











 text new page (beta)
text new page (beta) Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO 
 Permalink
PermalinkINTRODUCCIÓN
En comparación con los demás órdenes de Hexapoda, los mántidos se pueden considerar escasos, ya que hay sólo 2,494 especies válidas registradas (Wieland & Svenson, 2018) de más de un millón de especies de hexápodos en el mundo (Giribert & Edgecombe, 2013; Gullan & Cranston, 2014). Este reducido número de especies es un patrón generalizado que se ha documentado en diferentes partes del mundo (Menezes & Bravo, 2014; Ferreira-Ribeiro et al., 2009; Ortega & Márquez, 1987; Rehn, 1904). El conocimiento de los mántidos a nivel nacional es escaso, se tienen registradas 72 especies para México y sólo 13 en Chiapas, éstas pertenecen a los siguientes siete géneros: Hondurantemna, Liturgusa, Melliera, Pseudomiopteryx, Stagmomantis, Vates y Zoolea (Hernández-Baltazar & Gómez, 2017; Hernández-Baltazar et al., 2018; Jantsch, 1999; Rehn, 1935a, 1935b; Rodrigues et al., 2017; Roy, 2012; Svenson, 2014; Terra, 1995).
Existen relativamente pocos estudios sobre estos artrópodos a nivel sistemático, por lo que su estudio es imprescindible para conocer más de ellos y sus relaciones con otros taxa (Agudelo et al., 2007; Ariza et al., 2012; Patel & Singh, 2016; Rivera, 2010).
Tomando en cuenta esta información, el objetivo de este trabajo fue analizar la diversidad de mántidos en cuatro localidades de la Reserva de la Biósfera Selva El Ocote. Esta región forma parte del Corredor Biológico Mesoamericano, considerado de gran importancia en México y el mundo, por su alto número de especies y endemismos (Comisión Nacional de Áreas Naturales Protegidas, CONANP 2001). No obstante, esta alta biodiversidad es poco conocida y vulnerable a procesos de transformación de su hábitat, principalmente por deforestación (Flamenco-Sandoval et al., 2007).
MATERIALES Y MÉTODOS
Área de estudio. La Reserva de la Biósfera Selva El Ocote (REBISO), se localiza en el estado de Chiapas, entre los municipios de Ocozocoautla de Espinoza, Cintalapa de Figueroa, Tecpatán de Mezcalapa y Jiquipilas, situada entre los 16° 45’ 42” y 17° 09’ 00” latitud norte y 93° 54’ 19” y 93° 21’ 20” longitud oeste, abarcando una superficie de 101,288 ha (Fig. 1).
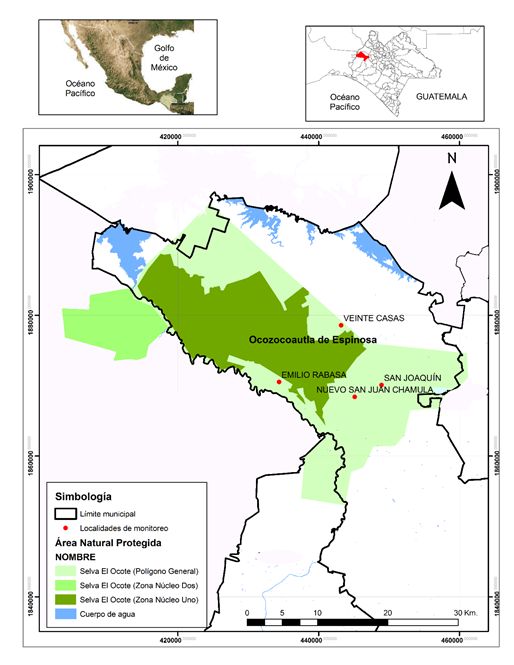
Figura 1 Localización geográfica de la Reserva de la Biósfera Selva El Ocote (REBISO), Chiapas, México.
El trabajo de campo se realizó en cuatro localidades de la REBISO, considerándose de acuerdo con Gómez et al. (2017) dos sitios como áreas de conservación con selva mediana subperennifolia (Emilio Rabasa y Veinte Casas), y dos como sitios perturbados con vegetación alterada (Nuevo San Juan Chamula y San Joaquín El Progreso). Las localidades seleccionadas se encuentran en la zona de amortiguamiento y limitan con la zona núcleo de la Reserva (Fig. 1).
Recolecta de individuos. Se realizaron ocho viajes de recolecta de cinco días cada uno, cuatro durante la temporada sin precipitación (febrero-mayo, 2016) y cuatro con precipitación (junio-agosto, 2016), tomando en cuenta la fluctuación de la precipitación pluvial de los últimos seis años (Comisión Nacional del Agua, CONAGUA 2011).
En cada localidad y siguiendo la metodología de Arteaga et al. (2014), se realizó un transecto de 1 km de largo y 10 m de ancho, en éste se realizaron diez puntos de muestreo a 100 m de distancia entre cada uno. En cada sitio de recolecta se realizaron dos métodos de captura de mántidos: a) revisión y recolección manual y b) trampas de luz (Agudelo & Chica, 2003). En cada toma de muestras se revisó, durante el día, la vegetación, especialmente las hojas (haz y envés) y troncos. Por la noche, se instalaron dos trampas de luz blanca y negra de 20:00 a 24:00 horas y separadas entre sí 200 metros (Mariño-Pedraza, 2011; Márquez-Luna, 2005).
Los organismos recolectados se colocaron en viales con alcohol al 70% (Arteaga et al., 2014; Márquez-Luna, 2005) y se transportaron al laboratorio para su posterior identificación. La determinación taxonómica se realizó mediante el análisis de la genitalia (Ariza et al., 2012; Brannoch et al., 2017; Cerda, 1993; Jensen et al., 2009; Klass, 1997; Rehn, 1935a; Roy, 2004) y empleando claves del grupo (Battiston & Picciau, 2008; Rafael et al., 2012; Romero & Nelson, 2008; Giglio-Tos, 1927; Rehn, 1935a; Roy, 2004; Saussure & Zehntner, 1894). La clasificación de las especies se ubicó siguiendo las propuestas de Rivera & Svenson (2016) y Wieland & Schütte (2017). El material se depositó en la colección de El Colegio de La Frontera Sur, Unidad San Cristóbal (ECO-SC-E).
Análisis de datos. Para la estimación de la diversidad alfa por localidad se utilizó la riqueza específica (S), y el número de individuos por especie (N) por cada localidad (Halffter & Moreno, 2005; Whittaker, 1972). Para estimar la representatividad de los muestreos y establecer comparaciones a nivel de riqueza, diversidad y especies dominantes en cada una de las localidades se usaron los números de Hill: Q0=S, dónde: S= Número de especies; Q1=e H' , donde H'= Índice de Shannon-Wiener (calculado con logaritmos naturales) y Q2=1⁄D St , donde D St = Índice de Simpson (Jost, 2006; Moreno, 2001; Moreno et al., 2011). También se realizaron curvas de rango-abundancia considerando las abundancias relativas por especie (número de individuos en un área) en log10, con lo cual se obtuvo la distribución de las especies (Rocchini & Neteler, 2012).
Estimación de la eficiencia de captura. Con el fin de conocer la representatividad del inventario biológico, se efectuaron tres curvas de acumulación, la primera de forma general contemplando a las cuatro localidades y dos por cada temporada (con precipitación y sin ella). Para el esfuerzo de muestreo se consideró el número de eventos de muestreo (16 días de colecta) por el número de individuos. Dicho análisis se elaboró en el programa Estimates ver. 9.1.0 (Colwell, 2016). Para la construcción de las curvas se aleatorizaron los datos 100 veces, con el fin de reducir la variación en la forma de las curvas, debido al orden en que se consideran las diferentes muestras y por los sesgos temporales o espaciales en la distribución del esfuerzo de muestreo en cada sitio (Colwell & Coddington, 1994; Gotelli & Colwell, 2001; Jiménez-Valverde & Hortal, 2003).
Los datos se ajustaron al estimador Chao 1, el cual indica el número de especies esperadas, considerando la relación entre el número de especies representadas por un individuo y el número de especies representadas por dos individuos en las muestras (Colwell & Coddington, 1994; Moreno, 2001). Para evaluar la calidad de muestreo se procedió a usar la ecuación de Clench, ocupando para este análisis el programa Statistica ver. 13.2 (Jiménez-Valverde & Hortal, 2003; Moreno, 2001).
Diversidad beta (β). Para conocer el grado de cambio o reemplazo en la composición de especies, entre las comunidades que se encuentran en un área mayor (Halffter & Moreno, 2005), se emplearon los índices de Sorensen, Morisita y Morisita Horn, basados en los números de Hill. La diversidad beta toma valores de 1.0 y 2.0, por lo que valores mayores a uno indican que las comunidades estudiadas son completamente distintas en composición (Jost, 2006, 2007; Jost et al., 2011). Estos análisis se realizaron con ayuda del programa R ver. 3.0.0 y PAST ver. 2.17
Diversidad beta (β) verdadera. Ésta se mide como el número efectivo de comunidades o unidades de muestreo diferentes en la región, que corresponden al número de comunidades “virtuales” que podrían encontrarse en la región. Ésta se obtuvo mediante la división del número efectivo total de especies en un conjunto de datos entre el número efectivo de especies promedio de las muestras (Jost, 2007). Además, se obtuvo un dendrograma con base al índice de similitud de Jaccard (Moreno, 2001) para comparar los grupos formados con el valor de β verdadera.
Distribución estacional. Se analizaron los datos del inventario, correlacionándolos con los registros de temperatura y precipitación. Se elaboraron figuras con base en la estacionalidad de las especies obtenidas en la REBISO y se determinó la distribución estacional de los mántidos en cada mes de muestreo (Morón & Terrón, 1984).
RESULTADOS
Se capturó un total de 38 ejemplares de mántidos, distribuidos en tres familias, cinco géneros y siete especies (Cuadro 1). La familia que presentó la mayor riqueza de especies fue Mantidae (Fig. 2A) con el 71% de las especies. Stagmomantis montana montana fue la especie más abundante con el 26.31% del total (Fig. 2B).
Cuadro 1 Listado de mántidos presentes en la REBISO (* ya reportada en Chiapas).
| Familia | Género | Especie |
|---|---|---|
| Mantidae | Stagmomantis |
S. vicina* Saussure, 1870 S. nahua Saussure, 1869 S. montana montana* Saussure & Zehntner, 1894 |
| Melliera | M. major Saussure, 1872 | |
| Choeradodis | C. rhombicollis Latreille, 1833 | |
| Thespidae | Pseudomiopteryxs | P. infuscata* Saussure & Zehntner, 1894 |
| Mantoididae | Mantoida | M. maya Saussure & Zehntner, 1894 |
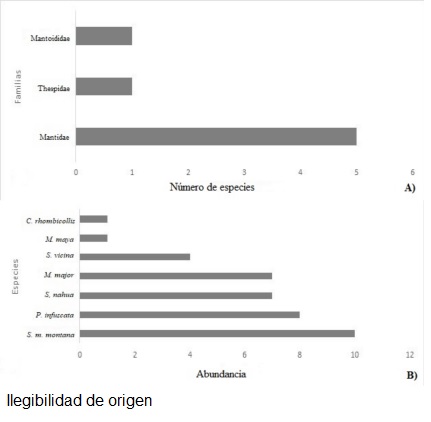
Figura 2 Riqueza y abundancia en la REBISO. A) Riqueza de familias presentes, B) Abundancia de especies de mántidos.
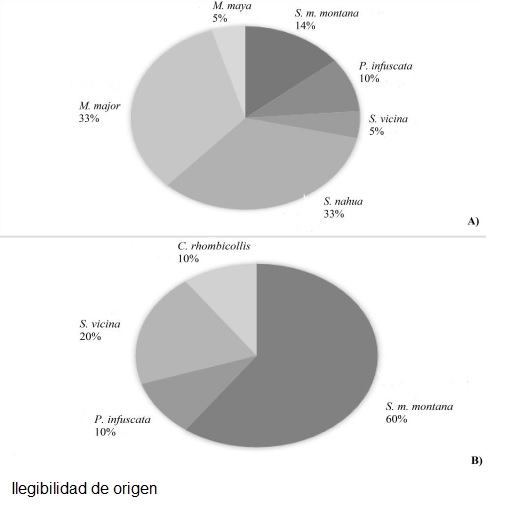
Riqueza y abundancia por localidades. La localidad de Emilio Rabasa (Fig. 3A) presentó la mayor riqueza de especies y abundancia en comparación con las otras localidades, de esta área se recolectaron seis especies y 21 ejemplares; seguida de Veinte Casas (Fig. 3B), sitio en el que se capturaron cuatro especies y diez individuos. De Nuevo San Juan Chamula se capturaron dos especies y seis individuos y S. m. montana fue la única especie encontrada (un individuo) en la localidad de San Joaquín El Progreso.
Con base en el número de Hill de orden cero, sin ponderar la abundancia de las especies, se encontró que los sitios de conservación (Emilio Rabasa y Veinte Casas) presentaron mayor riqueza de especies (Q0=7), mientras que los sitios perturbados (Nuevo San Juan Chamula y San Joaquín El Progreso) reflejaron menor riqueza específica (Q0=3). Para las especies típicas se obtuvieron cinco (Q1) y una especie dominante (Q2) para las localidades de conservación, mientras que las localidades perturbadas presentaron dos especies típicas (Q1) y dos dominantes (Q2) (Cuadro 2).
Cuadro 2 Números de Hill con base en los sitios conservados y perturbados de la REBISO.
| Localidades | Q0 | Q1 | Q2 |
|---|---|---|---|
| Emilio Rabasa y Veinte Casas | 7 | 5.50 | 1.26 |
| Nuevo San Juan Chamula y San Joaquín | 3 | 2.21 | 2.22 |
Por último, la curva de rango-abundancia indicó que la especie dominante para los sitios de conservación fue S. m. montana, para los sitios perturbados fueron Pseudomiopteryx infuscata, Stagmomantis nahua y Melliera major.
Eficiencia de muestreo. La representatividad del esfuerzo de captura mediante el estimador Chao 1, indicó que en la REBISO existen ocho especies, por lo que el inventario realizado para la zona de estudio está casi completo (88% de la fauna de mántidos que potencialmente se puede encontrar en la Reserva). Asimismo, el estimador indicó que para cada una de las estaciones (con precipitación y sin ella) se puede registrar una especie más. Sin embargo, si se pretende encontrar a la especie faltante durante la época sin precipitación, el estimador indicó que es necesario aumentar el esfuerzo de muestreo (101 días de colecta) para alcanzar un inventario del 97%. Mientras que para la época con precipitación es necesario aumentar 230 días de colecta, para alcanzar el mismo porcentaje de fiabilidad en el inventario.
Diversidad beta (β). La diversidad beta, con base en el índice de Sorensen, demostró mayor similitud entre las localidades de Veinte Casas y Nuevo San Juan Chamula con un 67%, y una β de 2.15 con base a la riqueza efectiva (Q0). Los valores encontrados en Q1 indicaron que, entre estas localidades, las especies típicas se comparten en un 20% (índice de Morisita) con una β de 1.52. Finalmente, el valor de Q2 (índice de Morisita-Horn) indicó que se comparten un 43% de especies dominantes con una β de 2.01 (Cuadro 3).
Cuadro 3 Diversidad beta con base en los índices de Sorensen, Morisita y Morisita-Horn en las cuatro localidades de la REBISO; ER=Emilio Rabasa, VC=Veinte Casas, NSJC=Nuevo San Juan Chamula y SJ=San Joaquín.
| Localidades comparadas | Sorensen β=2.15 | Morisita β=1.52 | Morisita-Horn β=2.01 | |
|---|---|---|---|---|
| ER | VC | 0.6 | 0.3 | 0.48 |
| ER | NSJC | 0.5 | 0.17 | 0.38 |
| ER | SJ | 0.28 | 0.22 | 0.55 |
| VC | NSJC | 0.67 | 0.2 | 0.43 |
| VC | SJ | 0.4 | 0.84 | 0.85 |
| NSJC | SJ | 0 | 0 | 0 |
Diversidad beta (β) verdadera. De acuerdo con las estimaciones, existen dos comunidades efectivas, lo cual concuerda con el dendrograma elaborado con el índice de similitud de Jaccard, debido a que se refleja que Veinte Casas, Nuevo San Juan Chamula y Emilio Rabasa conforman la primera comunidad efectiva, en tanto que San Joaquín El Progreso representó a la segunda comunidad.
Distribución estacional. La riqueza y abundancia de mántidos fue mayor cuando no se registró precipitación y cuando ésta descendió considerablemente, como en abril y julio, constituyeron los meses más representativos, con cinco y cuatro especies en el caso de la riqueza y para la abundancia 19 y 11 individuos, respectivamente.
Abundancia por especies. Para el caso de la estacionalidad por especies, se encontraron diferencias en cuanto a la distribución de las especies por cada mes de muestreo. Es así como S. m. montana abundó mayormente en el mes de mayo (cinco individuos) cuando la precipitación empezó a aumentar. Aunado a ello, en los meses lluviosos de junio, julio y agosto P. infuscata y Stagmomantis vicina presentaron la tasa de abundancia más alta, julio fue el mes donde se recolectaron seis y tres individuos, respectivamente. En contraste a la época lluviosa, en ausencia de precipitación (abril) se recuperaron siete individuos tanto de S. nahua como de M. major. Por último, de las especies Mantoida maya y Choeradodis rhombicollis solamente se capturaron un ejemplar en los meses de julio y agosto, cuando la precipitación fue muy alta (Fig. 4).

DISCUSIÓN
Este trabajo representó el primer aporte sobre la diversidad y distribución de los mántidos para la Reserva de la Biósfera Selva El Ocote (REBISO). Con ello, se enriquece con cuatro registros nuevos la diversidad conocida para el estado de Chiapas (Hernández-Baltazar & Gómez, 2017; Hernández-Baltazar et al., 2018; Jantsch, 1999; Rehn, 1935a, 1935b; Rodrigues et al., 2017; Roy, 2012; Svenson, 2014; Terra, 1995). Las 17 especies de Chiapas representa el 23.6% de las 72 especies reportadas para México (Hernández-Baltazar et al., 2018), pero solo el 3.41% de las 498 especies registradas para el neotrópico (Rivera, 2010).
Dentro de la fauna registrada, la familia con mayor riqueza específica y abundancia fue Mantidae (cinco especies y 29 individuos), lo cual no es raro, pues es la familia con mayor diversidad en México con 45 especies (Hernández-Baltazar & Gómez, 2017) como en Colombia (Ariza et al., 2012; Arteaga et al., 2014), Brasil (Menezes & Bravo, 2014) y en general en el neotrópico (Ehrmann & Koçak, 2009; Rivera, 2010). A nivel mundial la familia Mantidae reúne el mayor número de especies, incluyendo casi el 50% de mántidos descritos actualmente con 1,016 especies (Wieland & Svenson, 2018).
Stagmomantis fue el género más abundante con tres especies, de las 17 registradas en México (Hernández-Baltazar & Gómez, 2017). El género Stagmomantis ha sido reconocido como el segundo más diverso en el neotrópico y con una distribución exclusiva en América, que se extiende desde Nueva York, EUA hasta la Patagonia (Anderson, 2018; Ehrmann & Koçak, 2009). En otros estudios similares se han encontrado resultados equiparables, como lo encontrado por Ariza et al. (2012), quienes mencionaron a Stagmomantis y Pseudovates (= Phyllovates) como aquellos con mayor presencia en Colombia. Stagmomantis es de tamaño grande, sus preferencias ecológicas son árboles y arbustos en los que tienen el hábito de emboscar (Svenson & Whiting, 2004). La captura de especies de este género se realizó principalmente con trampas de luz, debido a que los machos adultos se sienten más atraídos por las luces en la noche y, en general, son mucho más activos (Anderson, 2018), contrariamente a otros géneros recolectados en este estudio.
En cuanto a la especie S. m. montana, ésta fue la más abundante en la REBISO; probablemente debido a que se trata de una especie propia de selvas húmedas, que se confunde perfectamente en el follaje de este tipo de vegetación desde donde prefiere acechar a sus presas y emboscarlas. La Selva de El Ocote, se encuentra dentro de la distribución de dicha especie, la cual se extiende desde Sinaloa hasta el sur de México, incluido Chiapas, en un amplio rango altitudinal que va de los 200 hasta los 1,100 metros (Rehn, 1935a, 1935b). Por el contrario, especies como C. rhombicollis que se distribuye en vegetación con líquenes donde suele camuflarse (Agudelo & Chica, 2002) y M. maya de tamaño pequeño (15-18 mm), de movimientos muy rápidos y hábitos nocturnos, son difíciles de capturar (Anderson, 2018; Deyrup, 1986; Salazar, 2005). La dificultad de capturarlas hace que estas dos últimas especies sean consideradas como poco frecuentes, al registrarse un solo ejemplar de cada una de ellas. La rareza de estas especies puede suscribirse a lo indicado por Longino & Colwell (1997) quienes definen a las especies raras como aquellas que viviendo y reproduciéndose en el lugar de muestreo, están representadas por poblaciones con pocos individuos. Rabinowitz et al. (1986) reconocen que la rareza de las especies ocurre a diferentes niveles y de distintos tipos, describiendo como rareza demográfica lo encontrado en C. rhombicollis y M. maya debido a la restricción en su distribución geográfica, hábitat y tamaño de las poblaciones.
Las localidades dentro de zonas de conservación (Emilio Rabasa y Veinte Casas) presentaron la mayor diversidad de especies en la Reserva (siete). Lo anterior es posible atribuirlo a que son localidades con áreas de cobertura arbórea y arbustiva suficiente y continua (Arroyo-Rodríguez et al., 2009). Por el contrario, las zonas perturbadas (San Joaquín El Progreso y Nuevo San Juan Chamula) fueron las de menor diversidad de especies (tres), posiblemente por el impacto que tiene la transformación de la selva en vegetación secundaria y zonas con actividades agropecuarias (ganadería, cafetales, huertos); con lo que se promueve la pérdida de especies y la modificación de la funcionalidad del ecosistema (Amat-García et al., 2007). Por tanto, se reconoce que, los mántidos se ven influenciados por la pérdida de su hábitat, ya que a medida que se pierde la cobertura en la selva, no encuentran corredores biológicos idóneos para la disposición continua de presas ni la vegetación para su camuflaje, comportamiento que es fundamental para depredar (Arteaga et al., 2014).
De acuerdo con el estimador Chao 1, en la Reserva se colectó 88% de la fauna de mántidos que potencialmente ocurren dentro de ella, faltando solamente una especie para que el inventario fuera completo. No obstante, la estimación presupone demasiado esfuerzo en la obtención de la especie faltante, que posiblemente podría ser cualquiera de las siguientes diez ya registradas para el estado: Hondurantemna chespiritoi, Liturgusa maya, Melliera chorotega, Stagmomantis carolina, Stagmomantis montana, Stagmomantis parvidentata, Stagmomantis theophila, Stagmomantis venusta, Vates pectinata y Zoolea orba (Hernández-Baltazar & Gómez, 2017; Hernández-Baltazar et al., 2018; Jantsch, 1999; Rehn, 1935a; Rodrigues et al., 2017; Roy, 2012; Svenson, 2014; Terra, 1995).
Los valores de beta y reemplazo de las especies indicaron que existe mayor similitud en las localidades de Veinte Casas y Nuevo San Juan Chamula, con dos especies compartidas: P. infuscata y S. vicina. La compartición de dichas especies podría deberse a que son altamente adaptables y pueden distribuirse a diferentes niveles altitudinales, encontrándose en un rango que va de los 450 a los 2,600 metros (Ariza et al., 2012; Hurd, 1999; Svenson & Whiting, 2004).
Por otro lado, Emilio Rabasa junto con Veinte Casas y Nuevo San Juan Chamula conforman una comunidad efectiva, quedando con ello, San Joaquín El Progreso como otra. Esto se debe a la cercanía entre las localidades de muestreo, lo que da lugar a hábitats con condiciones similares, mismo tipo de vegetación y actividades agropecuarias (Wright & Reeves, 1992). Tanto Emilio Rabasa como Veinte Casas poseen selvas conservadas y Nuevo San Juan Chamula presenta áreas de remanentes de selva conservada, lo que condiciona que las especies compartidas entre estas localidades tengan las condiciones apropiadas para su estabilidad en la Reserva. Por el contrario, San Joaquín El Progreso es una localidad con un grado de perturbación elevado, en ella predomina los cafetales y áreas abiertas que generan sitios que limitan la distribución de las especies, repercutiendo en el declive de los mántidos (Amat-García et al., 2007). Además, las variaciones en la composición florística y cambios graduales en las condiciones ambientales por cada cambio altitudinal conllevan a la gran disimilitud que presenta con las localidades más conservadas (Sonco-Suri, 2013). Esta segunda comunidad efectiva, San Joaquín El Progreso, presenta una serie de adversidades que no han impactado del todo a la mantidofauna del sitio, pues aún presenta parches que siguen ofreciendo algunos recursos que son aprovechados por los mántidos para su supervivencia y sus poblaciones aún se mantienen estables y con ello, pueden colonizar los nuevos hábitats producto de las intervenciones antrópicas que presenta la localidad (Arteaga et al., 2014; Calderón-Patrón et al., 2012; Gurrutxaga, 2004; Rabinovich, 1978).
Ponderando la riqueza y abundancias por mes, abril y julio tuvieron los rangos más altos. No obstante, en un análisis por temporadas (con lluvia o sin ella), estos valores son equilibrados, con cinco especies y 19 individuos, respectivamente. Ambos resultados se ajustan al ciclo de vida de los mántidos, que puede durar de diez meses a un año en condiciones naturales (Agudelo & Chica, 2002; Mariño-Pedraza, 2011; Prete et al., 1999). Mariño-Pedraza (2011) mencionó que la presencia de ninfas y adultos de estos insectos ocurre en primavera y verano, lo que coincide con la mayor concentración de estos insectos en los meses correspondientes.
Existe poca literatura que corrobore los meses de actividad de cada especie. Maxwell (2014) indicó que las especies de Stagmomantis emergen entre abril y junio, las ootecas aparecen de septiembre a octubre, por lo que no es raro que en las localidades de la Reserva se hayan colectado las especies de este género (S. m. montana, S. nahua y S. vicina) en los meses de abril, mayo, julio y agosto.
Lücking et al. (2010) mencionaron que un valor alto de precipitación es de gran importancia para la presencia de C. rhombicollis. Probablemente por esto en el mes de julio, con una precipitación considerable (136.7 mm), se haya recolectado esta especie. Para el caso de M. maya, recolectado en agosto, Anderson (2018) documentó su estacionalidad y menciona que esta especie ocurre con mayor frecuencia entre principios de junio hasta mediados de septiembre. Los adultos de P. infuscata se encuentran en los meses de febrero a septiembre (Rehn, 1935a) coincidente con los meses que se colectaron. Por último, de M. major se indica que los adultos están activos de julio a diciembre (Hebard, 1922), por lo que, no es extraño encontrar a esta especie en abril en la Reserva Selva El Ocote.

