










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkCiencias marinas
Print version ISSN 0185-3880
Cienc. mar vol.30 n.3 Ensenada Sep. 2004
Artículos
Tendencias temporales (1989-1999) en las poblaciones de Haliotis fulgens y H. corrugata (Gastropoda:Haliotidae) de Isla de Cedros, Baja California, México
Temporal trends (1989-1999) in populations of Haliotis fulgens and H. corrugata (Gastropoda: Haliotidae) from Cedros Island, Baja California, Mexico
J.A. Rodríguez-Valencia*, F. Caballero-Alegría y J. Castro-González
* Instituto Nacional de la Pesca. CRIP-Ensenada Parque Industrial Fondeport Km 107 Carretera Tijuana-Ensenada El Sauzal, Baja California, Mexico. * E-mail: alejandro.rodriguez@web.de
Recibido en septiembre de 2003;
aceptado en abril de 2004.
Resumen
Este trabajo, junto con publicaciones recientes sobre la abundancia del abulón en Isla Natividad y el Archipiélago San Benito, forma un cuadro que refleja el estado y la dinámica temporal de este recurso en la costa oeste de Baja California. Se analizaron las tendencias temporales (1989-1999) de las poblaciones de Haliotis fulgens y H. corrugata de Isla de Cedros. La densidad total media de H. fulgens en el extremo norte (Punta Norte) y suroeste (San Agustín) de Isla de Cedros muestra tendencias decrecientes significativas, mientras que H. corrugata muestra esa tendencia sólo en San Agustín. Únicamente la densidad media del stock de talla legal de H. fulgens muestra tendencias significativas al decremento en la isla. La densidad extraída por pesca de H. corrugata en Punta Norte muestra una tendencia significativa al incremento, sin que la abundancia de su stock legal tenga una tendencia decreciente significativa. Los niveles de extracción autorizados para ambas especies en Isla de Cedros han sido menores al máximo permisible en México (30% del stock legal de cada especie en cada zona de captura). Los porcentajes de reclutamiento recientes en ambas especies son bajos y no mostraron tendencias temporales significativas. Aparentemente, el reclutamiento de ambas especies en Punta Norte decrece al incrementarse la temperatura del agua. En Punta Norte se encontró una relación negativa significativa entre la condición fisiológica de ambas especies y el incremento en la temperatura superficial del agua. Sin embargo, dicha relación estuvo ausente en San Agustín, sugiriendo que los efectos de los eventos cálidos pueden diferir entre localidades aledañas, o que sus poblacionales difieren en tolerancia fisiológica. La frecuencia y magnitud de las fluctuaciones en la condición fisiológica de ambas especies deberían ser tomadas en cuenta para la asignación de cuotas de captura, pues estas repercuten seguramente sobre la magnitud y calidad de los desoves y la mortalidad natural.
Palabras clave: abulón, dinámica temporal, abundancia, México.
Abstract
This work analyzes the temporal trends (1989-1999) of populations of Haliotis fulgens and H. corrugata from Cedros Island, Baja California (Mexico). The mean total density of H. fulgens in the northern (Punta Norte) and southwestern (San Agustín) points of Cedros Island showed a significant decreasing trend, while H. corrugata showed that trend only at San Agustín. Only the mean density of the legal-sized stock of H. fulgens presented a significant decreasing trend at the island. The density extracted due to fishing of H. corrugata at Punta Norte showed a significant increasing trend, but the abundance of its legal-sized stock did not show a significant decreasing trend. Authorized extraction levels for both species at Cedros Island have been lower than the maximum allowed in Mexico (30% of the legal-sized stock of each species for each fishing zone). Recent percentages of recruitment of both species are low and show no significant temporal trends. Apparently, recruitment of both species at Punta Norte decreases when the water temperature increases. A significant negative relationship was found between the physiological condition of both species and increased water temperature at Punta Norte, but such a relationship was absent at San Agustín, suggesting that the effect of warm events may differ among neighboring locations or that the populations differ in physiological tolerance. The magnitude and frequency of fluctuations in the physiological condition of both species should be taken into account when fishing quotas are assigned, since they will surely influence the magnitude and quality of spawning and natural mortality.
Key words: abalone, temporal dynamics, abundance, Mexico.
Introducción
La principal zona de captura de abulón en México se localiza en las islas y el litoral occidental de Baja California. Allí destaca Isla de Cedros, por ser la isla más grande de la zona, y porque en sus alrededores se ha llevado a cabo la pesquería del abulón desde hace 60 años. Actualmente esta pesquería está sustentada por el abulón azul (Haliotis fulgens Philippi, 1854) y el amarillo (H. corrugata Gray, 1828).
Las cuantificaciones de abulón en México iniciaron a finales de los sesenta, cubriendo prácticamente toda la costa oeste de la Península de Baja California, pero la heterogeneidad del hábitat ocasionó que sus resultados fueran poco precisos (Guzmán-del Próo, 1992). Desde fines de los ochenta la pesquería se ha regulado mediante temporadas de veda y cuotas anuales de captura estimadas a partir de cuantificaciones poblacionales limitadas a cada zona de pesca (Ramade-Villanueva et al., 1998). El diseño del método de cuantificación se dificulta por la distribución agregada de este recurso, lo que generalmente evita obtener estimaciones precisas (McShane, 1994, 1995). Los resultados publicados de las cuantificaciones mexicanas recientes comprenden la descripción de los patrones temporales de abundancia de 1988-1996 de H. fulgens y H. corrugata en Isla Natividad por Shepherd et al. (1998); los patrones temporales de abundancia (1989-1999) para las mismas especies en el Archipiélago San Benito por Rodríguez-Valencia y Caballero-Alegría (2002); y la estimación de la abundancia de Haliotis spp. en Isla San Jerónimo por Rodríguez-Valencia et al. (2002).
La intención del presente trabajo es analizar las tendencias temporales de 1989-1999 de las poblaciones de abulón azul y amarillo de Isla de Cedros, en términos de su densidad total, densidad del stock de talla legal, densidad extraída por pesca, reclutamiento, tallas, condición fisiológica, proporciones sexuales, y madurez gonadal. Estos resultados son importantes, porque completan un cuadro que refleja el estado y la dinámica temporal de las especies que soportan la pesquería mexicana del abulón en su principal zona de captura.
Materiales y métodos
Área de estudio
Isla de Cedros se localiza en el centro de la costa oeste de la Península de Baja California (fig. 1a). Allí ocurren fuertes surgencias estacionales que favorecen el crecimiento de macroalgas (González-Avilés y Shepherd, 1996). La Sociedad Cooperativa de Producción Pesquera Pescadores Nacionales de Abulón es el único concesionario para la captura de abulón y langosta alrededor de la isla. Esta cooperativa cuenta con dos campos pesqueros situados en los extremos norte y suroeste de la isla, llamados Punta Norte y San Agustín, respectivamente (fig. 1a). El abulón se captura en las inmediaciones de estos campos y, eventualmente, en algunos bancos localizados en mar abierto (20 km al oeste de la isla), pero los datos de esos bancos de mar abierto no se incluyen en este trabajo. La captura de abulón se realiza entre la línea de costa y la isobata de 30 m de profundidad. El área de captura en Punta Norte es ≈1000 hectáreas y en San Agustín es ≈1350 hectáreas (Rodríguez-Valencia et al., 1998).

Método de cuantificación
Las cuantificaciones se han realizado en las áreas de pesca de Punta Norte y San Agustín, durante el periodo de veda del abulón (octubre a enero) en 1989, 1991, 1993, 1995, 1997, 1998 y 1999. Para cuantificar el abulón, estas áreas se dividieron en bloques de ≈500 m de ancho, con ayuda de boyas. Cada bloque se extiende desde la línea de costa hasta la isobata de 30 m de profundidad, por que su longitud perpendicular a la línea de costa varía entre 130-1286 m en Punta Norte y 144-2188 m en San Agustín. Este diseño involucra 37 bloques en Punta Norte y 27 bloques en San Agustín y su ubicación ha sido la misma en todas las cuantificaciones (fig. 1a). Dentro de cada bloque se han localizado sitios de muestreo al azar, donde los buzos abuloneros de la cooperativa han sido los encargados de localizar visualmente y recolectar manualmente al abulón azul y amarillo. El número de sitios de muestreo de cada bloque y la unidad de muestreo han variado a lo largo del tiempo (fig. 1b): (a) entre 1989 y 1995 se muestreaban de 6 a 12 sitios al azar dentro de cada bloque, dependiendo del número de buzos disponibles, y la unidad de muestreo era un cuadrante de 4 m2; y (b) entre 1997 y 1999 cada bloque se dividió en tres estratos batimétricos, de 0-10 m, de 10-20 m y de 20-30 m, dentro de los que se muestrearon tres, cinco y dos sitios al azar, respectivamente, utilizando como unidad de muestreo un cuadrante de 10 m2.
Estimaciones de la densidad
Para estimar las densidades de abulón azul y amarillo únicamente se utilizaron los datos provenientes de sitios de muestreo con fondo rocoso. Se calcularon la densidad total media, obtenida considerando todas las tallas, y la densidad media del stock de talla legal (stock legal), obtenida considerando únicamente las tallas mayores a las talla mínimas legales de captura del abulón azul (150 mm de longitud de concha) y abulón amarillo (140 mm de longitud de concha).
Estimación del efecto de la pesquería sobre la abundancia
Las capturas comerciales de abulón azul y amarillo reportadas por la cooperativa para cada campo durante las temporadas de pesca 1989-1990, 1991-1992, 1993-1994, y 1995-1996 (tabla 1) se dividieron entre el promedio y los límites superior e inferior al 95% de confianza del peso del músculo (su obtención se explicará más adelante) de las evaluaciones de 1989, 1991, 1993 y 1995, respectivamente. Así se obtuvieron el promedio y los números máximo y mínimo de organismos de ambas especies que fueron extraídas durante dichas temporadas de pesca. Éstos fueron divididos entre el área total de sustrato rocoso correspondiente a cada campo, de 6,130,233 m2 en Punta Norte y 6,585,695 m2 en San Agustín (Rodríguez-Valencia et al., 1998), para obtener las densidades media, máxima, y mínima de organismos extraídos por pesca de cada especie.

Estimaciones del reclutamiento
Métodos de muestreo como el utilizado en este trabajo no proporcionan estimaciones precisas del reclutamiento, pero pueden reflejar fallas serias ocasionadas por la sobrepesca de reclutas y/o por cambios ambientales (Shepherd et al., 1998). Con base en las frecuencias de tallas, estimamos el porcentaje de reclutas de ambas especies para cada cuantificación y campo pesquero, aplicando los criterios de tallas definidos por Shepherd et al. (1998) para Isla Natividad (50-100 mm de longitud de concha para H. fulgens y 40-110 mm de longitud de concha para H. corrugata).
Obtención de datos biométricos
Se midió la longitud de las conchas (al milímetro más cercano) y se pesaron los músculos (al gramo más cercano) de todos los abulones capturados, obteniendo datos de por lo menos tres individuos por cada intervalo de talla de 5 mm. Durante las mediciones los abulones se mantuvieron en lugares sombreados y cubiertos para evitar su desecación.
El cociente de la división entre el peso del músculo y la longitud de la concha de cada organismo se utilizó como factor de condición, obteniéndose un promedio por cada especie, evaluación y campo pesquero. Durante cada evaluación se determinaron el sexo y la madurez gonadal de algunos organismos seleccionados. Vega et al. (1997) propusieron que períodos con agua cálida pueden causar variaciones en la dinámica reproductiva, y debilitamiento del abulón, repercutiendo ambos en la reducción de la abundancia. Por esta razón, probamos la relación entre los porcentajes de reclutamiento, la condición fisiológica y la madurez gonadal de ambas especies con datos de la temperatura media superficial local del agua de la National Oceanographic and Athmospheric Administration (NOAA) de los EUA (http://www.epic.noaa.gov/cgi-bin/NOAAServer) mediante regresión lineal simple. Los datos de la NOAA muestran que la temperatura se incrementó 2°C durante el período de estudio.
Análisis de los datos
Se detectaron las tendencias visuales (incremento y decremento) de las variables antes descritas, y su validez se definió mediante el análisis de varianza de regresiones lineales simples, en las cuales las variables antes descritas actuaron como variables dependientes y el tiempo actuó como variable independiente.
Resultados
Tendencias temporales de la densidad total
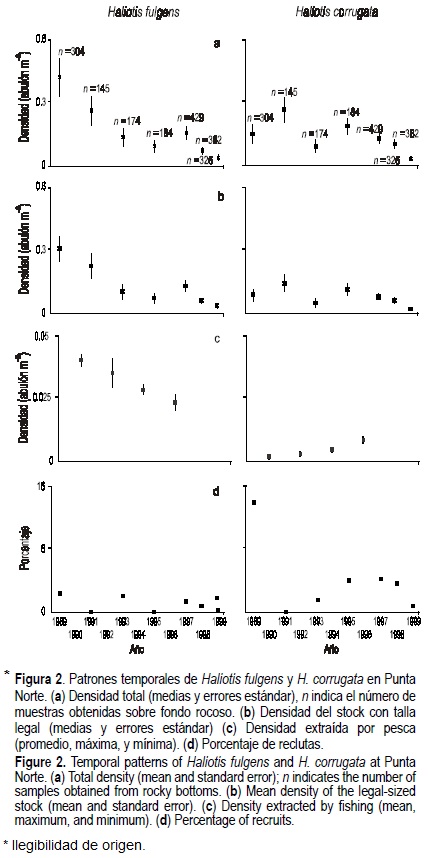
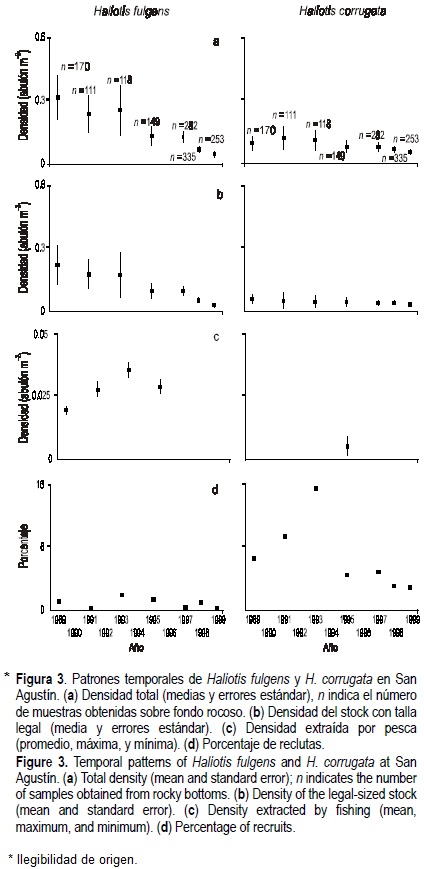
En general, las densidades totales de ambas especies en los dos campos han disminuido durante el período 1989-1999. La densidad total media del abulón azul en ambos campos muestra tendencias significativas al decremento (Punta Norte: F(1,5) = 16.2, P < 0.05; San Agustín: F(1,5) = 79.1, P < 0.01), pero la del abulón amarillo sólo ha mostrado esta tendencia en San Agustín (F(1,5) = 14.2, P < 0.05) (figs. 2a, 3a). El método de evaluación ha permitido obtener promedios con errores estándar estrechos y estables (tabla 2). Hasta 1993 los niveles de densidad total del abulón azul estaban por arriba de los del abulón amarillo en ambos campos, posteriormente ambas especies alcanzaron niveles similares.

Tendencias temporales de la densidad del stock con talla legal
Únicamente las densidades medias del stock legal de abulón azul han mostrado tendencias decrecientes significativas en ambos campamentos (Punta Norte: F(1,5) = 16.4, P < 0.01; San Agustín: F(1,5) = 114.3, P < 0.01), mientras que las del abulón amarillo no muestran esta tendencia (Punta Norte: F(1,5) = 3.3, P > 0.05; San Agustín: F(1,5) = 0.35, P > 0.05) (figs. 2b, 3b).
Tendencias temporales de la densidad extraída por pesca
Durante el período estudiado la pesquería extrajo, en promedio, 22% y 18% del stock legal del abulón azul presente en Punta Norte y San Agustín, respectivamente, correspondiendo a promedios de 17% y 13% de la densidad total de cada localidad. La extracción de abulón amarillo ha sido notoriamente menor: 5% y 9% del stock legal de Punta Norte y San Agustín, respectivamente, correspondiendo a promedios de 3% y 5% de la densidad total de cada localidad. En Punta Norte, la densidad de abulón azul extraída por pesca muestra una tendencia significativa al decremento (F(1,3) = 527.6, P < 0.01), mientras que la de abulón amarillo se ha incrementado significativamente (F(1,3) = 22.4, P < 0.05) (fig. 2c). En San Agustín, la densidad de abulón azul extraída por pesca aparentemente se incrementó entre 1989-1993 y se redujo después, sin embargo no existe una tendencia significativa al incremento (F(1,3) = 2.0, P > 0.05) (fig. 3c). No tenemos datos suficientes para evaluar la tendencia en la extracción por pesca del abulón amarillo en San Agustín, sin embargo, la densidad extraída por pesca en 1995 fue notoriamente menor a la de abulón azul (fig. 3c).
Tendencias temporales en el porcentaje de reclutamiento
En ambos campos los porcentajes de reclutamiento del abulón amarillo muestran niveles más altos que los del abulón azul (figs. 2d, 3d). El reclutamiento del abulón amarillo muestra períodos de incremento, en momentos diferentes, en ambos campos: 1993-1998 en Punta Norte y 1991-1995 en San Agustín. Los porcentajes de reclutamiento recientes de ambas especies son bajos y ningún caso mostró tendencias temporales significativas (abulón azul: F(1,6) = 0.07, P > 0.05 en Punta Norte y F(1,6) = 0.4, P > 0.05 en San Agustín; abulón amarillo: F(1,6) = 1.7, P > 0.05 en Punta Norte y F(1,6) = 2.4, P > 0.05 en San Agustín).
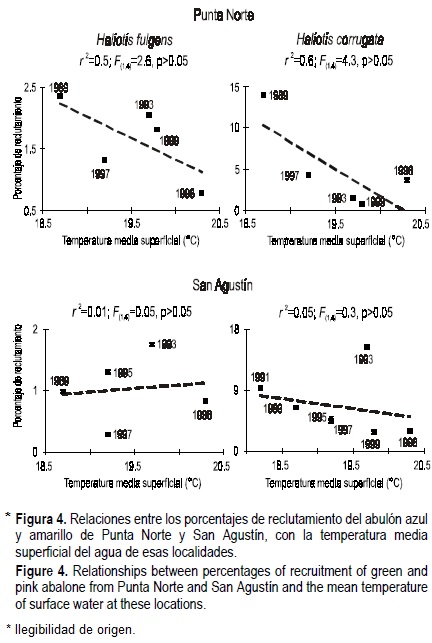
El reclutamiento de ambas especies no esta significativamente relacionado con la temperatura del agua en ambos campos. Sin embargo, dichas relaciones para el abulón azul y amarillo en Punta Norte fueron sugestivas. En estos casos, aparentemente el reclutamiento puede reducirse al incrementarse la temperatura del agua (fig. 4).
Tendencias temporales de tallas y condición fisiológica
La tabla 3 muestra la magnitud de las tallas de abulón azul y amarillo en ambos campos. La tabla también muestra la tendencia visual y su significancia estadística en cada medición. Llama la atención que las tallas de ambas especies en San Agustín muestran, principalmente, constancia temporal en sus valores. En Punta Norte, los casos con tendencias al decremento e incremento fueron más frecuentes. No obstante, únicamente las tendencias al incremento en la talla media del abulón azul de Punta Norte y en la talla máxima del abulón amarillo de San Agustín fueron estadísticamente significativas. El abulón azul siempre ha mostrado mayores porcentajes de ejemplares por encima de la talla mínima legal de captura en ambos campos pesqueros (tabla 3).
La condición fisiológica del abulón azul siempre ha mostrado niveles más altos que los del abulón amarillo en ambos campos (Punta Norte: abulón azul vs amarillo F(1,1574) = 322.2, P < 0.01; San Agustín: abulón azul vs amarillo F(1,1154) = 72.2, P < 0.01) (fig. 5 a, b). Ningún caso mostró tendencias temporales significativas (Punta Norte, abulón azul y amarillo, respectivamente: F(1,5) = 0.007, P > 0.05 y F(1,5) = 2.5, P > 0.05; San Agustín, abulón azul y amarillo, respectivamente: F(1,5) = 2.9, P > 0.05 y F(15) = 0.5, P > 0.05).
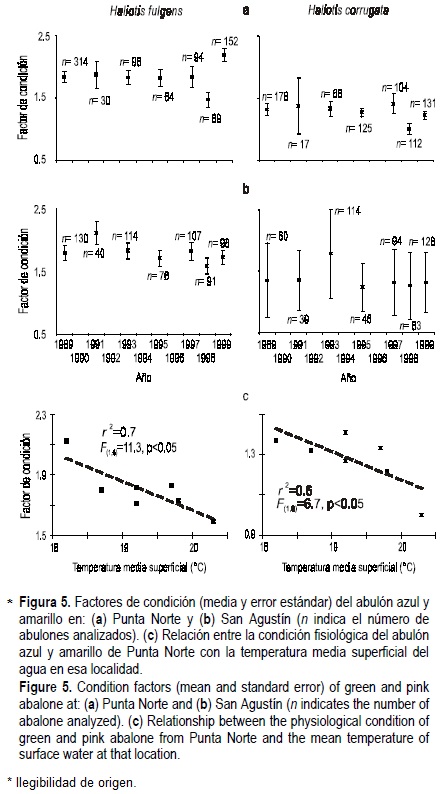
La condición fisiológica de ambas especies en Punta Norte se reduce significativamente al incrementarse la temperatura superficial del agua (fig. 5c). Sin embargo, esto no fue válido para las especies de San Agustín (abulón azul: r2 = 0.1, F(1,6) = 0.4, P > 0.05; abulón amarillo: r2 = 0.1, F(1,6) = 0.1, P > 0.05).
Tendencias temporales de la proporción sexual y madurez gonadal
La tabla 4 muestra las tendencias temporales en los porcentajes de hembras y machos de ambas especies en los dos campos. En la mayoría de los casos las tendencias visuales fueron prácticamente constantes, observándose tendencias aparentemente decrecientes sólo en los porcentajes de hembras del abulón azul de Punta Norte y de machos del abulón azul de San Agustín. No obstante, las tendencias visuales de todos los casos no fueron significativas.
La tabla 5 muestra las tendencias temporales en los porcentajes de organismos sexualmente maduros de abulón azul y amarillo en ambos campos. En 1995 se observó un pico en el porcentaje de organismos sexualmente maduros en todos los casos, el cual introdujo una gran dispersión en los valores. La gran mayoría de los casos mostraron tendencias visuales decrecientes, con excepción del porcentaje de hembras sexualmente maduras de abulón amarillo en San Agustín, en el cual se nota una tendencia aparentemente constante al ignorarse el pico de 1995. Las tendencias visuales en ningún caso fueron significativas.
En ningún caso la temperatura superficial del agua tuvo un efecto significativo sobre el porcentaje de organismos sexualmente maduros (abulón azul de Punta Norte: r2 = 0.2, F(1.6) = 1.0, P > 0.05; abulón amarillo de Punta Norte: r2 = 0.1, F(1.6) = 0.7, P > 0.05; abulón azul de San Agustín: r2 = 0.1, F(1.6) = 0.4, P > 0.05; abulón amarillo de San Agustín: r2 = 0.0, F(1.6) = 0.0, P > 0.05).
Discusión
El método de cuantificación aplicado en Isla de Cedros ha producido estimaciones de densidad con precisión aceptable. Las desventajas de este tipo de metodologías han sido comentadas por Shepherd et al. (1998) y Rodríguez-Valencia y Caballero-Alegría (2002). En el presente trabajo, sus desventajas radican en la baja proporción de hábitat potencial cubierto (Punta Norte: 0.01-0.02% del hábitat potencial entre 1989-1995 y 0.05-0.07% entre 1995-1999; San Agustín: 0.01% del hábitat potencial entre 1989-1995 y 0.04-0.05% entre 1995-1999) y la ausencia de un método específico para cuantificar juveniles. Las cuantificaciones son frecuentemente criticadas por ser de naturaleza destructiva y tener altos costos, pero pensamos que esos argumentos son injustificados pues el porcentaje de hábitat potencial que se perturba es muy bajo y los costos de las cuantificaciones apenas representan un 4% del costo total de producción de las cooperativas (Ponce-Diaz et al., 1998).
Desafortunadamente, los datos muestran claras tendencias a la reducción de la abundancia en ambos campos, igual a las mostradas por los datos publicados para Isla Natividad y el Archipiélago San Benito. El decaimiento general de la abundancia, detectado desde 1984 por Guzmán-del Próo (1992), sigue predominando, y los niveles de densidad son cercanos a aquellos en los pueden darse fallas en el reclutamiento (Shepherd y Partington 1995). Únicamente el abulón amarillo ha mostrado algunos indicios de recuperación en la región, reflejados en leves repuntes en su abundancia detectados en 1996 en Isla Natividad (Shepherd et al., 1998) y en 1999 en el Archipiélago San Benito (Rodríguez-Valencia y Caballero-Alegría 2002), así como por la ausencia de tendencias decrecientes significativas en su densidad total en Punta Norte y su densidad del stock legal en ambas zonas de pesca de Isla de Cedros. Aquí se muestra que la extracción del abulón amarillo por la pesquería ha sido notablemente menor que la de abulón azul, y es posible que esto haya contribuido a las leves muestras de recuperación del primero.
Al analizar las tendencias de las densidades extraídas por pesca observamos que únicamente la del abulón amarillo en Punta Norte se ha incrementado significativamente. Sin embargo, los niveles de extracción autorizados para ambas especies en Isla de Cedros se han mantenido por debajo del máximo permisible para las cuotas de captura (30% del stock legal de cada especie, ver Shepherd et al., 1998). Aparentemente el incremento en la densidad extraída por pesca del abulón amarillo no ha afectado la abundancia de su stock legal, ya que ésta no muestra tendencias al decremento en ninguno de las zonas de pesca de Isla de Cedros.
El fácil acceso a Isla de Cedros por vía marina y aérea incrementa el riesgo de la captura ilegal. Legalmente, sólo la cooperativa local de pescadores está autorizada para extraer abulón, pero los miembros del equipo de vigilancia de esta cooperativa han detectado casos de pesca ilegal. Escuelas y clubes de buceo mexicanos y estadounidenses también visitan frecuentemente la zona, y aunque no existen datos oficiales, hay testimonios de que ocasionalmente ellos también extraen abulón.
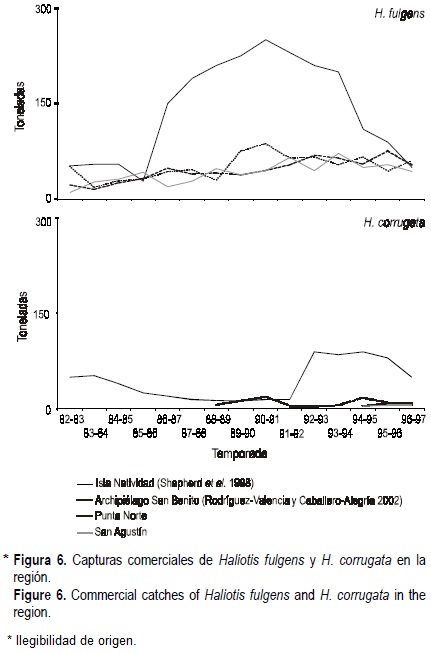
Desafortunadamente, no se cuenta con estimaciones de la mortalidad por pesca, pero podríamos esperar que éstas fueran menores a las reportadas para Isla Natividad por Shepherd et al. (1998), pues ahí las capturas comerciales de ambas especies han sido notoriamente mayores a las de Isla de Cedros (fig. 6). Con base en las capturas comerciales esperaríamos que la mortalidad por pesca en Isla de Cedros y el Archipiélago San Benito fueran similares. Ya hemos visto que la pesquería ejerce mayor presión sobre el abulón azul, y por ello deberíamos esperar que la mortalidad por pesca fuera mayor para esa especie. La preferencia por el abulón azul se debe a su mayor abundancia en tallas legales, a que se obtiene mayor cantidad de carne por pieza, y a que su concha tiene mayor precio.
Los patrones temporales de los porcentajes de reclutamiento de ambas especies son similares a los reportados para el Archipiélago San Benito por Rodríguez-Valencia y Caballero-Alegría (2002). Las relaciones entre el porcentaje de reclutamiento de ambas especies y la temperatura del agua no fueron significativas, pero las sugestivas relaciones observadas para el abulón amarillo y azul de Punta Norte apoyan la propuesta de Vega et al. (1997), acerca de un posible efecto negativo de los eventos cálidos sobre el reclutamiento del abulón. Por otro lado, los patrones de reclutamiento observados para el abulón amarillo apoyan la propuesta de Shepherd et al. (1989) en el sentido de que los eventos cálidos no intensos pueden afectar positivamente su reclutamiento: En términos generales, el reclutamiento del abulón amarillo en Isla de Cedros pudo haberse incrementado hasta 1995 porque sus larvas se asentaron durante condiciones "El Niño" no severas, mientras que éste se redujo a partir de 1996 porque los asentamientos ocurrieron durante frecuentes eventos "El Niño" severos. Rodríguez-Valencia y Caballero-Alegría (2002) también encontraron estos patrones en el Archipiélago San Benito. Asímismo, la aparente diferencia detectada aquí entre las respuestas del reclutamiento del abulón azul y amarillo al incremento en la temperatura del agua coincide con los resultados de Rodríguez-Valencia y Caballero-Alegría (2002) para el Archipiélago San Benito, sugiriendo que los eventos cálidos afectan en mayor grado el reclutamiento del abulón azul.
La detección de una relación negativa significativa entre la condición fisiológica de ambas especies con el incremento en la temperatura del agua en Punta Norte también coincide con los resultados de Rodríguez-Valencia y Caballero-Alegría (2002) en el Archipiélago San Benito. La ausencia de estos efectos en San Agustín sugiere que los efectos de los eventos cálidos pueden diferir entre localidades aledañas, o bien, que sus poblacionales difieren en tolerancia fisiológica. Del Río-Portilla (2000) y Del Río-Portilla y González-Aviles (2001) demostraron diferencias genéticas entre las poblaciones de abulón amarillo del Archipiélago San Benito e Isla de Cedros.
Las propuestas de Vega et al. (1997) y Shepherd et al. (1998) sugieren que la reducción en la abundancia del abulón puede deberse, parcialmente, al deterioro en su condición fisiológica durante los eventos "El Niño". Nosotros pensamos, al igual que Rodríguez-Valencia y Caballero-Alegría (2002), que la frecuencia y magnitud de las fluctuaciones en condición deben ser tomadas en cuenta al momento de asignar las cuotas de captura, ya que éstas seguramente repercuten en la magnitud y calidad de los desoves, así como en la mortalidad natural.
Agradecimientos
Dedicamos este trabajo a la memoria de F. Salgado, G. Pérez, y A. Lelevier. La ayuda de todos los miembros de la Sociedad Cooperativa de Producción Pesquera "Pescadores Nacionales de Abulón", y en especial de D.G. Romero-Árce, ha sido fundamental para poder realizar las evaluaciones. Nuestros colegas F. Uribe, J. Cordova, J.L. Rivera, J. Castro, R. Sánchez, L. Vélez, M. Navarrete, J. Talavera, y O. Pedrín-Osúna apoyaron de forma importante la realización de las evaluaciones. Agradecemos las mejoras al manuscrito propuestas por cuatro revisores anónimos.
Referencias
Del Río-Portilla, M.A. (2000). Population genetics of the yellow abalone, Haliotis corrugata, in Cedros and San Benito Islands. J. Shellfish Res., 19(1): 508. [ Links ]
Del Río-Portilla, M.A. and González-Aviles, J.G. (2001). Population genetics of the yellow abalone, Haliotis corrugata, in Cedros and San Benito Islands: A preliminary survey. J. Shellfish Res., 20(2): 765-770. [ Links ]
González-Aviles, J.G. and S.A. Shepherd. (1996). Growth and survival of the blue abalone Haliotis fulgens in barrels at Cedros Island, Baja California, with review of abalone barrel culture. Aquaculture, 140: 169-76. [ Links ]
Guzmán-del Próo, S.A. (1992). A review of the biology of abalone and its fishery in Mexico. In: S.A. Shepherd, M.J. Tegner and S.A. Guzmán-del-Proo (eds.), Abalone of the World. Fishing News Books, pp. 341-360. [ Links ]
McShane, P.E. (1994). Estimating the abundance of abalone (Haliotis spp.): Stock examples from Victorian and southern New Zealand. Fish. Res., 19: 379-394. [ Links ]
McShane, P.E. (1995). Estimating the abundance of abalone: The importance of patch size. Mar. Freshwater Res., 46: 657-662. [ Links ]
Ponce-Díaz, G., Vega-Velázquez, A., Ramade-Villanueva, M., León-Carballo, G. and Franco-Santiago, R. (1998). Socioeconomic characteristics of the abalone fishery along the west coast of the Baja California Peninsula, Mexico. J. Shellfish Res., 17(3): 853-857. [ Links ]
Ramade-Villanueva, M., Luch-Cota, D.B., Lluch-Cota, S.E., Hernández-Vazquez, S., Espinoza-Montes, A. and Vega-Velázquez, A. (1998). An evaluation of the annual quota mechanisms as a management tool in the Mexican abalone fishery. J. Shellfish Res., 17(3): 847-851. [ Links ]
Rodríguez-Valencia, J.A. and Caballero-Alegría, F. (2002). Temporal fluctuations (1989-1999) in the populations of Haliotis fulgens and H. corrugata (Gastropoda: Haliotidae) at Islas San Benito (Baja California, Mexico). J. Shellfish Res., 21(1): 163-172. [ Links ]
Rodríguez-Valencia, J.A., Caballero-Alegría, F., Castro-González, J., Sánchez-Romero, R. y Navarrete-Gutiérrez, M. (1998). Evaluación del abulón (Haliotis spp.) en los campos de Punta Norte, San Agustín, y el Bajo del Coloradito, Isla de Cedros, B.C. (6 al 16 de marzo de 1998). Rep. Téc. CRIP-Ensenada (CRIP/335/ 98). [ Links ]
Rodríguez-Valencia, J.A., Caballero-Alegría, F., Uribe-Osorio, F. and Arano-Castañon, A. (2002). Abundance and associations of two commercially important gastropods (Astraea and Haliotis) in Isla San Jeronimo, Baja California, Mexico. Cienc. Mar., 28(1): 49-66. [ Links ]
Shepherd, S.A. and Partington, D. (1995). Studies on southern Australian abalone (Genus Haliotis). XVI. Recruitment, habitat and stock relations. Mar. Freshwater Res., 46: 669-80. [ Links ]
Shepherd, S.A., Turrubiates-Morales, J.R. and Hall, H. (1998). Decline of the abalone fishery at La Natividad, Mexico: Overfishing or climate change? J. Shellfish Res., 17(3): 839-846. [ Links ]
Vega, A., Luch-Belda, D., Muciño, M., León. G., Hernández, S., Luch-Cota, D., Ramade, M. and Espinoza, G. (1997). Development, perspectives, and management of lobster and abalone fisheries off northwest Mexico under a limited access system. In: D.A. Hancock, D.C. Smith, A. Grant and J.P. Beumer (eds.), Developing and Sustaining World Fisheries Resources. CSIRO, Collingwood, Victoria, Australia, pp. 136-142. [ Links ]

