










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkCiencias marinas
Print version ISSN 0185-3880
Cienc. mar vol.31 n.2 Ensenada Jun. 2005
Artículos
Crecimiento y mortalidad de Pinctada imbricata (Mollusca: Pteridae) en Guamachito, Península de Araya, Estado Sucre, Venezuela
Growth and mortality of Pinctada imbricata (Mollusca: Pteridae) in Guamachito, Araya Peninsula, Sucre State, Venezuela
Jesús S. Marcano1*, Antulio Prieto2, Asdrúbal Lárez1, José J. Alió1 y Hargadys Sanabria1
1 Instituto Nacional de Investigaciones Agrícolas (INIA) Ministerio de Ciencia y Tecnología Apartado 236 Cumaná, Estado Sucre 6101, Venezuela. * E-mail: jmarcano@inia.gov.ve
2 Departamento de Biología Escuela de Ciencias Núcleo de Sucre Universidad de Oriente Cumaná, Venezuela. E-mail: plica4@yahoo.com
Recibido en junio de 2004;
aceptado en diciembre de 2004.
Resumen
Se analizó el crecimiento, la distribución de tallas y la mortalidad de una población de Pinctada imbricata en Guamachito, Península de Araya, Venezuela, entre enero y diciembre de 2000. Los muestreos se realizaron mensualmente sobre transeptos perpendiculares a la línea de la costa con una rastra artesanal, tomando también datos sobre temperatura, oxígeno disuelto y salinidad de la columna de agua. La distribución de tallas fue variable con un predominio de tallas entre 50 y 70 mm, que constituyeron el 75% de la población. La mayor cantidad de juveniles fue observada en marzo, cuando la longitud media en la población fue de 55.8 mm. Los promedios de los pesos húmedo y seco siguieron la misma tendencia, con valores mínimos en junio, julio, septiembre y octubre de 2000. Las relaciones longitud-peso húmedo y longitud-peso seco fueron altamente significativas (P < 0.05). Los parámetros de la ecuación de crecimiento estacionalizada de von Bertalanffy fueron L∞ = 85.15 mm; K = 1.42 año-1, ts = 0.2, y C = 0.20; indicando un crecimiento lento (WP = 0.42) en el periodo de julio a noviembre, asociado con los parámetros ambientales del área con una longevidad estimada en 3 años. La mortalidad total fue de 14.35 año para los individuos pequeños (10-36 mm) y 3.45 año, para los individuos grandes (36-84 mm). La alta tasa de crecimiento de P. imbricata muestra ser una ventaja para considerarla como una especie con potencial para actividades de acuicultura en la región. El banco ostrícola presenta buenas condiciones para mantener una explotación pesquera sostenida, tomando en consideración parámetros pesqueros como la captura registrada, el esfuerzo óptimo, la talla mínima de extracción, la mortalidad por pesca y la mortalidad natural.
Palabras clave: bivalvo, ostra perla, Pinctada imbricata, crecimiento, mortalidad, Venezuela.
Abstract
Growth, size distribution and mortality of Pinctada imbricata from Guamachito, Araya Peninsula, Venezuela, were analyzed between January and December 2000. Monthly samples were taken on perpendicular transepts to the coastline with an artisanal dredge, and data on sea surface temperature, dissolved oxygen and salinity were registered. The size distribution was variable, with a predominance of size between 50 and 70 mm in length, which represented 75% of the population. The greatest amount of juveniles was observed in March, when the mean length of the population was 55.8 mm. The variation in mean wet and dry weights presented a similar trend, with minimum values in June, July, September and October. The length/wet weight and length/ dry weight relationships were highly significant (P < 0.05). The parameters of the seasonal von Bertalanffy growth equation were L∞ = 85.15 mm, K = 1.42 yr 1, ts = 0.2 and C = 0.20, indicating a slow growth in the period from July to November (WP = 0.42) that could be associated with the environmental conditions of the area. Longevity was estimated to be three years. Total mortality rate (Z) was 3.45 per year for small individuals (10-36 mm) and 14.36 per year for the larger individuals (36-84 mm). The high growth rate of P. imbricata supports its use for aquacultural purposes in the region. The Guamachito oyster bank shows good conditions to support sustained fishery exploitation, considering fishery parameters such as registered catch, optimal effort, minimum extraction size, fishing mortality and natural mortality.
Key words: bivalve, pearl oyster, Pinctada imbricata, growth, mortality, Venezuela.
Introducción
La ostra perla Pinctada imbricata L. es un bivalvo de la familia Pteridae ampliamente distribuida en el Océano Atlántico desde Carolina del Norte, EUA, hasta Brasil en América del Sur. Se desarrolla en áreas poco profundas generalmente en zonas de corales, conchas y praderas de Thalassia testudinum en profundidades de 5 a 30 m, con aguas relativamente claras (Abbott, 1974; Lodeiros et al., 1999).
La especie fue muy importante en el Caribe subtropical debido al gran interés que históricamente despertó la producción de perlas (MacKenzie et al., 2004). Aunque otras especies del mismo genero, como P. margaritifera en la Polinesia francesa y P. fucata en Australia y Japón desempeñan actualmente un papel económico de primer orden por la gran producción de perlas, en el Mar Caribe esta actividad ha perdido el auge de épocas pasadas y en la actualidad se utiliza la carne de las ostras para el consumo en fresco en los bancos naturales del oriente de Venezuela (Novoa et al., 1998; Marcano, 2001). La explotación de las ostras está regulada por la Resolución INAPESCA No. 009 del 12 de agosto de 2002, la cual establece un periodo de extracción del 15 de marzo al 30 de julio. Recientemente se han desarrollado estudios destinados a examinar la factibilidad de su cultivo potencial, tanto en poblaciones naturales como bajo condiciones controladas (Urban, 2000; Lodeiros, et al., 2002; Lodeiros y García, 2004).
Esta especie es muy abundante en las costas nororientales de América del Sur, formando densos bancos en el Mar Caribe, tanto en Colombia como en Venezuela. Existen pocas investigaciones sobre P. imbricata en comparación con otras especies del mismo género. La mayoría de los estudios realizados en Venezuela han estado orientados hacia aspectos reproductivos (Marcano, 1984; Ruffini, 1984), índice de engorda (León y Millán, 1996), evaluación del recurso (Salaya y Salazar, 1972), producción somática en una población natural (Verginelli y Prieto 1991) y estudios ecotoxicológicos (Lodeiros, 1999; Nusetti et al., 2004).
El objetivo de este estudio fue analizar el crecimiento, en talla, de una población de P. imbricata situada en el banco ostrícola de Guamachito, costa norte de la Península de Araya, Estado Sucre, y establecer un modelo de crecimiento estacional de la especie.
Materiales y métodos
Las muestras de P imbricata fueron recolectadas de enero a diciembre de 2000, en Guamachito, localidad situada en la costa norte de la Península de Araya, Estado Sucre, Venezuela (10°38'N, 64°19'W y 10°39'N, 64°5'W). El sustrato del área es areno-fangoso, con restos de piedra y cubierto parcialmente con T. testudinum. Los vientos alisios del noroeste caracterizan la zona costera, la cual presenta extrema aridez y muy poca vegetación.
Para la extracción de las muestras se empleó una rastra artesanal de 1 m de pala por 0.80 m de saco. Se establecieron seis transectos en el área de distribución de P. imbricata, perpendiculares a la costa y separadas 500 m entre sí. De cada uno se extrajeron tres réplicas con la rastra, trabajando a profundidades de 2 a 12 m. A los ejemplares recolectados se les determinó la longitud máxima, Lt (medida desde la charnela hasta el borde inferior), con un vernier de precisión 0.05 mm; el peso húmedo total de la carne (Pt en gramos), fue medido en una balanza con precisión 0.0001 g; mientras que el peso seco de las partes blandas (Ps en gramos) se determinó secando el tejido a 70°C hasta peso constante, de acuerdo a lo señalado por Crisp (1971).
Las relaciones entre los pesos (Ph y Ps) y la longitud (Lt) se determinaron por la ecuación P = a Ltb ajustada por el método de los mínimos cuadrados (Wilburg y Owen, 1964), y la significancia estadística de la correlación se estimó utilizando el método de Hotelling (Sokal y Rohlf, 1995).
La composición de tallas de la fracción de la población muestreada mensualmente se analizó mediante el uso de histogramas de frecuencias, elaborados con intervalos de clase de 2 mm. También se evaluó la variación del peso húmedo (Ph) durante el periodo de muestreo.
Los parámetros de crecimiento, L∞ (longitud máxima teórica) y K (coeficiente de crecimiento), se estimaron utilizando la forma estacional de crecimiento de von Bertalanffy. Con este propósito se utilizó, en primer lugar, el método de Battacharya (1967), para descomponer gráficamente las distribuciones polimodales de frecuencias, suponiendo normalidad en cada componente. Luego, para obtener estimados preliminares de L∞, K y los valores de oscilación estacional (C y WP), se utilizaron los métodos de Gulland y Holt (1959), Fabens (1965) y Munro y Pauly (1983). Finalmente, se aplicó el análisis de superficie de respuesta (Rn) a fin de conocer la forma estacional del modelo según Pauly y Gaschutz (1979) y Gayanilo et al., (1989), el cual quedó representado por la ecuación:

donde C es la amplitud de oscilación del crecimiento, to es el tiempo a partir del cual comienza a crecer el bivalvo, y ts es el periodo del año cuando el crecimiento es máximo. Estos métodos, basados en el análisis de datos de frecuencia de longitud, forman parte del paquete computarizado FISAT (Gayanillo and Pauly, 1997).
La mortalidad total de la población se estimó utilizando el método de la curva de captura linearizada, con tallas convertidas a edades (Pauly, 1983), según la expresión loge (N/Δt) = a-bt, donde -b = Z es la tasa instantánea de mortalidad total.
En cada muestreo se determinaron, la temperatura superficial (°C), la salinidad (‰) y el oxígeno disuelto del agua (mg L-1), de acuerdo con los métodos descritos por Strickland y Parsons (1972).
Resultados
La estimación de los parámetros de la ecuación de crecimiento de P. imbricata en el banco de Guamachito sugiere que la especie es de rápido crecimiento con una longevidad máxima teórica de 3 años. Los ajustes obtenidos por los métodos de Gulland y Holt (1959) y Fabens (1965), fueron muy semejantes, con valores de 85.5 mm y 87.27 mm respectivamente, difiriendo de los 72.18 mm obtenidos por Munro y Pauly (1983). Así, los mejores ajustes para la ecuación del crecimiento con estacionalidad fueron: L∞ = 85,15 mm, K = 1.42 año-1; C = 0,20; WP = 0.42; ts= 0.2 y Rn = 1.39. La expresión final del modelo estacional (fig. 1) está representada por la ecuación:
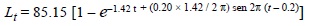

Se compararon diferentes curvas de crecimiento de P. imbricata estimadas en otros estudios y en diferentes localidades, utilizando el parámetro (Φ') Phi prima (Munro y Pauly, 1983; Pauly y Munro, 1984). Esta prueba sugiere que el desarrollo del crecimiento está reflejado por (Φ') = log10 K + 2log10 L∞ y los resultados indican que los valores de Φ' son muy similares (tabla 1).
La estructura de frecuencias mensuales de tallas en la población fue muy variable durante 2000 (fig. 2), observándose siempre un predominio de bivalvos con longitudes en el intervalo de 50 a 70 mm, los cuales constituyeron el 77% de la población. La proporción de individuos juveniles fue baja, detectándose la mayor cantidad en marzo de 2000. El promedio de talla para el banco durante el año fue de 55.8 mm.

Los promedios de peso seco y húmedo de la carne presentaron valores altos en los periodos febrero-mayo, agosto y octubre-noviembre, mientras que los mínimos se observaron en junio-julio, septiembre y diciembre (fig. 3), detectándose diferencias significativas entre los promedios mensuales (P < 0.05). La ecuación que relacionó el peso total con la longitud de la concha estuvo representada por la ecuación Pt = 0.00007 Lt3.15 (r2 = 0.84; n = 1352).
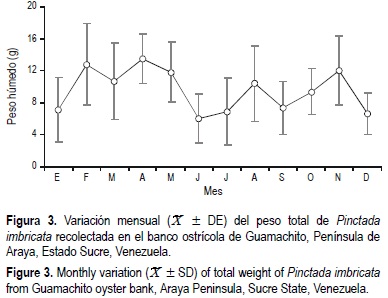
La variación mensual del peso seco siguió la misma tendencia del peso total, con promedios mínimos en junio, julio, septiembre y diciembre de 2000. La relación entre el peso seco y la longitud estuvo representada por la ecuación Ps = 0.00001 Lt3.71 (r2 = 0.90, n = 300). No se detectaron diferencias significativas entre los coeficientes de regresión en las relaciones entre longitud total de la concha y peso húmedo total y peso seco, por ello se utilizó una ecuación común en ambos casos.
Al analizar la abundancia total relativa de la distribución de tallas en la población se pueden distinguir dos grupos que dominan numéricamente, uno de 10 a 36 mm de Lt y otro de 38 a 88 mm (fig. 4). Al estimar la mortalidad total en los dos grupos se determinó que los individuos con tallas entre 10 y 36 mm presentaron una tasa de mortalidad Z = de 14,35, que es equivalente a una mortalidad anual del 99%, mientras que en los individuos más grandes (en el intervalo de 36 a 84 mm) se obtuvo un valor de Z = 3.96, equivalente a una mortalidad anual del 96 % (fig. 5).


La temperatura del agua varió entre 24.8°C (abril de 2000) y 28.5°C (septiembre de 2000), con una media de 25°C ± 0.12 (fig. 6). La salinidad tuvo escasa variación, con valores entre 36.2% (mayo de 2000) y 37.3% (enero de 2000). La concentración del oxígeno disuelto presentó un patrón irregular, con valores bajos de noviembre a diciembre de 2000 y valores altos de abril a julio de 2000 (fig. 6).

Discusión
La amplitud de la oscilación del crecimiento de P. imbricata (indicada por C = 0.20) estaría asociada principalmente con las variaciones de los parámetros ambientales en el área durante las estaciones de lluvia y sequía, ya que la temperatura presenta una variación estacional de 4-5°C y la salinidad varía en hasta 2%. El punto de invierno (WP = 0.42) indica que las diferencias en las tasas de crecimiento de los organismos fueron significativas, teniendo lugar el crecimiento más lento en talla durante el periodo entre julio y noviembre, con un punto medio en agosto; mientras que ts, que representa el crecimiento máximo del año, fue 0.2 y se observó en el primer semestre con un punto medio en febrero. Estas variaciones en las tasas de crecimiento en longitud de este bivalvo, se explican por las condiciones ambientales del área. Así, de enero a junio en la zona tienen lugar surgencias costeras caracterizadas por bajas temperaturas del agua y grandes contenidos de nutrientes y biomasa fitoplanctónica, originando un periodo de elevada productividad primaria. En contraste, de julio a noviembre la zona presenta un periodo de estratificación de las capas superficiales del mar, con una contracción de la zona productiva e incrementos de temperatura y baja disponibilidad del fitoplancton (Griffiths y Simpson, 1972; Ferraz-Reyes, 1989). No se analizó si la disminución del crecimiento en este periodo está relacionada con la transferencia de energía hacia otros tejidos, especialmente las gónadas, pero se ha informado que en Lyropecten nodosus y Pinna carnea sometidas a cultivo suspendido, ocurre una disminución del crecimiento del tejido muscular durante el período de formación de gametos (Narváez et al., 2000; Mendoza et al., 2001).
El valor de L∞ (85.15 mm) obtenido en la localidad de Guamachito para P. imbricata fue ligeramente superior al reportado por Urban (2000) en una población de la Guajira colombiana (L∞ = 84.00), y muy superior al estimada por Verginelli y Prieto (1991) en una población del Golfo de Cariaco (L∞ = 74.69), en la cual las longitudes máximas no sobrepasaron los 76 mm. El valor de la tasa de crecimiento en la población estudiada (K = 1.41 año-1) fue muy superior a las informadas previamente. Esto indica que Guamachito constituye una zona con condiciones excelentes para el desarrollo del bivalvo, el cual puede alcanzar una longitud de 61 mm un año después de su fijación. A pesar de esto, el ritmo de crecimiento de P imbricata es menor que el del bivalvo Arca zebra, arcido de gran importancia comercial en el oriente de Venezuela que comparte la misma área geográfica y alcanza una talla de 58 mm a los seis meses después de su fijación. Probablemente la mayor producción de materia orgánica de A. zebra, y en consecuencia su más alto cociente producción/biomasa (P/B), así como la excesiva e irracional explotación de los bancos de la ostra perla, sean las principales razones (entre otras) del desplazamiento de P. imbricata por A. zebra, de sus antiguos bancos, como lo han sugerido Salaya y Salazar (1972) y Lárez y Marcano (1982).
Los valores de Φ calculados para la especie a partir de los parámetros de crecimiento estimados en trabajos previos y en este estudio, están distribuidos normalmente. Munro y Pauly (1983), así como Pauly y Munro (1984), encontraron que las especies en una misma familia, tienen valores similares de Φ y calcularon este parámetro para gran número de especies encontrando que, dentro de una familia, los valores de Φ están normalmente distribuidos.
El patrón de reclutamiento de juveniles encontrado en Guamachito, concentrado en marzo, coincide parcialmente con el registrado por Verginelli y Prieto (1991) en una población explotada de Pariche, Golfo de Cariaco, Estado Sucre. Sin embargo, estos autores también recolectaron juveniles en otros meses del periodo de muestreo (marzo de 1985 a marzo de 1986), utilizando un método basado en buceo autónomo y cuadrantes de 0.0625 m2. Las diferencias en la estructura de tallas de ambas poblaciones pueden deberse a cualquiera de las siguientes causas: (a) la utilización de diferentes métodos de muestreo; (b) la existencia de estrategias variables de desove y fijación de reclutas en la especie, dependientes de las condiciones hidroquímicas e hidrográficas en cada localidad; y (c) muy relacionada con la anterior, la complicada sexualidad de las especies del género Pinctada, que maduran por primera vez como machos y luego cambian a hembra, exhibiendo un comportamiento protándrico que en algunos casos muestra variaciones impredecibles. El proceso de cambio de sexo puede estar potencialmente relacionado con la disponibilidad de alimento en el ambiente (Tranter, 1958; Rose et al., 1990; Saucedo y Monteforte, 1997; Pouvreau et al., 2000).
La distribución general de frecuencias de tallas fue bimodal, lo que indica que los reclutas se agregan estacionalmente a la población. Esto contrasta con la distribución unimodal reportada para una población de la Guajira colombiana (Urban, 2000). La presencia de distribuciones de tallas multimodales puede considerarse normal para una especie con una estrategia de reproducción sincrónica y varios desoves anuales provenientes de individuos con diferentes edades, como se ha informado para P margaritifera de la Polinesia francesa (Pouvreau et al., 2000).
El promedio de talla encontrado en el banco ostrícola de Guamachito para P. imbricata (55.8 mm), es similar al reportado por Marcano (2001), para organismos de la Cabecera de la Isla Cubagua, Estado Nueva Esparta, Venezuela (55.32 mm). En consistencia con este criterio, la talla mínima comercial de extracción de 50 mm es adecuada, considerando además que la porción explotable del total de las ostras es la constituida por animales de talla superior a la sugerida, y que en este banco esa porción constituye 79.6% de la población. En el Mar Caribe colombiano, Borrero et al. (1996) señalan para esta especie una talla mínima comercial de 50 mm Lt.
Las tendencias observadas en los promedios de peso seco coinciden con la informada para la misma especie en Pariche (Golfo de Cariaco). La variación del peso seco en un animal estándar de 40 mm Lt, evaluada por ecuaciones mensuales de regresión Ps-Lt, presentó disminuciones de peso de julio a noviembre (Verginelli y Prieto, 1991). Estas variaciones probablemente se relacionen con la fase de post desove o reposo sexual durante la cual se reabsorbe el tejido gonadal. No existen en la zona estudios histológicos sobre la gametogénesis de P. imbricata. Sin embargo, investigaciones realizadas en la Guajira colombiana han señalado que la baja condición fisiológica observada de julio a diciembre, representada por bajos pesos secos, se debe a la escasez de nutrientes y fitoplancton ocasionada por la reducción de las surgencias costeras en tal período, sugiriendo que el ciclo gametogénico de la especie depende más de la disponibilidad de alimento que de la temperatura (Verginelli y Prieto, 1991).
La mortalidad observada a partir de la curva de captura indica la existencia de dos tendencias en la población: en los individuos pequeños, con intervalo de tallas de 10 a 36 mm, la mortalidad total observada (Z = 14.35) fue mayor que en los individuos grandes en el intervalo de 37 a 88 mm (Z = 3.45). Este patrón también ha sido reportado en Donax dentifer (Riascos y Urban, 2002) y, dado que la población está sometida a explotación comercial, se supone que estos valores de mortalidad total incluyen tanto la mortalidad natural como la de pesca.
Generalmente se supone que los individuos pequeños de ostra perla son más vulnerables a la depredación que los grandes, debido a la fragilidad de su concha que los hace presa fácil de distintos grupos de invertebrados como crustáceos y gasterópodos perforadores. Urban (2000) documentaron en el Mar Caribe colombiano la depredación en colectores y sistemas de cultivo experimentales de ostra perla, por gasterópodos del género Cymatium spp. (Familia Ranellidae): C. pileare, C. cingulatum y C. nicobaricum, así como por varias especies de crustáceos pertenecientes a las familias Portunidae, Xanthidae, Magidae y Palinuridae. En sistemas de cultivos del Golfo de Cariaco, Venezuela, también se ha señalado la presencia de gasterópodos del género Cymatium, los cuales entran a las cestas como larvas, y crecen rápidamente para consumir ostras juveniles (Lodeiros et al., 2002).
No se determinó la curva de crecimiento en peso debido a la variabilidad observada por los continuos periodos de desove. Éstos hacen difícil el acoplamiento entre el crecimiento de la concha y el de los tejidos, situación característica de bivalvos tropicales (Rajagopal et al., 2002).
Los estudios sobre dinámica poblacional de P. imbricata revisten una complejidad característica debido a la cercanía geográfica de los diferentes bancos que se localizan en el oriente de Venezuela. Por ello, para entender correctamente la dinámica de las diferentes poblaciones de la especie es necesario relacionar este proceso con el patrón de historia de vida del bivalvo en cada uno de ellos.
Los resultados obtenidos en la presente investigación indican que P. imbricata presenta una tasa de crecimiento alta, alcanzando el tamaño comercial antes de un año después de su fijación. La mayor cantidad de juveniles se observó en marzo de 2000, aunque la tasa de reclutamiento fue baja. La talla mínima sugerida para la extracción es de 50 mm. Estas características indican que la especie en el área presenta excelentes condiciones para soportar una explotación sostenida, tomando en consideración otros parámetros pesqueros tales como la captura y el esfuerzo óptimo, la talla mínima de extracción, la mortalidad por pesca y la mortalidad natural. También puede ser utilizada en programas de cultivos controlados que contemplen medidas de protección contra depredadores para disminuir la alta mortalidad de juveniles. Debido al interés estratégico alimentario de este recurso, reservado de manera exclusiva a los pescadores artesanales, se plantea la necesidad de iniciar estudios dirigidos a la determinación de parámetros biológico-pesqueros que conlleven a la administración racional de este recurso en el oriente de Venezuela.
Agradecimientos
La presente investigación fue parcialmente financiada por el Instituto Nacional de Investigaciones Agropecuarias (INIA) y por el Consejo de Investigación de la Universidad de Oriente (Proyecto CI-5-1001-1094/02).
Referencias
Abbott, R.T. (1974). American Seashells. 2nd ed., D. van Nostrand Reinhold Co., New York, pp. 1-663. [ Links ]
Battacharya, C. (1967). A simple method of resolution of a distribution into Gaussian components. Biometrics, 23: 115-135. [ Links ]
Borrero, J., Díaz, J. y Seczon, A. (1996). Las ostras perlíferas (Bivalvia, Pteridae) en el Caribe colombiano. Inst. Inv. Mar y Costeras (INVEMAR). Serie de Publicaciones Especiales, No. 1, 53 pp. [ Links ]
Crisp, D.J. (1971). Energy flow measurements. Chapter 12. In: Methods for the Study of Marine Benthos. IBP Handbook No. 16, Blackwell Scientific Publications, Oxford and Edinburgh, 334 pp. [ Links ]
Fabens, A.J. (1965). Properties and fitting of the von Bertalanffy growth curve. Growth, 29: 265-285. [ Links ]
Ferraz-Reyes, E. (1989). Influencia de los factores físicos en la distribución vertical de la biomasa fitoplanctónica en el Golfo de Cariaco (Venezuela). Bol. Inst. Oceanogr. Venezuela, 28: 47-56. [ Links ]
Gayanilo, F. and Pauly, D. (1997). FAO-ICLARM Stock Assessment Tools (FISAT) Reference Manual. FAO Comp. Info. Ser. (Fisheries), 8, 262 pp. [ Links ]
Gayanilo, F., Soriano, M. and Pauly, D. (1989). A draft guide to the compleat ELEFAN. ICLARM Software No. 265. [ Links ]
Griffiths, J. y Simpson, J.G. (1972). Afloramientos y otras características oceanográficas de las aguas costeras del noroeste de Venezuela. Serie Recursos y Explotación Pesquera, MAC/ PNUD/FAO Caracas, 2: 1-72. [ Links ]
Gulland, J.A. and Holt, S.J. (1959). Estimation of growth parameter for data at unequal time intervals. J. Cons. CIEM, 25: 47-49. [ Links ]
Lárez, A. y J. Marcano (1982). Informe sobre la explotación de los bancos ostrícolas en Nueva Esparta y Sucre. Informe Técnico MAC, 18 pp. [ Links ]
León, L. y Millán, Q. (1996). Variación del índice de condición y de la condición bromatológica de la ostra perlífera Pinctada imbricata Röding 1798 (Mollusca: Bivalvia) en Las Cabeceras, Isla de Cubagua, Venezuela. Mem. Soc. Cien. La Salle, 146: 23-41. [ Links ]
Lodeiros, C. (1999). Selección de especies de bivalvos representantes de ambientes marino-costeros con potencialidades para su empleo en evaluaciones de toxicidad. Informe Servicio INTEVEP S.A., 67 pp. [ Links ]
Lodeiros, C. and García, N. (2004). The use of sea urchins to control fouling during suspended culture of bivalves. Aquaculture, 231: 293-298. [ Links ]
Lodeiros, C., Marín, B. y Prieto, A. (1999). Catálogo de Moluscos del Nororiente de Venezuela: Clase Bivalvia. Edición APUDONS, Cumaná, Venezuela, 9 placas, pp. 1-109. [ Links ]
Lodeiros, C., Pico, D., Prieto, A., Narváez, N. and Guerra, A. (2002). Growth and survival of the pearl oyster Pinctada imbricata (Roding 1768) in suspended and bottom culture in the Golfo de Cariaco, Venezuela. Aquacult. Int., 10: 327-338. [ Links ]
MacKenzie C., Trocooli, L. and Leon L. (2004). History of the Atlantic pearl-oyster Pinctada imbricata industry in Venezuela and Colombia, with biological and ecological observations. Mar. Fish Rev., 65(1): 1-20. [ Links ]
Marcano, J.S. (2001). Evaluación de la pesquería de la ostra de perla Pinctada imbricata en el oriente de Venezuela. INIA, Informe técnico de gestión anual, 18 pp. [ Links ]
Marcano, V. (1984). Aspectos biológicos de la reproducción en la ostra perla Pinctada imbricata (Röding, 1978), de Punta Las Cabeceras, Isla de Cubagua, Venezuela. Universidad de Oriente, Trabajo especial de grado, Cumaná, Venezuela, 74 pp. [ Links ]
Mendoza, Y., Freites, L., Núñez, P. y Lodeiros, C. (2001). Alternativas para el cultivo final del pectinido Lyropecten (Nodipecten) nodosus (Linne, 1758) en el golfo de Cariaco, Estado Sucre, Venezuela. En: Memorias II Congreso Suramericano de Acuicultura. Venezuela 99. Puerto La Cruz, Venezuela. [ Links ]
Munro, J.L. and Pauly, D. (1983). A simple method for comparing growth of fishes and invertebrates. ICLARM Fishbyte, 1: 5-6. [ Links ]
Narváez, N., Lodeiros, C., Freites, L., Núñez, M., Pico, D. y Prieto, A. (2000). Abundancia de juveniles y crecimiento de la concha abanico Pinna carnea (Gmelin 1791) en cultivo suspendido. Rev. Biol. Trop., 48: 785-795. [ Links ]
Novoa, D., Mendoza, J., Marcano, L. y Cárdenas, J. (1998). El Atlas Pesquero Marítimo de Venezuela. MAC-SARPA y VECEP, Caracas, 197 pp. [ Links ]
Nusetti, O., Marcano, L., Zapata, E., Nusetti, S., Esclapes, M.y Lodeiros, C. (2004). Defensas inmunológicas y estrés oxidativo en el bivalvo marino Pinctada imbricata (Mollusca: Pteridae) expuesto a niveles de Fuel Oil. Interciencia, 29(6): 324-328. [ Links ]
Pauly, D. (1983). Some simple methods for the assessment of tropical fish stocks. FAO Fish Tech. Pap., 234: 52. [ Links ]
Pauly, D. and Gaschutz, G. (1979). A simple method for fitting oscillating length data with a program for pocket calculators. ICESCA, 6: 24 pp. [ Links ]
Pauly, D. and Munro, J.L. (1984) Once more on the comparison of growth in fish and invertebrates. ICLARM Fishbyte, 2(1): 21. [ Links ]
Pouvreau, S., Gangnery, A., Tiapari, J., Lagarde F., Garmier, M. and Godoy, A. (2000). Gametogenic cycle and reproductive effort of the tropical blacklip pearl Pinctada margaritifera (Bivalvia: Pteridae), cultivated in Takapoto atoll (French Polynesia). Aquat. Living Resour., 13: 37-48. [ Links ]
Rajogopal, S., Venugolapan, V.P., Nair, K.V.K., Jenmer, H.A. and Dan Hartog, C. (2002). Reproduction growth rate and culture potential of the green mussel Perna viridis (L.) in Edaiyur backwater, east coast of India. Aquaculture, 162: 187. [ Links ]
Riascos, J. y Urban, H.J. (2002). Dinámica poblacional de Donax dentifer (Veneridae: Donacidae) en la Bahía Málaga, Pacífico colombiano durante el fenómeno "El Niño" 1997-1998. Rev. Biol. Trop., 50: 1123. [ Links ]
Rose, R., Dybdahl, R.E. and Harders, S. (1990). Reproductive cycle of the western Australian silverlip pearl oyster Pinctada maxima (Jamenson) (Mollusca: Pteridae). J. Shellfish Res., 9: 261-272. [ Links ]
Ruffini, E. (1984). Desarrollo larval experimental de la ostra perlífera Pinctada imbricata y algunas observaciones sobre su reproducción en el banco natural de Punta Las Cabeceras, Isla de Cubagua, Venezuela (trabajo especial de grado). Universidad de Oriente, Depto. de Biología, 51 pp. [ Links ]
Salaya, J.J. y Salazar, L. (1972). Exploraciones y explotaciones de la ostra perla Pinctada imbricata en Venezuela. Informe Técnico No. 44. Proyecto de Investigación y Desarrollo Pesquero. MAC/PNUD/FAO, Caracas, Venezuela, 52 pp. [ Links ]
Saucedo, P. and Monteforte, M. (1997). Breeding cycle of pearl oysters Pinctada mazatlanica and Pteria sterna (Bivalvia: Pteriidae) at Bahia de La Paz, Baja California Sur, Mexico. J. Shellfish Res., 16(1): 103-110. [ Links ]
Sokal, R. and Rohlf, F. (1995). Biometry. 3rd ed. W.H. Freeman, New York, 889 pp. [ Links ]
Strickland, J.D. and Parsons, T.R. (1972). Manual of Seawater Analysis. Bull. Fish Res. Bd. Canada, 310 pp. [ Links ]
Tranter, D.J. (1958). Reproduction in Australian pearl oyster (Lamellibrachia). I. Pinctada albina (Lamarck): Primary gonad development. Aust. J. Mar. Freshwater Res., 9: 135-143. [ Links ]
Urban, H.J. (2000). Culture potential of the pearl oyster Pinctada imbricata from the Caribbean. I. Gametogenic activity, growth, mortality and production of a natural population. Aquaculture, 189: 361-373. [ Links ]
Verginelli, R. y Prieto, A. (1991). Producción secundaria de Pinctada imbricata en una población del Golfo de Cariaco, Venezuela. Acta Cient. Venez., 42: 138-144. [ Links ]
Wilburg, K.M. and Owen, C.M. (1964). Growth. In: K.M. Wilburg and C.M. Younge (eds.), Physiology of Mollusca. Academic Press, New York, pp. 212-242. [ Links ]

