










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkCiencias marinas
Print version ISSN 0185-3880
Cienc. mar vol.29 n.3 Ensenada Sep. 2003
Artículos
Caracterización multiescala de la temperatura del mar en una zona de bancos de abulón (Haliotis spp.) en Bahía Asunción, Baja California Sur, México
Multiscale characterization of the sea temperature in an area of abalone banks (Haliotis spp.) at Bahía Asunción, Baja California Sur, Mexico
Germán Ponce-Díaz1,2*, Salvador E. Lluch-Cota1, J. Jesús Bautista-Romero1 y Daniel Lluch-Belda2
1 Centro de Investigaciones Biológicas del Noroeste, S.C., Mar Bermejo #195, Col. Playa Palo de Santa Rita, Apartado postal 128, La Paz, CP 23090, Baja California Sur, México. *E-mail: gponce@cibnor.mx
2 Centro Interdisciplinario de Ciencias Marinas, Terrenos "El Conchalito" s/n, La Paz, Baja California Sur, México.
Recibido en marzo de 2002;
aceptado en abril de 2003.
Resumen
La pesquería de abulón en Baja California es una de las más importantes en México debido a su alto valor comercial. A pesar de diversos esfuerzos de manejo, esta pesquería ha sufrido cambios drásticos que han sido atribuidos principalmente a la sobrepesca y a la variabilidad del clima marino. El presente trabajo analiza la variabilidad de la temperatura del mar en la región de Bahía Asunción, BCS, a escalas temporales que pueden afectar procesos biológicos del abulón tales como mortalidad, reproducción, reclutamiento y crecimiento individual, entre otros. El estudio se basa en diferentes fuentes de información. Se presenta el ciclo circadiano tipo y sus variaciones a lo largo del año y entre años para el periodo 1997-2000, el ciclo estacional y desviaciones interanuales para el periodo 1992-2000, la variabilidad a escala decadal sobre la base de valores anuales promedio del periodo 1959-1997 y el comportamiento de largo plazo con datos de todo el Siglo XX. Adicionalmente, se presenta un análisis de densidad espectral donde se estimaron los valores de contribución porcentual de cada región espectral (escala temporal) a la señal de variabilidad completa. Los resultados de variaciones de alta frecuencia (circadiana a interanual) sugieren una relación entre los ciclos reproductivos y de variación térmica, un efecto potencial negativo de los fenómenos ENSO de gran intensidad sobre las postlarvas de abulón con menor tolerancia a altas temperaturas, así como un efecto diferencial de la temperatura sobre juveniles de diferentes especies. Respecto de la baja frecuencia (decadal a siglo), es evidente que los años recientes representan una condición mucho más cálida que la que persistió durante las décadas anteriores, lo que sugiere estados diferentes de las comunidades biológicas en la región. En el área de estudio es notoria la ausencia de información sobre aspectos del clima marino, particularmente acerca de mediciones finas o de alta frecuencia.
Palabras clave: Haliotis, temperatura superficial del mar, Bahía Asunción, variabilidad climática, ENSO.
Abstract
The Baja California abalone is one of the most important Mexican fisheries because of its high economic value. Despite strong management efforts, this fishery has faced dramatic fluctuations attributed mainly to overfishing and variability of the marine climate. In this report we analyze sea temperature variability off Bahía Asunción, Baja California Sur (Mexico), at temporal scales that may affect abalone biological processes, such as mortality, reproduction, recruitment, and individual growth rate. Since the analysis is based on different data sources, we conducted a series of correlation analyses to determine the coherence between them. We report the average daily cycle and its variations throughout the year and between years for the period 1997-2000, the seasonal cycle and interannual deviations for the period 1992-2000, the behavior of the decadal scale variability based on annual values for the period 1959-1997, and the long-term trends after analyzing the entire 20th century. Furthermore, we conducted a spectral density analysis to estimate the proportional contribution of each temporal scale to the entire variability signal. Results from the high-frequency variability (daily to interannual) suggest a relationship between temperature and the abalone reproductive cycles, a potential negative effect of the very strong ENSO events on the postlarvae, and a differential effect of temperature on juveniles of different species. Regarding the low-frequency variability (decadal to century), we show that recent years represent a much warmer period compared to previous decades, suggesting a different biological community structure between periods. The spectral density analysis indicates seasonal variability as the major contributor to the regional variability, followed by the interannual (related to ENSO). There is almost no information of the marine climate in this region, particularly in regard to fine and high-resolution observations. This contribution will provide new quantitative elements for studies dealing with the ecology of these coastal productive systems.
Key words: Haliotis, sea surface temperature, Bahía Asunción, climate variability, ENSO.
Introducción
Los abulones son gasterópodos marinos bentónicos que viven desde la zona intermareal hasta los 30-40 m de profundidad (Guzmán-del Próo, 1992). Estos organismos habitan en zonas rocosas entre bosques de macroalgas y asociados a una comunidad de gran riqueza específica. Las especies de abulón de mayor talla se encuentran en zonas que presentan corrientes con aguas frías y templadas a lo largo de las costas continentales donde predominan las algas cafés (Cox, 1962).
En México la producción pesquera de abulón de los últimos años ha oscilado alrededor de las 350-400 t anuales de callo, con un valor estimado entre 17 y 36 millones de dólares (Ponce-Díaz et al., 1998; INP, 2000). En años recientes la pesquería mexicana de abulón ha sufrido una disminución drástica de sus capturas (INP, 2000). Esta situación se ha relacionado con procesos de sobrepesca (Breen, 1992; Guzmán-del Próo 1992; Prince y Guzmán-del Próo 1993); sin embargo, alternativamente se ha propuesto la idea de que los bajos rendimientos de esta pesquería podrían ser explicados en parte por la variabilidad del clima marino (Guzmán-del Próo, 1994; Vega et al., 1995; Lluch-Belda et al., 1997; Shepherd et al., 1998; Ponce-Díaz et al., 2000).
La temperatura ambiental ha sido señalada como uno de los factores importantes que influyen en la distribución y abundancia de los organismos. Se han mencionado patrones y cambios de largo plazo de la variabilidad interanual dentro del sistema de la Corriente de California, la cual tiene influencia en el área de distribución del recurso abulón (Lluch-Belda et al., 2001). Por ejemplo, en Bahía Magdalena, ubicada al sur de la Península de Baja California, se ha observado que los cambios de temperatura se asocian a eventos intra e interanuales y presentan una considerable magnitud de cambio, lo que implica que las especies que habitan esta región deben estar adaptadas a muy diferentes condiciones térmicas (Lluch-Belda et al., 2000).
En ese sentido, es importante analizar la variabilidad del clima marino en términos de los cambios de la temperatura del mar a las diferentes escalas temporales (circadianas, estacionales, interanuales y de largo plazo) a que están expuestos los recursos pesqueros, en este caso el abulón. Tales cambios de temperatura pueden tener consecuencias sobre procesos biológicos que inciden en la productividad del recurso, tales como mortalidad, reproducción, reclutamiento y crecimiento individual, entre otros.
El presente trabajo muestra una caracterización de la variabilidad de la temperatura del mar en la región de Bahía Asunción, analizando diferentes ciclos de variación, desde los circadianos hasta los decadales, incluyendo eventos de El Niño, junto con una visión de largo plazo para identificar la tendencia general durante el siglo más reciente.
Área de estudio
Bahía Asunción (fig. 1) se encuentra enclavada en la parte central de la costa occidental de la Península de Baja California y presenta una línea costera en donde se alternan litorales rocosos con zonas de playa arenosa. A lo largo de este litoral ocurren especies de importancia comercial como el abulón y la langosta (Vega et al., 1995; León-Carballo y Muciño-Díaz, 1996; Vega-Velázquez et al., 1996). En la zona de Bahía Asunción, al menos, se tienen cuantificados 50 bancos en producción de Haliotis fulgens Philippi, 1854 y H. corrugata Wood, 1828 (Muñoz, 1976), en asociación con macroalgas de la especie Eisenia arborea Areschoug 1876 y, en menor grado, con Macrocystis pyrifera (L.) C. Agardh 1820, la cual está limitada a la región norte de esta área de estudio (Hernández-Carmona et al., 2000).
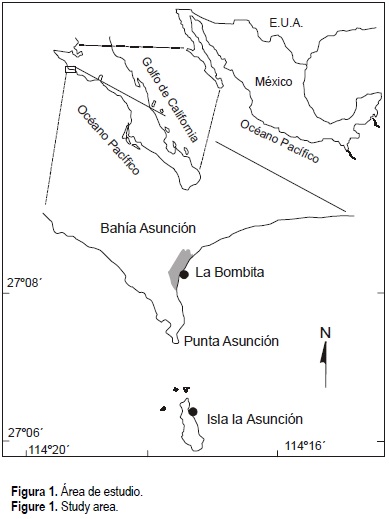
La parte sur del área de estudio (en las inmediaciones de Punta San Hipólito) es considerada como una zona de surgencias temporales y de giros que propician altas producciones biológicas en estos litorales (Walsh et al., 1974, 1977). De hecho, se ha documentado el papel que tienen los nutrientes provenientes de estos procesos oceanográficos sobre las poblaciones de macroalgas en la región de estudio (Hernández-Carmona et al., 2000), mismas que son la fuente primordial de alimento para abulones.
En esta parte de la costa occidental de la Península de Baja California ocurren eventos oceanográficos que determinan que el área de estudio sea considerada como una zona de transición entre aguas templadas y tropicales y, por ende, una frontera provincial de varios grupos taxonómicos, incluyendo peces costeros, braquiuros, briozoarios y moluscos (Hewitt, 1981). Asimismo, esta porción de la costa occidental de la península ha sido caracterizada por la presencia de variaciones de periodo corto en la circulación anticiclónica, o giros oceánicos con intensificaciones o relajamientos de surgencias (Amador-Buenrostro et al., 1995).
Material y métodos
Los datos de temperatura utilizados en este trabajo provienen de diversas fuentes:
(a) Datos de temperatura in situ registrados cada hora o cada dos horas. Las mediciones se hicieron a una profundidad de entre 5 y 6 m, por medio de un termógrafo de registro continuo en un banco abulonero cercano a la Isla Asunción, a una distancia aproximada de 300 m de la línea de costa durante el periodo de 1997 a 2000 (Ponce-Díaz et al., 2003).
(b) Datos de temperatura superficial del mar registrados cada 24 h en el interior de Bahía Asunción, en el periodo de 1992 a 2000 (Ponce-Díaz et al., 2003).
(c) Datos de temperatura superficial del mar de una serie integrada y proporcionada por el Dr. Arthur V. Douglas (Department of Atmospheric Sciences, Creighton University, EUA), la cual consiste en registros mensuales para el cuadrante 115°-116° W y 27°-28° N, derivados a partir del análisis de regresión lineal múltiple entre mediciones de temperatura superficial del mar y datos de anillos de crecimiento de árboles (Douglas, 1976, 1980). Se utilizó esta información a partir 1959 y hasta 1997, años para los que se tienen datos sobre la pesquería de abulón en la zona de estudio.
(d) Datos de temperatura superficial del mar de la base COADS (Comprehensive Ocean-Data Set/National Oceanic and Atmospheric Administration) con registros mensuales para el cuadrante contiguo a la zona de estudio (114°-116° W, 26°-28° N), que contiene información desde 1900 hasta 1996, y que son resultado de observaciones de muestreos directos con boyas oceanográficas y registros tomados en barcos de oportunidad (Woodruff et al., 1993).
Análisis de datos
A partir de las fuentes de información básicas de temperatura del mar registrada con frecuencia horaria y cada 24 h, se realizó un análisis de correlación para determinar la correspondencia entre las series. Asimismo, se llevó a cabo este análisis entre los datos de mediciones de la temperatura superficial del mar realizadas cada 24 h y la serie de datos de temperatura mensual media de Douglas, y entre esta última y la serie de COADS.
Ciclo circadiano
El ciclo circadiano tipo se estimó a partir de los promedios de todos los valores horarios registrados por el termógrafo para 2000 y se ajustó un ciclo de dos armónicos. Posteriormente se calculó la forma en que varían los ciclos diarios a lo largo del año mediante los promedios de invierno (diciembre a febrero), primavera (marzo a mayo), verano (junio a agosto) y otoño (septiembre a noviembre) de 2000. Para cada caso se ajustó un ciclo.
Para evaluar la magnitud de los cambios en el ciclo circadiano que pueden esperarse entre años, se calculó el valor mensual promedio de amplitud del ciclo diario (valor máximo-mínimo) y se comparó con el valor estimado de amplitud sobre la base del año 2000.
Ciclo estacional
En la serie de mediciones de escala interanual se presentaron, al menos, dos eventos intensos de la Oscilación del Sur El Niño (ENSO, por sus siglas en inglés) durante el periodo de estudio (1992-1993 y 1997-1998); por esta razón los análisis de escala circadiana y estacional se realizaron tomando para todos los cálculos el año 2000 como referencia, ya que se puede considerar un año cercano a las condiciones medias de los años recientes.
La variabilidad estacional fue tratada mediante el promedio de los datos diarios de la variable registrada, de los que se obtuvieron los valores mensuales de temperatura del mar.
Variabilidad interanual
Los valores correspondientes a 2000 se ajustaron a un ciclo de dos armónicos, el cual se utilizó para estimar desviaciones o anomalías de la temperatura superficial del mar para el periodo 1992-2000.
Variabilidad de largo plazo
La variabilidad de largo plazo fue determinada mediante la utilización de los promedios anuales de la temperatura superficial del mar, derivados a partir de la base de datos de Douglas para el cuadrante 1° x 1° más contiguo a la zona de estudio, y de la base COADS para el cuadrante 114°-116° W, 26°-28° N.
Dado que las anomalías de la serie Douglas están referidas al periodo 1959-1997, se calculó la relación entre éstas y las del cuadrante COADS, que cubre desde 1900 hasta 1996 (r = 0.909**, n = 41), con una diferencia entre ambas de + 0.2827°C.
Partiendo de la información de COADS, para el análisis de la condición general de la zona con respecto a la variación de largo plazo se compararon las anomalías anuales obtenidas con las tendencias suavizadas mediante un filtro Hamming de 10 años, a lo largo del siglo, en el cuadrante seleccionado.
Análisis espectral
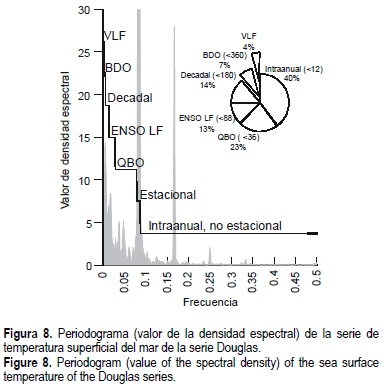
La serie Douglas, la cual coincide con los años de información de la pesquería de abulón en la zona, fue tratada mediante técnicas de series de tiempo para calcular la contribución relativa de las diferentes escalas de variabilidad a la señal completa de la serie. El procesamiento incluyó la eliminación de la media y la tendencia, la aplicación de un filtro pesado Hamming de 9 términos, y se estimaron las densidades espectrales derivadas de una transformación rápida de Fourier. Posteriormente, se agruparon los valores de densidad espectral para regiones definidas por su frecuencia: intraanual no estacional (< 11 meses); estacional (11-13 meses); oscilación guasi-bianual (QBO, por sus siglas en inglés) que incluye la alta frecuencia del ENSO (13-36 meses); ENSO (su baja frecuencia; 36-88 meses); decadal (88-180 meses); bidecadal (180-360 meses); y de muy baja frecuencia (> 360 meses). Para cada categoría se promediaron los valores de densidad espectral y se ponderaron sobre la base del recorrido correspondiente de las frecuencias. Finalmente, se estimaron los valores de contribución porcentual de cada región espectral a la señal completa y se estimó de nueva cuenta eliminando el estacional.
Resultados
El análisis de correlación entre las mediciones de temperatura del mar, temperatura in situ y temperatura superficial del mar registradas de manera diaria en las inmediaciones de la Bahía Asunción, y su relación con la derivada de la serie Douglas y, a su vez, de ésta última con respecto a la de COADS, es estadísticamente significativa en todos los casos, por lo que el análisis llevado a cabo tiene un soporte adecuado.
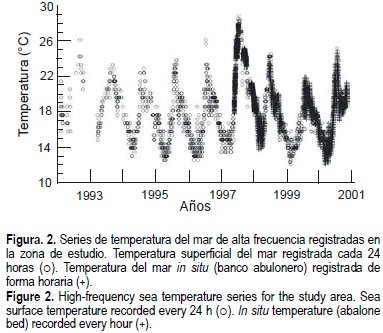
En la figura 2 se muestran las series de datos de mediciones diarias y horarias en la zona de estudio. En ella se observa la presencia de dos eventos ENSO intensos, 1992-1993 y 1997-1998; el rango de la temperatura para las series oscila entre los 11°C y 29°C aproximadamente.
La relación funcional entre los datos del termógrafo de registro continuo y los datos tomados en la estación de registro diario en el interior de Bahía Asunción alcanza un coeficiente de correlación r = 0.952 con n = 704 (fig. 3a). La relación entre los datos de la serie histórica de Douglas y los registros diarios de temperatura (fig. 3b) muestra igualmente una correlación alta con un coeficiente r = 0.909 y con n = 49.

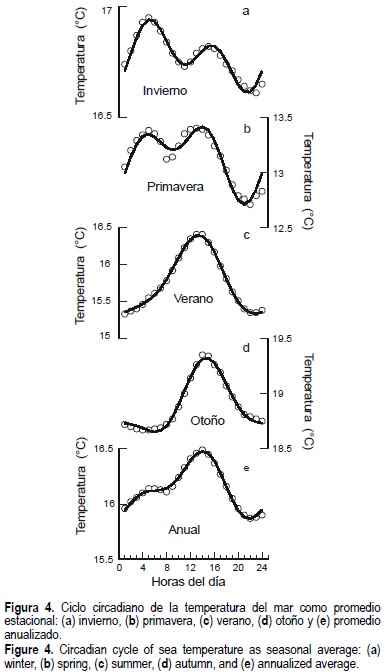
La variabilidad de la temperatura con una escala circadiana se analiza con una perspectiva anual y estacional. Se observa que tanto en invierno como en primavera se presenta un ciclo circadiano con dos máximos al día. En la estación de primavera se presentan las temperaturas del mar más bajas (fig. 4a, b).
Por otra parte, esta escala circadiana se analiza en términos de las variaciones en la amplitud de este ciclo con respecto a una comparación entre años (fig. 5). En 2000, la amplitud del ciclo diario varía estacionalmente de alrededor de 0.6°C en el invierno hasta alrededor de 1.5°C en el verano, mientras que durante el periodo previo las amplitudes son, en general, más bajas y constantes a lo largo del año, especialmente durante El Niño 1997-1998.

Tanto el patrón estacional con base en los datos de 2000 como las desviaciones de este patrón, considerando los años de 1992-1999, se presentan en la figura 6. En la figura 6a se aprecia que la temperatura más baja se presentó durante el mes de mayo, mientras que la más alta ocurrió en septiembre. Por otra parte, en la figura 6b, de las desviaciones de la serie se observan con claridad la presencia de los eventos El Niño de 1992-1993 y 1997-1998, así como el fenómeno opuesto (La Niña) durante los años 1999-2000.

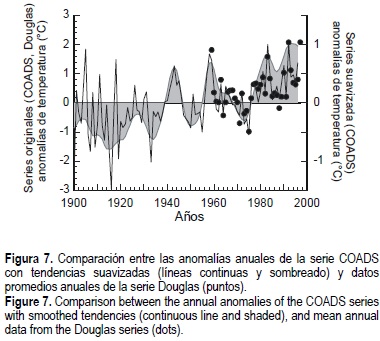
En la figura 7 se comparan las anomalías anuales medias entre la serie Douglas, ajustadas a la diferencia observada con respecto a la serie COADS del cuadrante seleccionado, sobre un marco general de tendencias suavizadas desde 1900.
En la figura 8 se aprecian los valores del periodograma y la contribución relativa de las señales de los componentes climáticos detectados por el análisis en esta caracterización ambiental de la zona de estudio. Los componentes estacional e interanual que reporta el análisis espectral resultan de particular importancia.
Discusión
Leighton (1974) mencionó que las postlarvas de H. corrugata y H. fulgens (abulón amarillo y azul, respectivamente) presentan límites de tolerancia a la temperatura entre los 23.5°C y 26.0°C, aunque las mayores sobrevivencias (cercanas al 100%) fueron típicamente con valores intermedios de temperaturas dentro de un rango de 7°C a 31°C, y se observó un rápido aumento en la mortalidad en las vecindades de los valores extremos. Lo anterior sugiere un efecto negativo potencial de los fenómenos ENSO de gran intensidad sobre las postlarvas de abulón, con menor tolerancia a valores altos de temperatura que pudiera estar ocurriendo en Bahía Asunción, si se toman en cuenta a los valores absolutos de temperatura in situ observados. Tal es el caso de las temperaturas más altas reportadas en este trabajo para esta área (28-29°C) y que coinciden con las épocas reproductivas de estas especies.
Las tasas de crecimiento son diferenciales para los juveniles de H. corrugata y H. fulgens; los mayores incrementos se alcanzan a los 24°C para la primera especie mientras que, para la segunda, se dan entre los 27°C y 28°C (Leighton, 1974). Lo anterior indica que el abulón azul durante esa etapa podría verse favorecido con incrementos de temperatura derivados de los eventos ENSO no intensos si, por supuesto, existiera alimento adecuado disponible. Existe información de campo sobre juveniles de abulón que permite plantear esta hipótesis (Rodríguez-Valencia y Caballero-Alegría, 2002).
La presencia de las temperaturas más bajas en primavera coincide con los fenómenos de surgencias reportados para la zona (Walsh et al., 1974, 1977). En invierno y primavera se observa un ciclo circadiano con dos máximos durante el día (fig. 4a, b), mientras que en verano y otoño el ciclo está marcado por la presencia de un solo máximo hacia mediodía y mínimos en las primeras horas del día (fig. 4c, d). En otoño es cuando se tiene reportado el mayor pico del desove de abulón (Guzmán-del Próo, 1992; Belmar y Guzmán-del Próo, 1992) y, en este caso, es cuando se observan las temperaturas más altas (19.5°C), por lo que asumimos que a estas temperaturas se disparan mecanismos reproductivos de las especies de abulón de Bahía Asunción.
El promedio del ciclo circadiano anualizado se comporta de manera muy parecida a lo que ocurre en verano y otoño, por lo que es de suponer que estos patrones dominan el promedio anual. Por otra parte, es importante mencionar que, si bien la variación en magnitud de la temperatura entre mínimos y máximos se encuentra prácticamente en todas las estaciones del año dentro de los límites de 1°C, en valores absolutos se registraron desde los 12.5°C como mínimo en primavera hasta aproximadamente 19°C como máximo en otoño, lo que hace una diferencia de hasta 6.5°C en el ciclo circadiano, por estación, durante el año (fig. 4 a-d).
El ciclo mensual ajustado muestra claramente la estacionalidad de la temperatura del mar. Abril y mayo son los meses con temperaturas más bajas, en contraste con agosto, septiembre y octubre que son los de temperaturas con mayor magnitud. Las desviaciones de la serie de temperaturas diarias, con respecto a 2000, muestran claramente desviaciones positivas durante los años de 1992-1993 y 1997-1998, tal y como era de esperarse dada la presencia del fenómeno de El Niño en esos años a los que les sigue un periodo de temperaturas más bajas en general.
Respecto al comportamiento interanual de la temperatura del mar, referido a 2000 (fig. 5) se aprecia que particularmente las diferencias en la amplitud del ciclo diario son menores en los años 1997-1998, o año Niño, que en otro periodo. Esto significaría que las altas temperaturas que se presentaron en estos años se mantuvieron relativamente constantes, a diferencia de lo observado en los otros años; adicionalmente, la variación es más amplia hacia la época invernal.
Es de resaltar que las condiciones de largo plazo en general están identificadas con una tendencia de temperaturas relativamente bajas hasta finales de los años treinta. Posteriormente, se presenta una etapa de marcada variabilidad desde principios de los años cuarenta hasta mediados de los setenta y, a partir de la segunda mitad de dicha década, se presenta un cambio hacia temperaturas más altas como lo muestran las anomalías de la temperatura superficial del mar (fig. 7). Esta condición de aumento de temperatura de manera constante y sostenida, sugiere que las condiciones de la comunidad donde se desarrolla el abulón, pudiesen verse alteradas paulatinamente y potencialmente afectar en alguna medida la biología poblacional de cada una de las especies de este gasterópodo que se distribuyen en la zona de estudio, o bien de alguna otra especie relevante de la comunidad (Guzmán-del Próo, 1994; Lluch-Belda et al., 1997; Shepherd et al., 1998).
En la perspectiva del siglo, es evidente que los años recientes (después de mediados de los setenta) presentan una condición mucho más cálida que la que persistió durante las décadas anteriores. De hecho, la región era alrededor de 1.3°C más fría durante las primeras décadas del siglo (1900-1935) que en los tiempos recientes (1978-1996). El periodo de inicio y desarrollo de la pesquería (1950-1975) fue, en promedio, 0.7°C más frío que el actual.
Considerando la caracterización ambiental con perspectiva de siglo, es posible que un aumento sostenido de la temperatura en los últimos años pudiera haber afectado mayormente a la flora y la fauna bentónicas característica de la comunidad rocosa de los bancos pesqueros de abulón de la zona: Macrocystis pyrifera, Eisenia arborea, Astraea undosa Wood 1828, Haliotis fulgens, H. corrugata, Bossiella orbigniana (Dec.) Silva 1957, Corallina officinalis Linnaeus 1758, C. pinnatifolia (Manza) Dawson 1953, Megathura crenulata Sowerby 1825 y Phyllospadix torreyii Hodgson 1980; ya que en este grupo, salvo por las especies de Corallina que tienen origen subtropical, el componente principal es de origen templado (Guzmán-del Próo et al., 1991). Esta hipótesis es coherente con los resultados del trabajo de Barry et al. (1995), quienes reportan un cambio de largo plazo en la composición de especies de invertebrados en una comunidad rocosa intermareal en la parte central de California, EUA, en la que las abundancias de ocho de nueve especies con distribución sureña se incrementaron significativamente, mientras que las abundancias de cinco de ocho especies de afinidad norteña decrecieron de forma significativa entre 1931-1933 y 1993-1994, mismo periodo en el que se registró, en promedio, un aumento de la temperatura del mar.
Adicionalmente, uno de los componentes más relevantes de la comunidad, como lo es M. pyrifera, ha sido estudiado con una perspectiva de largo plazo en la parte sur de California y se concluyó que la biomasa de esta macroalga disminuyó en aproximadamente dos tercios desde 1957. Esta disminución se ha relacionado estrechamente con un aumento de la temperatura del mar y con la limitación concomitante de nutrientes claves para el desarrollo de esta especie (Tegner et al., 1996).
Actualmente H. fulgens es la especie que más contribuye a la pesquería en Bahía Asunción, con alrededor del 90%, mientras que H. corrugata aporta aproximadamente un 9% (León-Carballo y Muciño-Díaz, 1996). Sin embargo, H. corrugata fue el principal componente de la pesquería durante los inicios de ésta y fue reduciendo su aportación de manera continua y de forma más evidente a mediados de los años setenta (Ponce-Díaz et al., 2000). Por otro lado, en California recientemente se ha prohibido la captura del abulón ya que se observaron tendencias de disminución en sus poblaciones desde principios de los cincuenta para H. corrugata, y a mediados de los setenta para H. fulgens, hasta llegar al punto de cerrar la pesquería (Karpov et al., 2000). Esta situación indica que estas especies presentan una tolerancia diferencial a las condiciones del clima en la región; así, H. fulgens tiene una mayor adaptación a esta zona de transición que otras especies de abulones según lo demuestra su elevada participación en la proporción de las capturas en años recientes que se distinguen por ser más cálidos de lo normal, así como por su distribución más sureña pues la especie ocurre hasta los límites de Bahía Magdalena (Cox, 1962). No obstante, esta posibilidad de un efecto de largo plazo (por temperatura) en la reducción de las poblaciones de H. corrugata no elimina la potencial causa de su disminución del abulón amarillo por la pesca o una interacción de estos dos factores.
Por otra parte, el análisis de densidad espectral indica que la mayor contribución a la variabilidad de la serie es el componente estacional; conviene subrayar que el componente interanual es el segundo factor que explica la mayor variabilidad de los datos, lo cual se identifica seguramente con la ocurrencia de fenómenos tipo ENSO en la zona y el consecuente efecto de estos fenómenos sobre la comunidad en general. Ante la presencia de eventos tipo ENSO se ha sugerido la ocurrencia de cambios en el sentido de una reducción de ciertas poblaciones de pelágicos menores, reducción de alimento, cambios temporales en la estructura trófica de los sistemas, invasión de especies tropicales hacia zonas templadas e incluso cambios en la biodiversidad; sin embargo, todos estos cambios, deletéreos para ciertas especies, tienen un carácter episódico como el propio fenómeno de El Niño y su efecto en la comunidad biótica depende de su intensidad y duración (Guzmán-del Próo et al., en prensa).
En relación a los efectos de El Niño 1997-1998, para la costa occidental central de la Península de Baja California se ha reportado un efecto directo sobre la comunidad de los mantos de M. pyrifera, los cuales desaparecieron totalmente hacia finales del otoño (noviembre de 1997) y, en el caso de E. arborea, la especie quedó reducida a unos cuantos especímenes defoliados en la zona contigua a Bahía Tortugas (Guzmán-del Próo et al., en prensa). Sin embargo, para zonas más al sur, el efecto sobre E. arborea no fue tan drástico (Hernández-Carmona et al., 2001). Este efecto deletéreo sobre estas macroalgas es relevante por su condición de ser fuente de dietas naturales para el abulón (Guzmán-del Próo et al., 1991; Serviere-Zaragoza et al., 1998). Otro efecto directo observado sobre los componentes de la comunidad, producto de la variación interanual de la temperatura del mar, es la disminución en el reclutamiento de juveniles de abulón para la zona de Bahía Tortugas durante el fenómeno ENSO más intenso (Carreón-Palau, 2000; Guzmán-del Próo et al., en prensa). Todo lo anterior revela la importancia de la variación interanual detectada en el análisis espectral.
En el área de estudio es notoria la ausencia de información sobre aspectos del clima marino, particularmente acerca de mediciones finas o de alta frecuencia; la información disponible está asociada a datos de gran escala y de baja frecuencia, como son las series de temperatura superficial del mar de COADS o la serie Douglas. Por tanto, consideramos que este trabajo es una aportación al conocimiento de la variabilidad climática de la zona y que brindará más elementos cuantitativos para futuros análisis relacionados con los organismos que conforman las comunidades presentes en estos sistemas costeros altamente productivos.
Agradecimientos
Los autores agradecen a la Federación Regional de Cooperativas de Producción Pesquera Baja California, a las cooperativas de producción pesquera Leyes de Reforma y California de San Ignacio, al Programa Langosta del Centro Regional de Investigaciones Pesqueras de La Paz (INP), a Fernando López-Salas y a Gustavo Hernández-Carmona (CICIMAR-IPN) por su apoyo en las tareas de campo y/o por proporcionar parte de la información utilizada, al Laboratorio de Modelación y Pronóstico Pesquero del CIBNOR por la sistematización de los datos, y al Dr. Sergio A. Guzmán-del Próo (ENCB-IPN) por leer y comentar críticamente el manuscrito inicial. Este trabajo fue financiado parcialmente por el CIBNOR (Programa Ecología Pesquera RP-6) y el CONACYT (proyecto 35151B). GPD y JJBR son becarios CONACYT (registros 119827 y 163276, respectivamente). Finalmente se agradece a los cuatro revisores anónimos, quienes aportaron valiosas sugerencias e ideas que contribuyeron a mejorar este trabajo.
Referencias
Amador-Buenrosto, A., Argote-Espinoza, M.L., Mancilla-Peraza, M. y Figueroa-Rodríguez, M. (1995). Variaciones de período corto de la circulación anticiclónica en Bahía Sebastián Vizcaíno, B.C. Cien. Mar., 21(2): 201-223. [ Links ]
Barry, J.P., Baxter, C.H., Sagarin, R.D. and Gilman, S.E. (1995). Climate-related, long-term changes in a California rocky intertidial community. Science, 267: 672-675. [ Links ]
Belmar-Pérez, J. y Guzmán-del Próo, S.A. (1992). Madurez sexual y cliclo gonádico en Haliotis fulgens y Astraea undosa en Bahía Tortugas. En: S.A. Guzmán-del Próo (ed.), Taller México-Australia sobre reclutamiento de recursos bentónicos de Baja California. SEPESCA / IPN, pp. 121-129. [ Links ]
Breen, P.A. (1992). A review of models used for stock assessment in abalone fisheries. In: S.A. Shepherd, J.M. Tegner and S.A. Guzmán-del Próo (eds.), Abalone of the World: Biology, Fisheries and Culture. Fishing New Books, Cambridge, UK, 607 pp. [ Links ]
Carreón-Palau, L. (2000). Microhabitat, distribución y abundancia de juveniles de abulón, Haliotis fulgens y H. corrugata en Bahía Tortugas, B.C.S., México. Tesis de maestría, Escuela Nacional de Ciencias Biológicas, Instituto Politécnico Nacional, 79 pp. [ Links ]
Cox, K.W. (1962). Californian abalones, family Haliotidae. Calif. Fish Game, 118: 1-133. [ Links ]
Douglas, A.V. (1976). Past air-sea interactions over the eastern North Pacific Ocean as revealed by tree-ring data. Department of Geosicences, Tucson, University of Arizona, 196 pp. [ Links ]
Douglas, A.V. (1980). Geophysical estimates of sea-surface temperatures off North America. CalCOFI Rep., 21: 102-112. [ Links ]
Guzmán-del Próo, S.A., Mille-Pagaza, S.R., Guadarrama-Granados, R., De la Campa-de Guzmán, S., Carrillo-Laguna, J., Pereira-Corona, A., Belmar-Pérez, J., Parra-Alcocer, M. de J. y Luque-Guerrero, A.C. (1991). La comunidad bentónica de los bancos de abulón (Haliotis spp. Mollusca: gasteropoda) en Bahía Tortugas, Baja California Sur, México. Anales de la Escuela Nacional de Ciencias Biólogicas, (36): 27-59. [ Links ]
Guzmán-del Próo, S.A. (1992). A review of the biology of abalone and its fishery in Mexico. In: S.A. Shepherd, J.M. Tegner and S.A. Guzmán-del Próo (eds.). Abalone of the World: Biology, Fisheries and Culture. Fishing New Books, Cambridge, UK, 607 pp. [ Links ]
Guzmán-del Próo, S.A. (1994). Biología, ecología y dinámica de población de abulón (Haliotis spp.) de Baja California, México. Tesis doctoral, Escuela Nacional de Ciencias Biológicas, Instituto Politécnico Nacional, 188 p. [ Links ]
Guzmán-del Próo, S.A., Carreón-Palau, L., Belmar-Pérez, J., Carrillo-Laguna, J. y Herrera-Fregoso, R. Efectos del fenómeno de "El Niño" sobre el reclutamiento de invertebrados bentónicos de Baja California. Geofís. Int. (en prensa).
Hernández-Carmona, G., García, O., Robledo, D. and Foster, M. (2000) . Restoration techniques for Macrocystis pyrifera (Phaeophyceae) populations at the southern limit of their distribution in Mexico. Bot. Mar., (43): 273-284. [ Links ]
Hernández-Carmona, G., Robledo, D. and Serviere-Zaragoza, E. (2001) . Effect of nutrient availability on Macrocystis pyrifera recruitment and survival near its southern limit off Baja California. Bot. Mar., (44): 221-229. [ Links ]
Hewitt, R. (1981). Eddies and speciation in the California Current. CalCOFI Rep., 22: 96-98 p. [ Links ]
INP (2000). Pesquería de abulón. En: Sustentabilidad y Pesca Responsable en México. Evaluación y Manejo 1999-2000. Instituto Nacional de la Pesca. CD-Rom. [ Links ]
Karpov, K.A., Haaker, P.L., Taniguchi, I.K. and Rogers-Bennett, L. (2000). Serial depletion and the collapse of the California abalone (Haliotis spp.) fishery. Workshop on rebuilding abalone stocks in British Columbia. Can. Spec. Publ. Fish. Aquat. Sci., 130: 11-24. [ Links ]
Leighton, D.L. (1974). The influence of water temperature on larval and juvenile growth in three species of southern Californian abalones. Fish. Bull., (72): 1137-1145. [ Links ]
León-Carballo, G. y Muciño-Díaz, M. (1996). Pesquería de abulón. En: M. Casas-Valdéz y G. Ponce-Díaz (eds.), Estudio del Potencial Pesquero y Acuícola de Baja California Sur. SEMARNAP, Gob. BCS, FAO, INP, UABCS, CIBNOR, CICIMAR, CETmar, pp. 15-41. [ Links ]
Lluch-Belda, D., Lluch-Cota, D.B., Lluch-Cota, S.E. y Hernández-Vázquez, S. (1997). Análisis de la pesquería de abulón de la península de Baja California. Informe Técnico, CIBNOR, 19 pp. [ Links ]
Lluch-Belda, D., Hernández-Rivas, M., Saldierna-Martínez, R. y Guerrero-Caballero, R. (2000). Variabilidad de la temperatura superficial del mar en Bahía Magdalena, B.C.S. Oceánides, 15(1): 1-23. [ Links ]
Lluch-Belda, D., Laurs, R.M., Lluch-Cota, D.B. and Lluch-Cota, S.E. (2001). Long-term trends of interannual variability in the California Current System. CalCOFI Rep., 42: 129-144. [ Links ]
Muñoz, L.T. (1976). Los bancos abuloneros de la parte central de Baja California. Memorias del Primer Symposium sobre recursos pesqueros Masivos de México. Vol. Esp. sobre Abulón/Langosta. INP 1: 161-184. [ Links ]
Ponce-Díaz, G., Vega-Velázquez, A., Ramade-Villanueva, M., León-Carballo, G. and Franco-Santiago, R. (1998). Socioeconomic characteristics of the abalone fishery along the west coast of Baja California Peninsula, Mexico. J. Shellfish Res., 17(3): 853-857. [ Links ]
Ponce-Díaz, G., Chávez, E.A. y Ramade-Villanueva, M. (2000). Evaluación de la pesquería de abulón azul Haliotis fulgens en Bahía Asunción, Baja California Sur, México. Ciencias Marinas, 26(3): 393-412. [ Links ]
Ponce-Díaz, G., López-Salas, F., Hernández-Carmona, G., Lluch-Cota, S., Bautista-Romero, J. y Lluch-Belda, D. (2003). Base de datos de la temperatura del mar en Bahía Asunción, Baja California Sur, México. Instituto de Investigaciones Oceanológicas/Universidad Autónoma de Baja California, México, Reporte Técnico 2003-01, 16 pp. (Disponible on-line, formato pdf, en: http://rcmarinas.ens.uabc.mx/~cmarinas/reportes/reportecnico012003.pdf). [ Links ]
Prince, J.D. and Guzman del Próo, S.A. (1993). A stock reduction analysis of Mexican abalone (Haliotid) fishery. Fish. Res., (16): 25-49. [ Links ]
Rodríguez-Valencia, J.A. and Caballero-Alegría, F. (2002). Temporal fluctuations (1989-1999) in the populations of Haliotis fulgens and H. corrugata (Gastropoda: Haliotidae), at Isla San Benito (Baja California, Mexico). J. Shellfish Res., 21(1): 163-171. [ Links ]
Serviere-Zaragoza, E., Gómez-López, D. and Ponce-Díaz, G. (1998). The natural diet of the green abalone (Haliotis fulgens Philippi) in the southern part of its range, Baja California Sur, Mexico, assessed by an analysis of gut contents. J. Shellfish Res., 17(3): 777-782. [ Links ]
Shepherd, S.A., Turrubiates, J.R. and Hall, K. (1998). Decline of the abalone fishery at La Natividad, Mexico: Overfishing or climate change? J. Shellfish Res., 17(3): 839-846. [ Links ]
Tegner, M.J., Dayton, P., Edwards, P.B. and Riser, K.L. (1996). Is there evidence for long-term climatic change in southern California kelp forest? CalCOFI Rep., 37: 111-126. [ Links ]
Vega, A., Lluch-Belda, D., Muciño, M., León, G., Hernández, S., Lluch-Cota, D., Ramade, M. and Espinoza, G. (1995). Development, perspectives and management of lobster and abalone fisheries off northwest Mexico, under a limit access system. Proc. 2nd World Fisheries Congress. Collingwood, VIC, Australia, 797 pp. [ Links ]
Vega-Velázquez, A., Espinosa-Castro, G. y Gómez-Rojo, C. (1996). Pesquería de langosta Panulirus spp. En: M. Casas-Valdez y G. Ponce-Díaz (eds.), Estudio del Potencial Pesquero y Acuícola de Baja California Sur. SEMARNAP, Gob. BCS, FAO, INP, UABCS, CIBNOR, CICIMAR, CETmar, pp. 227-261. [ Links ]
Walsh, J.J., Kelly, J.C., Whitehead, T.E., MacIssac, J.J. and Huntsman, S.A. (1974). Spin-up of the Baja California upwelling ecosystem. Limnol. Oceanogr., 19(4): 553-573. [ Links ]
Walsh, J.J., Whitehead, T.E. Kelly, J.C., Huntsman, S.A. and Pillsbury, R.D. (1977). Further transition states of the Baja California upwelling ecosystem. Limnol. Oceanogr., 22(2): 264-280. [ Links ]
Woodruff, S.D., Lubker, S.J., Wolter, K., Worley, S.J. and Elms, J.D. (1993). Comprehensive Ocean-Atmosphere Data Set (COADS) Release 1a: 1980-92. Earth System Monitor, 4(1): 1-8. (http://www.cdc.noaa.gov/coads/coads1a.html). [ Links ]

