, etc., donde se incluyen a Halichoeres bivittatus, Chaetodon capistratus, Lactophrys triqueter y Equetus punctatus. Otro grupo de especies (32.07%) se alimenta de pequeños peces y crustáceos, como Odontoscion dentex, Hypoplectrus puella y Cephalopholis cruentata. Un 18.9% se alimentan exclusivamente de pequeños crustáceos bentónicos como Coryphopterus glaucofraenun y Haemulon steindachneri. Las restantes especies (9.43%) son piscívoras, tales como Gymnothorax moringa, Synodus foetens, Sphyraena barracuda y Mycteroperca rubra.
Los herbívoros estuvieron representados por todas las especies de las familia Scaridae y Acanthuridae, así como Kyphosus sectator, Aluterus scriptus y Abudefduf taurus. En el grupo de los omnívoros se ubicaron principalmente los miembros de la familia Pomacentridae (excepto A. taurus), al igual que Pomacanthus paru, Hypoplectrus unicolor, Diplodus argenteus, Cantherhines pullus, Stephanolepis hispidus y los Tetraodontidae, Canthigaster rostrata y Sphoeroides spen-gleri.
Parámetros fisicoquímicos
La variación de la temperatura superficial del agua presentó el patrón característico de las aguas costeras del nororiente de Venezuela. Desde enero hasta mayo de 2001 la temperatura osciló entre 20°C y 24°C, con el valor más bajo en febrero de 2001. A partir de julio hasta diciembre de 2001 se registró un aumento que alcanzó el valor más alto en agosto 2001 (28°C), registrándose durante el periodo un promedio de 26.10°C. Este ciclo se repitió durante los primeros meses de 2002. La salinidad presentó un patrón de variación menos definido con una escasa fluctuación que osciló entre 35 y 32 (promedio de 33.59960). Las salinidades más altas se observaron en los primeros meses de ambos años.
Discusión
La riqueza de especies observada (86 especies) fue superior a las 51 especies reportadas en esta misma zona por Ruiz et al. (2003) mediante el uso de nasas como arte de pesca, y superior a la señalada por González (1996) y por Rodríguez y Villamizar (2000), quienes encontraron, utilizando igualmente censos visuales, 43 y 68 especies, respectivamente, en zonas arrecifales del Parque Nacional Morrocoy, Venezuela. Asimismo, supera las 46 especies encontradas por Méndez et al. (2001) para una zona protegida (Manzanillo) del Parque Nacional Mochima. Sin embargo, es ligeramente inferior a la estimada por Alvarado (2000) para el Parque Nacional Morrocoy, al occidente de Venezuela (95 especies). Esta diferencia en cuanto a la riqueza de especies registrada por Alvarado (2000) pudiera estar determinada por la mayor área abarcada en su trabajo, que incluye seis zonas arrecifales con características distintas, una de ellas declarada Zona de Protección Integral y que permitiría un mayor resguardo de las especies. El Parque Mochima no posee ningún área bajo esta figura. La riqueza específica observada por Lasso-Alcalá et al. (2003) en zonas arrecifales de la Bahía de Turiamo (104 especies), también al occidente del país, igualmente supera a la señalada en este trabajo quizás debido a la mayor área mues-treada por ellos (ocho estaciones) y al hecho de que esa bahía constituye un área de uso militar especialmente protegida. Igualmente, la diversidad hallada en Cautaro es inferior a la registrada por Adams y Ebersole (2002) (92 especies) para la zona protegida de un arrecife en St. Croix (Islas Vírgenes), pero supera las 72 especies registradas por estos autores en un parche arrecifal evaluado en el mismo trabajo.
La abundancia relativa de las especies en los sistemas arrecifales pareciera variar de acuerdo a factores no claramente definidos. La abundancia en Cautaro de Diplodus argenteus, Microspathodon chrysurus, Thalassoma bifasciatum y Haemulon aurolineatum no coincide con otras áreas estudiadas del mismo parque, como Manzanillo, un sector protegido del oleaje donde las especies más abundantes fueron Scarus iseri, Diplodus argenteus, Chaetodon capistratus y Halichoeres bivittatus (Méndez et al. 2001). En el caso del Parque Nacional Morrocoy, Alvarado (2000) registra como especies más importantes en términos numéricos a Scarus iseri, Stegastes planifrons, Haemulon aurolineatum y Acanthurus bahianus. Tanto Acanthurus bahianus como Stegastes planifrons no se presentan frecuentemente ni en Cautaro ni en Manzanillo. Particularmente resalta la poca abundancia de Scarus iseri en Cautaro, ya que esta especie se ubica entre las más abundantes, no sólo en las áreas ya mencionadas, sino en otros sectores del Parque Morrocoy (Rodríguez y Villamizar 2000) y en el Parque Nacional Los Roques (Posada et al. 2003). Tal observación ha sido previamente planteada por Fariña et al. (2005), quienes encontraron baja importancia numérica relativa de dicha especie en un arrecife del archipiélago Los Monjes. En este sentido, se requiere la ampliación de los estudios en zonas coralinas con diferentes grados de complejidad estructural, determinando un mayor número de variables ambientales y utilizando metodologías comparables, a fin de dilucidar los factores que pudieran estar influenciando la estructura de las comunidades de peces.
La variación estacional del número de especies y del número de organismos evidenció un incremento en los meses de agosto, octubre y noviembre. Estos meses coinciden con la época en la cual bajan en intensidad los vientos alisios del noreste, incrementándose la temperatura, reduciéndose el oleaje y, por lo tanto, aumentando la claridad de las aguas (Gómez 1996). De esta manera, zonas someras como la evaluada adquieren condiciones relativamente ideales para la coexistencia de las especies que pudieran estar influyendo en los valores mencionados.
En el trópico los cambios mareales potenciados por el viento constituyen una de las mayores causas de perturbaciones físicas difíciles de predecir. Sin embargo, se ha observado una rápida colonización de parches de corales afectados en algunas áreas de Australia (Sale 1991) y pocos cambios en la comunidad íctica en zonas coralinas de Guadalupe (Antillas francesas) después de haber sido afectadas por huracanes (Bouchon et al. 1994).
La diversidad íctica observada en la franja arrecifal fue escasamente fluctuante durante todo el periodo de muestreo, a excepción de marzo de 2002, cuando se registró el índice de diversidad más bajo. Tal disminución pudo deberse a la influencia de los vientos alisios del noreste que, para esa fecha, afectaron de manera considerable la zona incrementando el oleaje y la turbidez del agua. Estas características, aunadas a un fenómeno de mar de fondo registrado el día del muestreo, influyeron también en un escaso número de especies (17) y de organismos (102). No obstante, la diversidad (H" ) fue relativamente alta durante todos los muestreos. Esto pudiera relacionarse con el alto porcentaje del área cubierta por rocas y corales (90%) (Sant 1999), ya que esa amplia complejidad topográfica ofrece mayores posibilidades de refugio y recursos para las especies, coincidiendo con lo señalado para el caribe mexicano por Núñez-Lara y Árias-González (1998). En el Caribe se ha informado que numerosas especies de los géneros Acanthurus, Sparisoma y Haemulon, utilizan estas áreas como sitios de desove, alimentación y residencia, las cuales son estacionalmente invadidas por peces predadores de las familias Serranidae, Carangidae y Scorpaenidae (Adams y Ebersole 2002). Sin embargo, la escasez de depredadores grandes de estas familias en el arrecife analizado puede explicarse por la gran actividad pesquera realizada en la zona. La explotación de especies comerciales carnívoras, situadas en el ápice de la cadena alimenticia, pudiera permitir la ocupación de nichos por carnívoros pequeños, incidiendo en una gran diversidad. Friedlander y De Martini (2002) han señalado resultados similares en zonas arrecifales de Hawai.
Diversos investigadores han señalado que la cobertura de coral vivo tiene una influencia positiva en la riqueza de la fauna ictiológica (Bell y Galzin 1984, 1988). Sin embargo, otros no han encontrado esta correlación (Roberts y Osmond 1987, McManus et al. 1981), señalando que una de las razones de estas diferencias puede ser la profundidad a la que se encuentras las colonias de corales. Los resultados obtenidos en esta investigación, sobre la franja arrecifal de escasa profundidad y poca pendiente, coinciden con los señalamientos de Chabanet et al. (1997), quienes indican que las correlaciones entre la riqueza de especies de peces coralinos y la cobertura de coral vivo en áreas someras tienden a ser mayores en los arrecifes planos que en los de pendiente fuerte, lo cual se puede explicar debido a que los peces tienden a permanecer muy cerca del sustrato.
En la estructura trófica de la comunidad íctica de Cautaro se observó una mayor abundancia de especies carnívoras pequeñas. De las 86 especies reportadas, 53 se alimentan de elementos de origen animal. Sin embargo, en relación al número de individuos, de las seis especies con mayores índices de constancia y dominancia, tres son omnívoras (Abudefduf saxatilis, Diplodus argenteus y Microsphatodon chrysurus) y una es herbívora (Sparisoma viride), que fue la especie de Scaridae más abundante (5.54%). Por lo tanto, los herbívoros representan el menor número de individuos en esta comunidad. Al contrario, en un arrecife de Playa Mero (Estado Falcón), donde la cobertura de algas sobre corales muertos representó el 68.8%, predominaron las especies herbívoras de las familias Pomacentridae y Scaridae (75.5%) sobre las carnívoras (16.4%) (Rodríguez y Villamizar 2000). Igualmente, Kurten et al. (2003) hallaron un porcentaje elevado de herbívoros (48.10%) en zonas arrecifales de la Bahía de Turiamo. Por otra parte, Nava-Martínez y Núñez-Lara (2000), para un arrecife de Cozumel (México), también establecieron como nivel trófico dominante el correspondiente a consumidores de plantas y detritus; sin embargo, en relación al número de individuos, predominaron los zooplantófagos.
Los resultados obtenidos en el presente trabajo parecieran sugerir una presión de herbivoría baja por los peces sobre las algas en la zona de estudio. A pesar de ello no se observó una cobertura de macroalgas significativa y el número de especies (23) es menor que en otros arrecifes del área (Barrios et al. 2003). En el caso de Cautaro quizás el crecimiento de algas esté más controlado por equinodermos, moluscos u otros herbívoros que por los peces. Estudios posteriores que abarquen tales taxa pudieran ayudar a dilucidar las relaciones tróficas de toda la comunidad.
Por otra parte, Hobson (1991) plantea que el mayor componente en una comunidad coralina es el de los zooplanctófagos. En el arrecife estudiado este grupo representó igualmente un porcentaje alto (33.9% de las especies carnívoras y un 43.7% del total de individuos carnívoros). A este respecto, Sierra et al. (1994) señalan que en los arrecifes grandes de Cuba predominan los peces herbívoros sobre los omnívoros en una proporción de 9:1. Sin embargo, en arrecifes aislados y de menor tamaño la proporción de carnívoros es alrededor de dos veces mayor que la de los omnívoros y herbívoros. La estructura trófica en el arrecife estudiado, en la cual los carnívoros y los omnívoros son los grupos más abundantes, puede deberse a la elevada productividad de las aguas de las costas orientales de Venezuela, la cual entre otras razones es producto de la surgencia costera (Castellanos et al. 2002) que eleva la disponibilidad de nutrientes a nivel planctónico.
Agradecimientos
Se agradece al Consejo de Investigación de la Universidad de Oriente por el financiamiento del Proyecto CI-5-1001-0887/ 99 denominado "Caracterización bioecológica de comunidades asociadas a ecosistemas coralinos en la Bahía de Mochima y Golfo de Santa Fe, Edo. Sucre, Venezuela".
Referencias
Adams A, Ebersole J. 2002. Use of back-reef fish communities. Mar. Ecol. Prog. Ser. 288: 213-226. [ Links ]
Alvarado D. 2000. Variabilidad espacial de la estructura de la comunidad de peces de arrecifes del Parque Nacional Morrocoy. Trabajo especial de grado, Universidad Simón Bolívar, Venezuela, 81 pp. [ Links ]
Antonius A. 1980. Occurrence and distribution of stony corals in the Gulf of Cariaco, Venezuela. Int. Rev. Hydrobiol. 65: 321-338. [ Links ]
Barrios J, Sant S, Méndez E, Ruiz L. 2003. Macroalgas asociadas a arrecifes coralinos en el Parque Nacional Mochima, Venezuela. Saber 15: 28-38. [ Links ]
Bell JD, Galzin R. 1984. Influence of live coral cover on coral reef fish communities. Mar. Ecol. Prog. Ser. 15: 265-274. [ Links ]
Bell JD, Galzin R. 1988. Distribution of coral and fish in the lagoon at Mataiva: Potential for increase through mining? Proc. 6th Int. Coral Reef Symp. 2: 347-352. [ Links ]
Bouchon C, Bouchon-Navaro Y, Louis M. 1994. Changes in the coastal fish communities following Hurricane Hugo in Guadeloupe Island (French West Indies). Atoll Res. Bull. 422: 113. [ Links ]
Castellanos P, Varela P, Muller-Karger F. 2002. Descripción de las áreas de surgencia al sur del Mar Caribe examinadas con el sensor infrarrojo AVHRR. Mem. Fund. La Salle Cienc. Nat. 154: 55-76. [ Links ]
Cervigón F. 1994. Los Peces Marinos de Venezuela. Vol. III. Ed. Ex Libris, Caracas, 294 pp. [ Links ]
Cervigón F. 1996. Los Peces Marinos de Venezuela. Vol. IV. Ed. Ex Libris, Caracas, 254 pp. [ Links ]
Chabanet P, Ralambondrainy H, Amanieu M, Faure G, Galzin R. 1997. Relationships between coral reef substrata and fish. Coral Reefs 16: 93-102. [ Links ]
Fariña A, Bellorin A, Sant S, Méndez E. 2005. Estructura de la comunidad de peces en un arrecife del Archipiélago Los Monjes, Venezuela. Cienc. Mar. 31: 1-7. [ Links ]
Friedlander A, De Martini E. (2002). Contrast in density, size and biomass on reef fishes between the northwestern and the main Hawaiian Islands: Effects of the fishing down apex predator. Mar. Ecol. Prog. Ser. 230: 253-264. [ Links ]
Gauthier E. 2003. Estructura trófica de la comunidad íctica de un parche arrecifal del Parque Nacional Mochima, Edo. Sucre, Venezuela. Tesis de maestría. Instituto Oceanógrafico de Venezuela, Universidad de Oriente, Cumaná, Venezuela, 58 pp. [ Links ]
Glynn P. (1976). Some physical and biological determinants of coral community structure in the eastern Pacific. Ecol. Monogr. 46: 431-456. [ Links ]
Gómez A. 1996. Causas de la fertilidad marina en el nororiente de Venezuela. Interciencia 21: 140-146. [ Links ]
González A. 1996. Peces depredadores de corales pétreos en un arrecife del Parque Nacional Morrocoy, Edo. Falcón. Trabajo especial de grado, Universidad Central de Venezuela, Escuela de Biología, Caracas, Venezuela, 68 pp. [ Links ]
Hobson E. 1991. Trophic relationships of fishes specialized to feed on zooplankters above coral reefs. In: PF Sale (ed.), The Ecology of Fishes on Coral Reefs. Academic Press, San Diego, California, pp.69-95. [ Links ]
Humann P. 1997. Reef Fish Identification. 2nd ed. New Word Publication, Florida, 398 pp. [ Links ]
Krebs C. 1989. Ecology: The Experimental Analysis of Distribution and Abundance. Harper & Row, New York, 694 pp. [ Links ]
Kurten M, Klein E, Alvarado D. 2003. Estructura de la comunidad de peces arrecifales del Parque Nacional Morrocoy y la Bahía de Turismo. V Congreso Venezolano de Ecología, Isla de Margarita, Venezuela. Resumen, p. 99. [ Links ]
Lasso-Alcalá O, Klein E, Kurten M, Villamizar E. 2003. Diversidad de peces arrecifales de la Bahía de Turismo, litoral centro- occidental, Venezuela. V Congreso Venezolano de Ecología, Isla de Margarita, Venezuela. Resumen, p. 101. [ Links ]
McManus J, Miclat R, Palaganas V. 1981. Coral and fish community structure of Sombrero Island at Bataganas, Philippines. Proc 4th Int. Coral Reef Symp. 2: 271-280. [ Links ]
Martínez-Orsatti L, Méndez de E E, Ruiz L, Torres de J A, Rivas A. 2001. Hábitos alimentarios de juveniles de tres especies de Haemulon (Pisces: Haemulidae) en praderas de Thalassia testudinum de la Bahía de Mochima, Estado Sucre, Venezuela. Ciencia 9: 28-39. [ Links ]
Méndez de E E, Ruiz L, Torres A, Sant S, Barrios J, Marín B, Prieto A. 2001. Comunidad íctica de dos parches arrecifales del Parque Nacional Mochima, Estado Sucre, Venezuela. Trigésima Reunión de la Asociación de Laboratorios Marinos del Caribe (ALMC), La Parguera, Puerto Rico. Resumen, p. 63. [ Links ]
Méndez de E E, Ruiz L, Torres A, Rivas A, Martínez L. 2003. Hábitos alimentarios de Coryphopterus glaucofraenum (Pisces: Gobiidae) en la Bahía de Mochima, Estado Sucre, Venezuela. Ciencia 11: 31-38. [ Links ]
Nava-Martínez G, Núñez-Lara E. 2000. Estructura de la comunidad de peces en arrecifes de la isla Cozumel. V Congreso de Ciencias del Mar, La Habana, Cuba. Programa Científico. 130 pp. [ Links ]
Núñez-Lara E, Arias-González E. 1998. The relationship between reef fish community structure and environmental variables in the southern Mexican Caribbean. J. Fish Biol. 53: 209-221. [ Links ]
Pauls S. 1982. Estructura de comunidades coralinas de la Bahía de Mochima, Venezuela. Tesis de maestría, Instituto Oceanográfico de Venezuela, Universidad de Oriente, Cumaná, Venezuela, 117pp. [ Links ]
Pielou EC. 1975. Ecological Diversity. J. Wiley, New York, 670 pp. [ Links ]
Posada JM, Villamizar E, Alvarado D. 2003. Rapid assessment of coral reefs in the Archipiélago de Los Roques National Park, Venezuela (Part 2: Fishes). In: Lang JC (ed.), Status of Coral Reefs in the Western Atlantic: Results of Initial Surveys, Atlantic and Gulf Rapid Reef Assessment (AGRRA) Program. Atoll Res. Bull. 496: 531-544. [ Links ]
Preston FN. 1948. The commonness and rarity of species. Ecology 29: 254-283. [ Links ]
Randall JE. 1967. Food habits of reef fishes of the West Indies. Proc. International Conference on Tropical Oceanography. Stud. Trop. Oceanogr. 5: 665-847. [ Links ]
Roberts CM, Osmond RF. 1987. Habitat complexity and coral reef diversity and abundance on Red Sea fringing reefs. Mar. Ecol. Prog. Ser. 41: 1-8. [ Links ]
Rodríguez J, Villamizar E. 2000. Estructura de la comunidad de peces arrecifales de Playa Mero, Parque Nacional Morrocoy, Venezuela. Rev. Biol. Trop. 48: 107-113. [ Links ]
Ruiz LJ, Méndez E, Torres A, Prieto A, Marín B, Fariña A. 2003. Composición, abundancia y diversidad de peces arrecifales en dos localidades del Parque Nacional Mochima, Venezuela. Cienc. Mar. 29: 185-195. [ Links ]
Sale PF. 1991. Reef fish communities: Open nonequilibrial systems. In: Sale P (ed.), The Ecology of Fishes on Coral Reefs. Academic, San Diego, pp. 564-598. [ Links ]
Sant S. 1999. Ecología de comunidades coralinas del Edo. Sucre, Venezuela. Tesis de maestría, Instituto Oceanográfico de Venezuela, Universidad de Oriente, Cumaná, Venezuela, 117 pp. [ Links ]
Sant S, Prieto A, Méndez E. 2003. Cambios en la composición y estructura de una comunidad coralina después de un fenómeno de mar de fondo en Cautaro, Parque Nacional Mochima, estado Sucre, Venezuela. Ciencia 12: 5-12. [ Links ]
Sierra L, Claro RJ, Posova O. 1994. Alimentación y relaciones tróficas. Capítulo V. En: Claro R (ed.), Ecología de los Peces Marinos de Cuba. Ediciones del Centro de Investigaciones de Quintana Roo, México, pp. 263-319. [ Links ]
Taylor LR, Kempton R, Wolwood A. 1976. Diversity, statistics and the log-series model. J. Anim. Ecol. 45: 337-365. [ Links ]

