











 text in
text in  English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO 
 Permalink
PermalinkThe genus Opuntia is a natural resource that is used as live fencing and is also cultivated on commercial plantations as a source of fodder and as an edible vegetable for the nutritional value of cladodes and fruits (Griffith, 2004; Defelice, 2004; Sáenz et al., 2006). It is a crop with high demand as a food worldwide, and it is exported to the U.S., Europe and Japan (Flores et al., 2013). Among the species of the genus Opuntia, we find a set of wild cultivars generically known as xoconostles or acidic cactus pears, the name deriving from the Náhuatl (xoco = sour and nochtli = prickly pear) (Gallegos-Vázquez et al., 2012). These species are considered of high ecological adaptability and their anatomy and morphology enable them to adapt to conditions of environmental stress, being considered valuable vegetable foods in Latin America. Xoconostle cultivars possess an appreciable amount of soluble fiber and antioxidant compounds such as ascorbic acid, while the seeds are a source of fiber, phenolics, flavonoids, polyunsaturated fatty acids, and tocopherols (especially γ-tocopherol), which provide good antioxidant capacity (Morales et al., 2012), as well as high-quality bioactive compounds that could be incorporated into functional food formulation (Morales et al., 2014; Morales et al., 2015). This renders the plant an ideal crop in zones where the growth of other species-of-agricultural-interest is a challenge (Sáenz et al., 2006).
In common with O. ficus-indica and O. monacantha, the xoconostle species also present phytosanitary problems caused by different microorganisms that affect the quality and yield of the crop (Rodríguez et al., 2001; Choueiri et al., 2005). Among the most common of these problems in Opuntia we find golden spot (Quezada et al., 2006), necrosis of the cladode (Swart and Kriel 2002), and golden spot produced by Alternaria alternata (Granata and Sidoti, 1997; Swart and Kriel, 2002). The symptoms of the latter disease have been described as an infection that begins in the areoles of the cladode, near the site of penetration, where a slight bump appears, with the tissue changing color from the original dark to light green until becoming golden (Méndez et al., 2008).
Similar symptomatology to that of golden spots in O. ficus-indica has recently been observed on both faces of the cladodes of xoconostle (Personal observation). However, to our knowledge, there have been no reports of this disease in xoconostle. Therefore, the aim of this study was to perform a comparative analysis of the disease symptomatology in xoconostle with those reported in other Opuntia species and to identify the microorganism that cause this symptomatology.
The study was conducted during the years 2017-2018 in Chapantongo (2017-2018) in Hidalgo, Mexico (20º 17’ N and 99º 24’ W, at 2,120 meters above sea level) in an agricultural area established in Phaeozem and Vertisol soil types. The climate is temperate-subhumid with an annual average precipitation of 550 mm and an average annual temperature of 18 °C. The study sites feature a north-south slope of 7% (INEGI, 2009), the sites were previously occupied by pastureland, and present an arable layer of up to 20 cm in depth.
Field sampling. Simple random sampling was performed from a field crop established in 10 plots on one-hectare with 13 rows containing 1,250 O. matudae cv. Rosa plants. A total of 200 plants were sampled with symptoms that resembled those of the disease described as golden spot in O. ficus-indica were found. Of these, 100 samples were randomly selected for isolation and purification of the pathogen causing the disease. The symptoms on the cladodes of the infected plants of O. ficus-indica are chlorotic spots around the spines. The spots develop into round and prominent golden yellow scabs, then the center necrotized leaving a yellowish halo. The disease caused by A. alternata in O. ficus-indica is also seen on the fruits, which often presents the same symptoms observed on the pads (Granata and Sidoti, 1997). Cladodes of O. matudae cv. Rosa plants presenting symptoms of the disease were collected, wrapped in sterile paper, and stored at 4 ºC until their analysis. Plant samples were identified and deposited in the culture collection in the Laboratory of Phytopathology of the Xochimilco Metropolitan Autonomous University.
Isolation and purification of fungus. The plant material was washed with potable water and 2 cm2 of tissue were disinfected during 3 min with a solution of 5% of sodium hypochlorite and 50% alcohol. After washed three times with sterile distilled water they were placed on sterile paper to remove residual water, and then 0.5 cm2 fragments were placed onto fifty plates containing Potato Dextrose Agar (PDA) and fifty containing Water Agar (WA). They were randomly placed and incubated at 28 ± 2 °C for five days. The fungi were identified using specific keys (Simmons, 2008). The same amount was used during purification.
The strains identified as Alternaria sp. were isolated with the hyphal tip technique in WA media and PDA and incubated at 28 ± 2 °C until sporulation was confirmed. The spores were collected, placed in an Eppendorf tube with 1 mL of Tween 20 solution (0.5%), and then stirred for 1 min. The samples were centrifuged for 30 s and then the supernatant was removed. Sterile distilled water was added to resuspend the spores. From this spore suspension 10-4 dilutions were prepared, 0.5 mL was placed on PDA and WA plates, and were incubated at 28 ± 2 oC. After 24 h a fragment of the culture medium containing germinated spores was cut out and placed on PDA and WA plates and incubated at 28 ± 2 °C until sporulation (Montiel et al., 2016). Fragments of monosporic cultures in Rose Bengal Agar culture media, Czapek Dox-agar and Sabouraud Agar plates were incubated at 28 ± 2 °C for 24 h. The spores were identified morphologically with taxonomic keys (Simmons, 2008).
Morphological and molecular identification. A fragment of two pure Alternaria colony were placed in PDA medium and incubated at 28 °C until maximum development; the mycelium developed was macerated with liquid nitrogen until a fine powder was obtained. From this colony genomic DNA extraction was performed with 0.5 g of powdered mycelium (Shure et al., 1983), verification was performed with the technique of electrophoresis in 0.8% agarose gel, stained with ethidium bromide and the concentration was determined with the NanoDrop 1000 spectrophotometer. The fungal DNA extracted was used for the amplification of the ribosomal RNA 18S gene.
The fungal DNA extracted was used for the amplification of the ribosomal RNA 18S gene using the oligonucleotides NS5 / NS6, and the intergenic region ITS1 (ITS5 / ITS2) and ITS2 (ITS5 / ITS4) of the genes that encode for ribosomal RNAs 18S, 5.8S and 28S, respectively (White et al., 1990). The amplification of the fragments (ITS1 was a length of 234 bp and ITS2 was a length of 565 bp) was done with the PCR technique (30 μL of a reaction mixture: JumpStart ™ Reaction mix 2x, 50 ng of fungal gDNA and 10 pM μL-1 of oligonucleotides). The mixture was placed in the TC-5000 Techne® thermal cycler according to the program (denaturalization at 94 °C, 160/35 s, 35 cycles; alignment at 55 °C, 35 cycles; extension at 72 °C, 45 s, 35 cycles and final extension at 72 °C, 300 s 35 cycles) (White et al., 1990). The PCR products were separated by electrophoresis in agarose gel at 0.8% and purified with the Sigma® GenElute Gel extraction kit. The purified PCR products were bound in pGEM T-Easy vector (Promega), transformed into competent cells of Escherichia coli strain DH5α, re-sowed in liquid medium LB with ampicillin, incubated at 37 ºC with constant movement during 24 h. From the obtained colonies the plasmid DNA was purified with Illustra plasmidPrep Mini Spin Kit of GE Healthcare and the digestion was done with the restriction enzymes EcoRI and PstI; the digestion was separated by electrophoresis in agarose gel at 0.8% with ethidium bromide, photo-documented on the BIO-RAD Fluor-S® Max Multi Imager and quantified on the NanoDrop 1000 spectrophotometer. The sequences were performed in the DNA Synthesis and Sequencing Unit of the Biotechnology Institute of UNAM. The obtained sequences were edited on FinchTV and identified as Altal-ITS1 and Altal-ITS2. To determine their species, the BLAST tool was used in the National Center Biotechnology Information (NCBI) database (http://www.ncbi.nlm.nih.gov/BLAST). To determine the sequence identity of the Altal-ITS1 and Altal-ITS2 (234 and 565 bp respectively), the isolates used were Alternaria alternata isolate wxm90 (GenBank: HM051066.1); Alternaria sp. ZH7-C2 (GenBank: FJ037737.1); Uncultured fungus clone AA45-2 (GenBank: GU065589.1); Septoria lycopersici isolate NW574B (GenBank: EU520171.1); Alternaria sp. ALT3 (GenBank: HM114272.1); Alternaria alternata strain KHF-5 (GenBank: MN173818.1); Alternaria alternata isolate Altal-ITS1 (GenBank: MW092952.1); Altal-ITS2 (GenBank: MW092953); Alternaria sp. JM-1 (GenBank: GQ389617.1); Alternaria azukiae strain wxm133 (GenBank: HM037984.1); Alternaria alternata (GenBank: LC269969.1) and Alternaria alternata isolate R1-1 (GenBank: FJ827039.1). The Altal-ITS1 and Altal-ITS2 sequence was aligned with the previous sequences and the phylogenetic tree was constructed using Unipro UGENE v. 41.0 (Okonechnikov et al., 2012).
Pathogenicity test. Field evaluation was carried out under greenhouse conditions, in which mature cladodes of xoconostle (O. matudae cv. Rosa) were obtained from healthy plants. Prior to performing the seeding, twenty cladodes were washed with sterilized water and disinfected with a solution of 5% sodium hypochlorite; these cladodes were left in the shade during 20 days for scarring of the cut. Later, they were established in 35 x 35 cm plastic bags filled with a mixture of tezontle (volcanic rock) and silica sand (2:1).
Prior to the inoculation carried out at the greenhouse, activation of one isolated was performed separately in 250 mL flasks in Potato Dextrose Agar (PDA) culture medium. After 8 days of incubation at 28 ± 2 °C, an additional 20 mL of Tween 80 at 0.05%, was added to each flask and these were placed under orbital shaking during 1 min. The solution obtained from the shaking was passed through a Whatman #40 filter and the filtrate was diluted with 130 mL sterile water, which was applied to the cladodes of the xoconostle plants. The cladodes inoculated by aspersion of 13 mL of a solution and a control (water) were constituted in the treatments. Each treatment had 10 repetitions (cladodes) distributed in an arrangement completely at random. Every 15 days, observations were performed in the search for the appearance of symptoms similar to those of golden spot, noting the treatment and the number of plants in cases in which the symptoms appeared. In case of the appearance of the symptomatology similar to those of golden spot 0.5 cm2 fragments of the cladodes were disinfected in the 2% sodium hypochlorite solution and seeded onto PDA plates and incubated at 28 ± 2 °C. After 8 days of growth, the colonies were identified morphologically with taxonomic keys (Simmons, 2008).
Statistical analysis. Data were submitted to tests of normality (Shapiro-Wilk W test) and homoscedasticity (Levene). Kruskal-Wallis Analysis of Variance (ANOVA) and means comparison (Steel-Dwass) tests were used when the supposed normality could not be corrected using transformations. ANOVA and the Tukey test (p <0.05) were used to separate treatments. Data analysis and processing was carried out with JMP® ver. 11 statistical software packages (SAS, 2012).
Field Sampling. The presence of symptoms was observed in 100% sampled plants of the xoconostle crop. The disease presented generally in the cladode on the upper edge or on that prior to fruting cladode (Figure 1A, B). This begins in the cladodes as points of yellow color or spot chlorosis that often presented near the areoles (Figures 1C, D), similar to those caused by A. alternata and reported by Granata and Sidoti (1997) on cactus pear, and these evolved until they formed a golden spot and necrosis on the epidermis (Figures 1E, F). Subsequently, these acquired the appearance of burns as a result of the fusion of the spots, with elevation of the cuticle, dehydration of the tissue, and crusting. Similar symptomatology has been reported for Opuntia ficus (Swart and Kriel, 2002; Granata and Sidoti, 1997). The symptoms found in the xoconostle plant in this study did not correspond to those described by Ammar et al. (2004) and Flores et al. (2013) produced by Botryodiplodia theobromae, Alternaria alternata and Fusarium solani on cladodes and fruit rots of prickly pear cactus, nor to those described by Méndez et al. (2008) for golden spot caused by Alternaria sp., Hansfordia sp., Ascochyta sp. in O. ficus-indica. These authors state that the damage presents in the fruiting cladode and that it affects the entire area of the cladode in this species, unlike in the xoconostle, where the damage is observed in the cladode prior to the apical cladode and that does not present in the fruits (Figure 1A, B).

Figure 1 Expression of golden spot in Opuntia matudae, var. Rosa in plants sampled in the field. The symptomology generally presented on the upper edge of cladode (A); and on that prior to fruting cladode (B); golden spot on the epithelial tissue (C); presence of golden spot in the areole and thickening of the epithelial tissue (D); thickening of the tissue caused by fusion of the golden spots, mainly on the upper edge of the cladode (E) and concentric lesion in cladode tissue and necrosis on the epidermis (F).
The lesions reported by Swart and Kriel (2002) for golden spot in the cladodes of O. ficus-indica present a similar pattern, since these authors describe chlorosis around the spines, which was observed in our study; however, the presence of exudates in the lesions, reported by Ammar et al. (2004), was not found.
Morphological and molecular identification. Purification allowed for the elimination of saprophytic organisms. From a 100 samples A. alternata was identified in 100% of the incubated plates. The distinctive feature of the colony in PDA culture medium was greenish-grey plush, concentric with whitish rim, dark brown plate underside. The conidiophores were 3 to 6 µm thick and 50 µm long on average The conidia were generally short in chains of 3 to 5 conidia, secondary conidiophores were also observed, short, straight, ovoid, with short cylindrical beak, 7-25 µm long, 5-12 µm wide, olive colored, with 3 transverse septa and 2 longitudinal septa. According to McKenzie (2013), the morphology corresponds to A. alternata (Figure 2C-D).
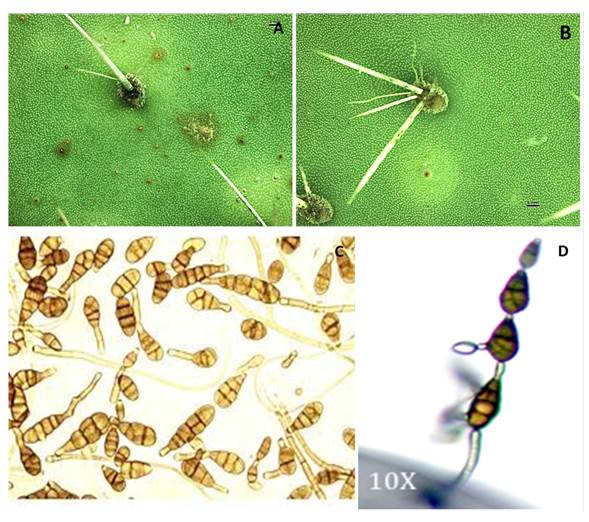
Figure 2 Symptoms caused by the inoculation of Alternaria alternata in cladodes of xoconostle under greenhouse conditions: without the presence of the chlorotic halo (A); with the presence of the chlorotic halo (B); A. alternata colony sporulation pattern in short (C); branched chain in the second conidium (D).
The Altal-ITS1 (length of 234 bp NCBI Access num: MW092952) and Altal-ITS2 sequence (length of 565 bp NCBI Acces num: MW092953) showed a homology percentage (98%). According to alignments in the National Center Biotechnology Information database and the previous morphological characterization, allowed the identification of Alternaria alternata (Figure 3).
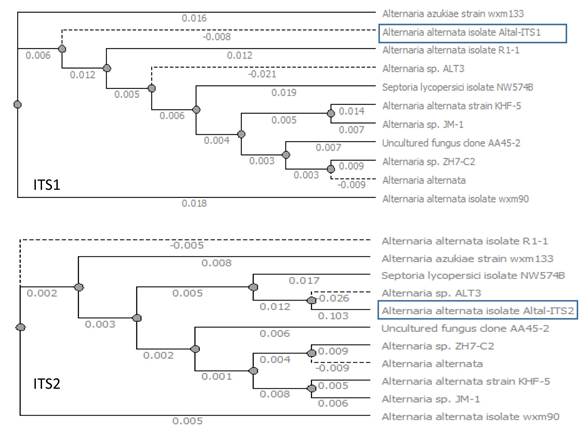
Figure 3 Phylogenetic tree construction comparing the Altal-ITS1 and Altal-ITS2 amplified regions in Alternaria alternate.
Greenhouse experiment. Ten months after inoculation carried out at the greenhouse, the appearance of symptoms similar to those caused by golden spot in O. ficus-indica began. A significant difference (P≤0.05) was observed among the treatments evaluated, finding that those in which the solution containing the strain, were those that presented experimental units with symptomatology, while the control treatment did not present these. In cladodes inoculated with solution containing the isolated were identified areas in brown color with a diameter of between 1 and 5 mm and an elevation of the tissue; but chlorotic halos were not registered (Figure 2A). However, there were also cladodes in which the chlorotic halo was present (Figure 2B). The microorganisms identified from the isolates corresponded to Alternaria alternata, observing the presence of their conidia in the spines of the cladodes.
Morphological observations allowed to identify A. alternata as the causal agent of the Golden spot in O. matudae and the symptoms are similar to that of the golden spot in O. ficus-indica. The molecular characterizations of the Altal-ITS1 and Altal-ITS2 isolation coincided with the A. alternata species. More research is needed to investigate the effect of the integrated management and specially of fungicides on the growth of the pathogenic fungi A. alternata in xoconostle.

