











 text new page (beta)
text new page (beta) Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO 
 Permalink
PermalinkIntroducción
Las mariposas agrupadas actualmente en la superfamilia Papilionoidea (Van Nieukerken, 2011; Heikkilä et al., 2012), comprenden en México especies en seis familias que corresponden, según el orden de mayor a menor diversidad a Hesperidae, Lycaenidae, Nymphalidae, Riodinidae, Pieridae y Papilionidae (Luis et al., 2000, 2004). Este grupo también se conoce tradicionalmente como Rhopalocera y se estima que para México hay 2,105 taxones (Luis et al., 2004, Llorente et al., 2014). Con base en las síntesis de Shields (1989) y Heppner 1991, 1998, México tiene 9.4% de las especies descritas de Papilionoidea de todo el mundo.
La situación geográfica de México, su complejidad topográfica y su historia geológica han propiciado que exista una variedad amplia de ecosistemas que a la vez resguardan una notable riqueza de especies (Morrone et al., 1999), no menor en este grupo del orden Lepidoptera (Llorente et al., 2014). Adicionalmente, el avanzado conocimiento sobre la taxonomía de las mariposas diurnas, su ubicuidad, su abundancia y la facilidad de recolección e identificación, han contribuido a que sean consideradas como un grupo indicador del estado de los hábitats y su riqueza (Llorente et al., 1993), por lo que es importante continuar desarrollando el conocimiento de sus poblaciones y especies.
El estado de Oaxaca sobresale en el país como el de mayor endemismo en Papilionoidea (Llorente et al., 2014). De los ecosistemas mexicanos, el bosque mesófilo de montaña es uno de los biomas más importantes en México, debido a la alta proporción de taxones endémicos a nivel específico o subespecífico que en él se encuentran (Halffter, 1976, 1987; Escalante-Pliego et al., 1998). Por otro lado, en un análisis de la distribución de especies en unidades biogeográficas dentro de las Áreas Naturales Protegidas de México, se ha identificado a los bosques mesófilos del este de México, particularmente en el norte de Oaxaca, como áreas inadecuadamente protegidas (Challenger, 1998; Navarro et al., 2004), por lo que es importante ampliar su conocimiento.
Con base en la información sobre la distribución geográfica de cada una de las especies y el reconocimiento de sus áreas originales, se ha venido detectando la posible extinción local de poblaciones, producto de la disminución gradual y constante de sus hábitats por efecto del crecimiento de los asentamientos humanos y el cambio de uso de suelo para ganadería y agricultura (Luis et al., 2004). Estos factores motivan a que se generen trabajos e investigaciones sobre las mariposas diurnas en las regiones que no hayan sido adecuadamente estudiadas, para que sirvan de línea de base en evaluaciones posteriores.
El estado de Oaxaca ha sido atractivo para los exploradores naturalistas desde finales del siglo XIX y principios del XX (Godman & Salvin, 1878-1901; Seitz, 1907-1924, entre otros). Hoffmann (1976) añadió 180 especies para Oaxaca a las 160 previamente conocidas, para obtener un total de 340 especies para este estado (Luis-Martínez et al., 1991). Otro referente importante es el trabajo Luis et al. (1991) en el que realizan un estudio sistemático sobre Papilionoidea en la Sierra de Juárez en Oaxaca, integrando 452 especies. Asímismo, Luis et al. (2004) reunieron 25,000 registros para todo el estado de Oaxaca; sumando 491 localidades de muestreo con 1,103 especies y 94 subespecies con un total de 1,197 taxones. De estos, sólo 302 registros (en 112 taxones) pertenecen a 9 localidades de la Sierra Mazateca (Fig. 1). Y más recientemente se publicó un trabajo sobre los avances faunísticos sobre los Papilionoidea del estado de Oaxaca; en este se presenta una lista de 1331 taxones a nivel específico, repartidos en 462 generos, 24 subfamilias y y seis familias de Papilionoidea (sensu lato) del estado de Oaxaca. Esta lista tiene un énfasis en 3 areas geográficas del estado, la región de A) valle de Cuicatlán, que colinda con la Sierra Mazateca hacia el este, B) región Mixe, y C) región Loxicha (Luis-Martinez et al., 2016).
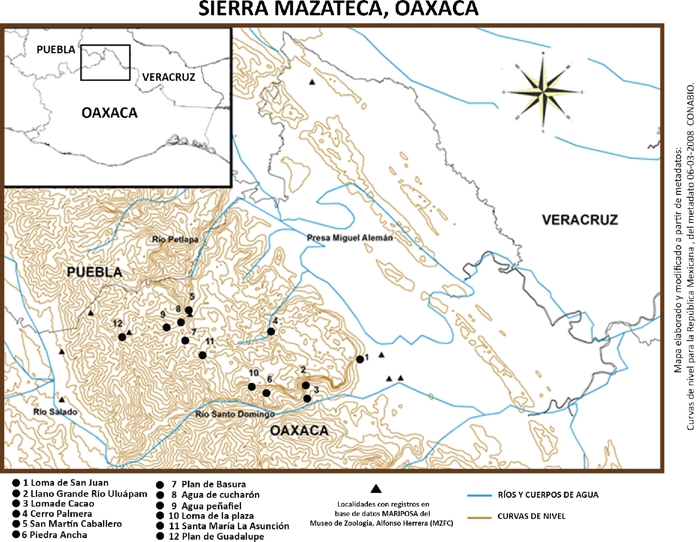
Figura 1 Mapa de la Sierra Mazateca con los sitios de recolecta de este estudio. Adicionalmente se integran sitios con recolectas esporádicas, (registros en Base MARIPOSA). Metadatos tomados del portal de geoinformación de CONABIO: curvas de nivel, hidrología y división política.
En un gradiente altitudinal, la temperatura disminuye al aumentar la altitud como consecuencia directa de la escasa densidad del aire y de su relativa carencia en vapor de agua y anhídrido carbónico, lo que disminuye su capacidad de absorber calor (Miller, 1982). La temperatura influye directamente en la actividad de las mariposas, debido a que son organismos ectotérmicos que dependen totalmente del ambiente para regular su temperatura corporal; y en ambientes con bajas temperaturas se afecta el vuelo de los imagos limitando su actividad para eventos reproductivos y/o de alimentación. Adicionalmente, hay especies que sólo pueden encontrarse en determinada altitud, hecho relacionado directamente con la distribución de sus plantas de alimentación. En México existen pocos estudios que describan la distribución altitudinal de la fauna de mariposas en una región determinada, debido a que 90% de los inventarios faunísticos se han realizado en áreas por debajo de los 1,500 m de altitud o en una sola estación de recolecta (Vargas et al., 1999).
Con esta perspectiva nos planteamos los siguientes objetivos para la Sierra Mazateca ubicada en el estado de Oaxaca:
Materiales y métodos
Descripción del área de estudio. La Sierra Mazateca se encuentra al norte del estado de Oaxaca y cubre aproximadamente 1,050 km2 del sistema montañoso, se localiza entre los 18°00 y 18°22 N y 96°30 y 97°15 O. Se ubica entre el Río Salado en el Valle de Tehuacán Cuicatlán y la planicie Costera del Golfo en los límites de Oaxaca y Veracruz; se limita al norte por el río Petlapa, entre Puebla y Oaxaca; y al sur por el río Santo Domingo (Fig. 1).
Políticamente la Sierra Mazateca se encuentra dentro del distrito de Teotitlán el cual está integrado por 25 municipios. Los municipios a los que existe un mejor acceso son: San Bartolomé Ayautla, San Juan Coatzospam, Huautepec, Santa María La Asunción, Huautla de Jiménez, San Mateo Yoloxochitlán, Santa Cruz Acatepec, San Jerónimo Tecoatl; ya que se encuentran próximos a la carretera federal 182 que va de Tuxtepec a Teotitlán del Camino. Los demás están conectados por caminos de terracerías.
La Sierra Mazateca pertenece a una sola provincia, la provincia de Sierras Orientales y a la subprovincia de la Sierra Madre de Oaxaca. La topografía es muy accidentada y presenta un gradiente altitudinal que va de los 100 a 2,750 msnm En la zona se encuentran laderas con pendientes pronunciadas y cañadas profundas, y en algunas áreas pequeños valles (Munn, 2003, citado en Torres Colín, 2004). Esta Sierra posee cuatro tipos de suelos de acuerdo con su compleja topografía, estos son: regosoles, luvisoles, cambisoles y rendzinas. Pertenece a la región hidrológica R-H28 Cuenca del río Papaloapan, en donde se ubican las cuencas del río Santo Domingo y río Petlapa. Según García 1988 y 1998 en la Sierra Mazateca se presentan 11 tipos climáticos: A(f), (A)C(fm), (A)C(m), (A)C(wo), Am, Aw2, Awo, , BS1hw, C(f), C(m) y C(w2).
En la Sierra Mazateca se encuentran cuatro tipos de vegetación. A continuación se presenta un breve resumen de los presentes en la Sierra Mazateca, tomado del trabajo de Torres-Colín (2004). El primero es el bosque caducifolio que se establece desde los 600 a los 2,000 msnm, en sitios donde predominan clima templado y suelos profundos con materia orgánica. Por lo general forma parte de agrupaciones secundarias en los bosques templados. Las especies por la que está dominado son Liquidambar styraciflua L., Pinus patula Schltdl. & Cham., Clethra conzattiana L. M. González, Dendropanax leptopodus (H.B.K.) Dcne et Manchon, Neolitsea serícea (Blume.) Koidz., Saurauia Willd. y Quercus Nixon. El segundo es el bosque mesófilo de montaña que se distribuye en las laderas y cañadas húmedas, a altitudes entre 1000 y 2500 msnm, en donde predominan climas semicálidos y templados húmedos con neblina y frecuentes lloviznas los suelos son profundos, arcillosos y con una gruesa capa de materia orgánica y se derivan principalmente de rocas metamórficas y afloramientos de rocas volcánicas (Rzedowski & Palacios, 1977; Arellanes, 2000) (ver Fig. 2). Fisonómicamente este bosque es denso y lo pueden formar de 2 o 3 estratos arbóreos con una rica composición florística. El estrato principal lo forman árboles de 7 a 20 m de altura de las especies Quercus candicans Née, Pinus patula Schltdl. & Cham., Liquidambar styraciflua L., Weinmannia pinnata L., Styrax glabrescens Benth., Ternstroemia sylvatica Cham. & Schldl., Ilex discolor Hemsl, Dendropanax populifolius (Marchal) A.C. Smith., Ocotea helicterifolia (Meisn.) Hemsl., Persea americana Mill., Podocarpus matudae Lundell, Symplococarpon purpusii (Brandegee) Kobuski, Cyathea Sm., Clethra L. y Saurauia Willd. Entre las variantes de este tipo de bosque se encuentran comunidades achaparradas que se desarrollan en elevaciones cercanas a los 2,500 msnm sobre suelos arcillosos o algo arenosos, ácidos permeables, poco a medianamente profundos con materia orgánica, sobre rocas ígneas intrusivas o metamórficas (Ishiki, 1988). Consiste en un estrato arbóreo de 4 a 6 m de altura donde dominan Weinmannia glabra L. f., Clethra odorata Hook. & Arn., Lyonia squamulosa M. Martens & Galeotti, Pinus oocarpa Schiede y Clethra mexicana Schiede. (Ishiki, 1988) (Fig. 2). También se pueden reconocer en la Sierra Mazateca otras variantes del bosque mesófilo de montaña donde el estrato dominante alcanza entre 30 y 60 m de altura, en lugares muy húmedos, a una altitud entre 1,000 y 2,300 msnm (Rzedowski & Palacios, 1977; Munn, 2003, citado en Torres Colín, 2004) sobre pendientes fuertes con suelo bien desarrollado, donde la especie dominante es Oreomunnea mexicana (Standl.) Leroy asociada con Ulmus mexicana (Liebm.) Planch., Quercus candicans Née, Magnolia schiedeana Schldl., Persea americana Mill., Billia hippocastanum Peyr. y Podocarpus matudae Lundell. El tercer tipo de vegetación que se encuentra en la Sierra Mazateca, es el bosque de pino, particularmente se le registra en el municipio de Huautla de Jiménez. Este se desarrolla en elevaciones de 300 a 3, 000 m, en un clima con frecuencia templado subhúmedo, aunque puede haber casos que se desarrolle en climas semicálidos o cálidos húmedos sobre rocas metamórficas de tipo esquisto, en suelos arcillosos, someros y ricos en materia orgánica. La composición florística puede estar conformada exclusivamente por especies del género Pinus L. o compartir hábitats con otros géneros como Quercus L., Liquidambar L. y Alnus Mill. El cuarto tipo de vegetación encontrado en la Sierra, es la selva alta perennifolia, que se desarrolla en elevaciones entre 200 y 1,000 msnm, con clima cálido húmedo, sobre suelos kársticos o lateríticos originados por aluviones. El estrato superior está compuesto por elementos de 30-40 m como Terminalia amazonica (J. F. Gmel.) Exell, Cordia alliodora (Ruiz & Pav.) Oken, C. megalantha S.F. Blake, Calophyllum brasiliense Cambess., Dialium guianense (Aubl.) Sandwith, Tapira mexicana Schldl., Elaeagia uxpanapensis Lorence, Dussia mexicana (Standl.) Harms y Pera barbellata Standl; en el estrato medio dominan árboles de 15 a 25 m como Spondias radkolferi Donn. Sm., Cymbopetalum penduliflorum (Dunal) Baill., Sloanea tuerckheimii Donn. Sm., Brosimun guianense (Aubl.) Huber, Nectandra membranacea (Sw.) Griseb., Hieronyma oblonga (Tul.) Mull. Arg. y Ficus insipida Willd.; el estrato bajo contiene elementos de 3-10 m con dominancia de Pleuranthodendron lindenii (Turcz.) Sleumer, Ficus obtusifolia Kunth, Guatteria galeottiana Baill, Cymbopetalum baillonii R.E. Fr. y Miconia Ruiz & Pav. (ver Fig. 2).
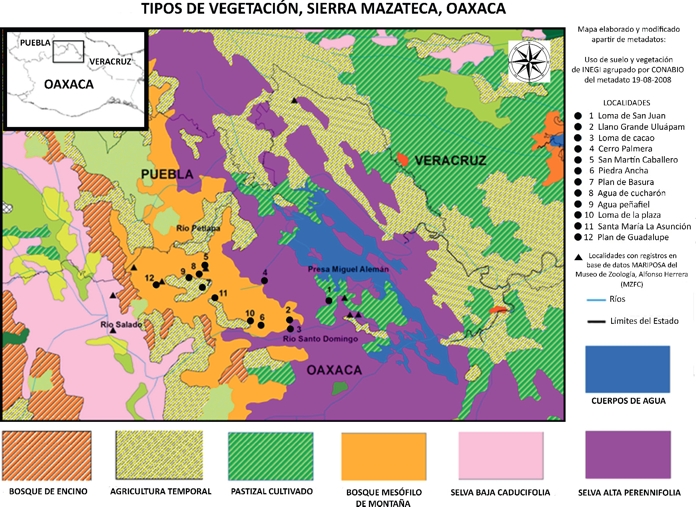
Figura 2 Tipos de vegetación en la Sierra Mazateca, Oaxaca. Mapa construido a partir de Metadatos de acuerdo a CONABIO, 2008 (Uso de suelo y vegetación de INEGI agrupado por CONABIO) y localización de los sitios de colecta.
Método. El área de estudio se encuentra dentro del distrito de Teotitlán. La elección de las localidades se hizo tomando los siguientes criterios: fácil acceso, grado de perturbación bajo, que estén dentro de un gradiente altitudinal entre 100 y 2500, y abarcar los principales tipos de vegetación (bosque mesófilo de montaña, bosque caducifolio, selva alta perennifolia y bosques de pino encino) (Tabla 1, Fig. 2). Se realizaron siete salidas de campo (una por mes) que duraron de cinco a seis días, completando 35 días de recolectas, abarcando los meses de junio a diciembre de 2010 y mayo del 2011, que incluye la temporada de mayor riqueza de mariposas (Vargas et al., 1999).
Tabla 1 Características de localidades de recolecta en la Sierra Mazateca, Oaxaca. SAP = selva alta perennifolia, BMM = bosque mesófilo de montaña.
| Municipio | Localidad | Altitud (m s.n.m.) | Longitud | Latitud | Vegetación | Tipo de Clima1 |
|---|---|---|---|---|---|---|
| San Felipe Jalapa de Díaz | Loma de San Juan | 129 | −96.542814 | 18.096881 | SAP | Am, (A)C(m), A(f) |
| San Felipe Jalapa de Díaz | Llano Grande río Uluapan | 354 | −96.643982 | 18.057217 | SAP | Am, (A)C(m), A(f) |
| San Bartolomé Ayautla | Loma de Cacao | 470 | −96.642132 | 18.034972 | SAP-BMM | Am, (A)C(m), A(f) |
| San José Tenango | Cerro Palmera | 992 | 96.693636 | 18.155669 | BMM-SAP | (A)Cf, (A)C(m), C(m), (A)C(w2) |
| Santa María Chilchotla | San Martín Caballero | 1234 | −96.846798 | 18.187598 | BMM | (A)Cf, (A)C(m), C(m), (A)C(w2) |
| San Bartolomé Ayautla | San Martín Piedra Ancha | 1369 | −96.712529 | 18.044037 | BMM | (A)Cf, (A)C(m), C(m), (A)C(w2) |
| Huautla de Jiménez | Plan de Basura | 1398 | −96.852698 | 18.135004 | BMM-BP | (A)Cf, (A)C(m), C(m), (A)C(w2) |
| Eloxochitlán de Flores Magón | Agua de Cucharón | 1442 | −96.860482 | 18.166906 | BMM | (A)Cf, (A)C(m), C(m), (A)C(w2) |
| Santa Cruz Acatepec | Agua Peñafiel | 1583 | −96.885189 | 18.157813 | BMM | (A)Cf, (A)C(m), C(m), (A)C(w2) |
| San Juan Coatzospam | Loma de la Plaza | 1718 | −96.737713 | 18.054881 | BMM | (A)Cf, (A)C(m), C(m), (A)C(w2) |
| Santa María La Asunción | Santa María La Asunción | 1740 | −96.822921 | 18.109645 | BMM | (A)Cf, (A)C(m), C(m), (A)C(w2) |
| Santa María Teopoxco | Plan de Guadalupe | 2199 | −96.962702 | 18.140938 | BMM-BP | (A)C(m)(f), C(m), C(f) |
Debido a las condiciones metereológicas, o que algunas localidades tenían un grado de perturbación más alto, el esfuerzo de recolecta no fue igualmente productivo en ellas, fue el caso de cuatro de las 12 las localidades visitadas (Loma de San Juan, Cerro Palmera, Santa María La Asunción y Plan de Guadalupe).
Las recolectas se efectuaban de las 9:00 a las 18:00 hrs. Se emplearon dos técnicas: a) búsqueda dirigida con red entomológica aérea y b) uso de trampas Van Someren-Rydon (Rydon, 1964). La primera consistió en buscar los imagos en los distintos microhábitats, caminos o senderos, zonas de penumbra, sitios húmedos, áreas con inflorescencias y algún otro sitio de afluencia. La segunda técnica consistió en colocar de cinco a siete trampas cebadas a lo largo de un transecto de 1 km, alternadas lateralmente, tratando de incluir diferentes microhábitats a una altura de 1.5 a 5m, tratando de que no incidieran directamente los rayos solares. El cebo consistió de plátano macho fermentado con azúcar mascabado y cerveza. Después de obtenidos los ejemplares se sacrificaron con un ligero apretón en el tórax y fueron colocados en bolsas individuales con los datos geográficos de la localidad de recolecta, nombre del colector, y altitud.
La determinación taxonómica se efectuó en la Colección Nacional de Insectos: Lepidoptera, del Departamento de Zoología del Instituto de Biología de la UNAM (IB-UNAM), misma donde se depositó el material recolectado. Para ello se preparó una muestra representativa de las especies recolectadas con base en las técnicas sugeridas por Morón & Terrón (1988). Los ejemplares se compararon con ejemplares de la colección, así como mediante la consulta de obras con claves ilustradas de Godman & Salvin (1878-1901), Seitz (1907-1924), Howe (1975), Beutelspacher (1980), Scott (1986), De la Maza (1987), De Vries 1987, 1997, Llorente-Bousquets et al. (1997), Luis et al. (2003) y Vargas et al. (2008); y, para el género Calephelis se consultó el trabajo de McAlpine (1971); Llorente (1984), para el género Enantia Hübner; y Duarte & Robbins (2010) para algunos licénidos. En el caso de Hesperidae se consultaron los trabajos de Evans 1951, 1952, 1953 y 1955. También se realizó la técnica de microdisección estandarizada por Burns (1997) para obtener genitalias y así comprobar la determinación en algunas especies.
La lista de especies se elaboró tomando como base los ejemplares recolectados junto con los registros previos incluidos en la Megabase Mariposa del Museo "Alfonso Herrera" de la Facultad de Ciencias de la UNAM, y una colección particular (Colección Galindo). Para el arreglo taxonómico de la superfamilia Papilionoidea se siguió el orden de las familias propuesto por Van Nieukerken et al. (2011), y dentro de las familias la nomenclatura adoptada por Llorente-Busquets et al. (2006); y para la familia Hesperidae, los estudios morfológicos y moleculares realizados por Warren (2006) y Warren et al. 2008, 2009.
En la estimación de la riqueza específica del área estudiada, se empleó la construcción de una curva de acumulación de especies, conforme al modelo descrito por Soberón & Llorente (1993), empleando los programas EstimateS 8.0 (Colwell, 2006) y Statistica (StatSoft, 1998). El proceso consistió en aleatorizar los datos con el programa para evitar que al añadir las muestras, el orden afectara la forma de la curva (Moreno, 2001; Jiménez-Valverde & Hortal, 2003).
Resultados
Composición faunística. Con base en la determinación de 3,108 ejemplares recolectados en el transcurso de este trabajo, la revisión de la Colección Galindo (103 ejemplares) y la revisión de la megabase MARIPOSA de la Colección de la Facultad de Ciencias, UNAM (298 registros provenientes del área de estudio) se integró la lista de la superfamilia Papilionoidea que se distribuye en la Sierra Mazateca, Oaxaca, la cual consta de 460 especies-subespecies, incluidos en seis familias, 19 subfamilias y 246 géneros. La familia con mayor número de taxones en nuestra región fue Nymphalidae (168), seguidas en orden descendiente por Hesperiidae (141), Lycaenidae (57), Pieridae (42), Riodinidae (33) y Papilionidae (19), en proporciones semejantes a la presencia de estas familas en el estado de Oaxaca (Figura 3). Se excluyeron cuatro taxones, dado que se duda sobre su determinación, estas son: Pereute charops nigricans Joicey & Talbot, 1928, Catonephele cortesi R. G. Maza, 1982, Hesperocharis graphites H. Bates, 1864 y Parides panares lycimenes (Boisduval, 1870). De las 460 taxones, sólo 113 (50 especies-subespecies no fueron recolectadas en este estudio) están registrados en la megabase Mariposa con base en recolectas esporádicas, de modo que 347 taxones son nuevos registros para la Sierra Mazateca.
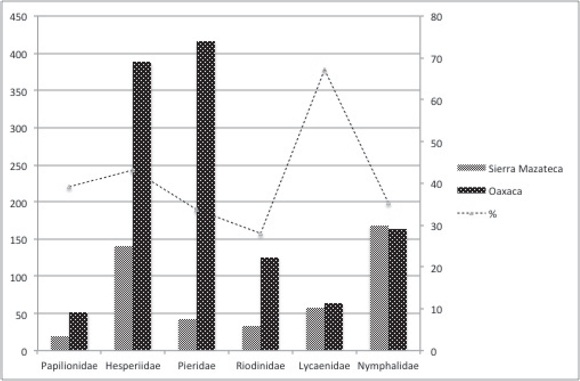
Figura 3 Comparación de los taxones (número y porcentaje) de Papilionoidea de la Sierra Mazateca con respecto al total en el estado de Oaxaca (100%). Los datos del total para Oaxaca fueron tomados de Luis et al, 2004 y 2011.
Llorente-Bousquets et al. (2006) y Luis-Martínez (2016) citan ocho y 10 taxones del género Calephelis para el estado de Oaxaca. En este estudio se recolectaron siete especies del genero Calephelis, cinco de estos taxones ya reportados por estos autores y de dos no se tenía registro, por lo tanto se determina que son nuevos registros para el estado de Oaxaca, siendo estas: C. aztecaMcAlpine, 1971 y C. tikal Austin, 1993. De acuerdo con los trabajos de Luis et al. 2004, 2016 se determinan también como nuevos registros de la familia Hesperiidae para el estado de Oaxaca: Cymaenes laurelolus laureolus (Schaus, 1913) y Niconiades sp. nov. Bolla aff. cylindus (Godman & Salvin, 1896) posiblemente sea también nuevo registro para México y Oaxaca, sin embargo, sólo se recolectó 1 ejemplar y esperamos a que sea confirmada su presencia, con la recolecta de más ejemplares.
México posee 423 especies y subespecies endémicas de Papilionoidea según la cifra dada por Luis et al. (2011). Más tarde se volvieron a contabilizar consideranto hasta el nivel subespecífico, habiendo para Hesperiidae 155 sspp., Papilionidae 32 sspp., Pieridae 37 sspp., Lycaenidae 10 sspp., Riodinidae 44 sspp. y Nymphalidae 159 sspp., sumando un total de 437 sspp. (Llorente-Bosquets et al., 2014). De las cuales 26 están presentes en la Sierra Mazateca (Tabla 2), y dos son exclusivos del estado de Oaxaca: Catasticta teutila flaviaciata y Morpho theseus utae El 99.34% (457) de los taxones que se encuentran en la Sierra Mazateca presentan una distribución dentro de la región Neotropical (Lamas 2004), pero sólo 288 (62.6%9 son exclusivos a esta región. Y el por el contario, 37.39% (172) de los taxones que aparecen en listas o están presentes en la región Neartica, pero sólo nueve son exclusivos de esta región (Pelham, 2012, desglose por familias en Tabla 3). Para Papilionidae en la región neotropical, en la Sierra Mazateca suman 19 especiessubespecies, para Hesperiidae 140, Pieridae 42, Riodinidae 32, y Nymphalidae 167 y Hesperiidae 140 especies-subespecies. En la región, Neartica, el número de especies por familia es el siguiente: Papilionidae 10 especies subespecies, Hesperiidae 60, Pieridae 25, Lycaenidae 20, Riodinidae 2, y Nymphalidae 55. En la tabla 3 se puede observar el número de taxones que son afines a cada región dentro de los parentesis.
Tabla 2 Lista completa de especies y subespecies presentes en la Sierra Mazateca de Oaxaca, indicando su distribución altitudinal. **, Taxones no colectados en este trabajo reportadas en las colecciones de la UNAM; ◊, taxones determinados en colección particular no reportados en la literatura, bases o colecciones; ▲, registros previos para la zona; ○, Nuevos registros para Oaxaca; E, endémicos para México; E, endémicos para Oaxaca.

Tabla 3 Endemismos, distribución y afinidades regionales de Papilionoidea de la Sierra Mazateca, Oaxaca. (Números en paréntesis indican los taxones afines a cada región).
| Endémica de México | Endémica de Oaxaca | Neártica Pelham (2012) | Neotropical | |
|---|---|---|---|---|
| Lamas (2004) | ||||
| Papilionidae | 3 | 0 | 10 (1) | 19 (10) |
| Hesperiidae | 6 | 0 | 60 (5) | 139 (82) |
| Pieridae | 4 | 1 | 25 (0) | 42 (17) |
| Riodinidae | 6 | 0 | 2 (0) | 32 (32) |
| Lycaenidae | 0 | 0 | 20 (0) | 57 (37) |
| Nymphalidae | 7 | 1 | 55 (2) | 168 (113) |
| TOTAL | 26 | 2 | 172 (8) | 457 (291) |
Reportamos casos en los que en el gradiente altitudinal se encontraron dos subespecies de la misma especie. En el caso de Parides panares lycimenes (Boisduval, 1870) se trata de dos ejemplares (número de catálogo: 365108 y 365109) en la base de datos MARIPOSA, del Museo Alfonso Herrera, de la Facultad de Ciencias, provenientes de la localidad de Jalapa de Díaz, Oaxaca. Esta subespecie solo se distribuye desde Chiapas a Panamá y pensamos que posiblemente está erróneamente determinado, por lo que no la añadimos a la lista, solamente conservamos la subespecie nominal. Mantenemos dos casos en los que sí pensamos que las dos formas están presentes en el gradiente, aunque no en los mismos pisos altitudinales o hábitats. El primer par son: Enantia mazai diazi Llorente, 1884 y Enantia mazai mazaiLlorente, 1984, las cuales con base a un estudio aún no publicado usando caracteres moleculares parece que se trata de la misma subespecie (C. Pozo, com. pers.). El segundo par es: Catasticta teutila teutila (Doubleday, 1847) y Catasticta teutila flavifasciata Beutelspacher, 1986, donde en la primera los 13 ejemplares presentan los puntos y líneas en su vista ventral de color anaranjado, tal como se ha señalado en su descripción, mientras que en la segunda, el ejemplar presentó la coloración amarilla de los puntos y líneas en su vista ventral, como se menciona en la descripción como principal carácter para separarla de la subespecie típica.
Estimación de la riqueza. Para calcular la completitud del muestreo y la riqueza potencial de taxones se consideró la ecuación de Clench. Este modelo está considerado para estudios en sitios de área extensa: bajo el supuesto de que cuanto más tiempo se pase en el campo mayor es la probabilidad de añadir nuevas especies al inventario (Soberón & Llorente, 1993, Moreno, 2001).
De acuerdo con la ecuación de Clench (R2 = 0.998481434) se estimó un total potencial de 533 especiessubespecies pertenecientes a Papilionoidea para el área de estudio (Fig. 4). Entonces en este estudio se recolectaron el 72% (380 taxones) de los estimados. Se le deben agregar 50 más registrados previamente en el área de estudio que no se recolectaron y que posiblemente persistan en el área, datos provenientes de las Colecciones de la UNAM y además sumar 35 taxones determinados por el primer autor en la Colección Galindo, dando un total de 465 taxones y 87.2% del total estimado. Por lo tanto, aún se esperaría encontrar en la Sierra Mazateca 68 más.
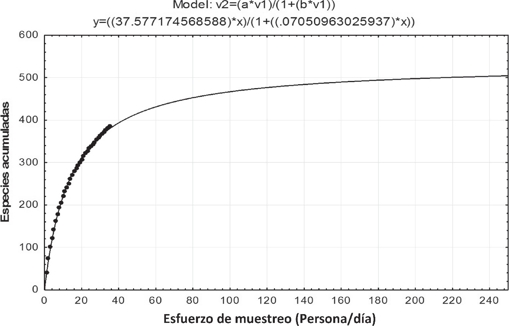
Figura 4 Curva de acumulación de especies de Papilionoidea y Hesperioidea de la Sierra Mazateca, Oaxaca, empleando la ecuación de Clench.
Distribución altitudinal. El gradiente altitudinal que comprende el área estudiada fue de 90-2200 msnm En este gradiente se observó que la riqueza disminuye conforme aumenta la altitud; el 77% (360 taxones) se distribuyen en el intervalo de 90 a 500 msnm y de ellos 223 (48%) se restringen a este piso altitudinal. En el intervalo altitudinal de los 900-1500, se encuentran 200 (44%) y 57 están restringidos a él. En el intervalo de los 1500-2000 se encuentran 102 (22%) y 13 están restringidos a él. Y finalmente a una altitud superior, en el intervalo de los 2000-2200 se encuentran solo 37 (8%) y ocho están restringidos (Tabla 4).
Tabla 4 Riqueza de taxones encontrados en cada intervalo altitudinal (msnm). P = número de taxones presentes y R = taxones restringidos en cada intervalo altitudinal.
| 90-500 | 900-1500 | 1500-2000 | 2000-2200 | |||||
|---|---|---|---|---|---|---|---|---|
| Familia | P | R | P | R | P | R | P | R |
| Papilionidae | 17 | 13 | 7 | 0 | 5 | 0 | 3 | 0 |
| Hesperiidae | 105 | 68 | 58 | 26 | 30 | 4 | 7 | 4 |
| Pieridae | 30 | 10 | 28 | 4 | 17 | 2 | 10 | 0 |
| Riodinidae | 32 | 22 | 12 | 2 | 4 | 0 | 1 | 0 |
| Lycaenidae | 44 | 33 | 19 | 8 | 7 | 2 | 6 | 2 |
| Nymphalidae | 132 | 77 | 76 | 17 | 39 | 5 | 10 | 2 |
| Total | 360 | 223 | 200 | 57 | 102 | 13 | 37 | 8 |
La disminución de la riqueza conforme aumenta la altitud también fue encontrada para otras áreas del país como la Sierra de Juárez, Oaxaca (Luis et al., 1991), la Sierra de Atoyac, Guerrero (Vargas et al., 1992) y la Sierra de Manantlán en Jalisco y Colima (Vargas et al., 1999), donde se consideraron intervalos altitudinales similares.
De los 460 taxones registrados para la Sierra Mazateca, solo tres (Pyrisitia nise nelphe R. Felder, Hermeuptychia hermes Fabricius y Pyrgus oileus Linnaeus) se distribuyen en las 12 localidades recolectadas en este trabajo; tres más (Eurema mexicana mexicana Boisduval), Diaethria anna anna Guérin-Ménéville y Smyrna blomfildia datis Fruhstorfer) en 10 localidades; cuatro (Eurema salome jamapa Reakirt, Hypanartia lethe Fabricius, Anthanassa ardys ardys Hewitson, y Pyrgus oileus Linnaeus) en nueve localidades; seis (Eurema salome jamapa Reakirt, Nathalis iole Boisduval, Morpho helenor montezuma Guenée, Hypanartia lethe Fabricius, Eresia phillyra phillyra_Hewitson y Urbanus dorantes dorantes Stoll) en ocho localidades. Ocho taxones en siete localidades, 10 en seis localidades, 18 en cinco localidades, 29 en cuatro localidades, 60 se distribuyen en tres localidades, 83 en dos localidades y 242 resultaron exclusivas a una sola localidad.
Las localidades con la mayor riqueza fueron Loma de Cacao (470 msnm) con 236 taxones, de éstas, 74 fueron exclusivos de esta localidad; y Rio Uluapam (354 msnm) con 195 especies-subespecies, de ellas 78 son exclusivos. Ambas localidades reúnen el 33% (152 taxones) del total de taxones exclusivos en la Sierra Mazateca (0 a los 500 msnm) y concurren el 70% (325) del total de los registros no exclusivos (Fig. 5).
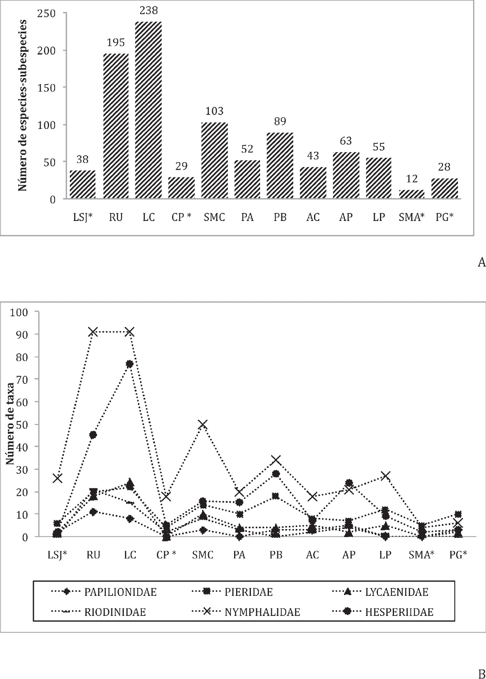
Figura 5 A. Riqueza de mariposas por localidad en la Sierra Mazateca, Oaxaca organizadas de acuerdo al gradiente altitudinal: LSJ = Loma San Juan, RU = Loma de Cacao, CP = Cerro Palmera, SMC = San Martín Caballero, PA = Piedra Ancha, PB = Plan de Basura, AC = Agua de Cucharon, AP = Agua peñafiel, LP = Loma de la Plaza, SMA = Santa María La Asunción y PG = Plan de Guadalupe. (*) Localidades con menor esfuerzo de recolecta. B. Riqueza de los taxones por familia en las localidades muestreadas en la Sierra Mazateca, Oaxaca. LSJ = Loma San Juan (129 msnm), RU = Loma de Cacao (470 msnm), CP = Cerro Palmera (992 msnm), SMC = San Martín Caballero (1234 msnm), PA = Piedra Ancha (1369 msnm), PB = Plan de Basura (1398 msnm), AC = Agua de Cucharón (1442 msnm), AP = Agua Peñafiel (1583 msnm), LP = Loma de la Plaza (1718 msnm), SMA = Santa María La Asunción (1740 msnm) y PG = Plan de Guadalupe (2199 msnm).* = menor esfuerzo de recolecta.
Discusión
Con respecto a otras Sierras del país, como la Sierra de Juárez, Oaxaca (Luis et al., 1991, 2016) (Tabla 5), la Sierra Mazateca tuvo menor riqueza de especies, sin embargo, cabe mencionar que en este estudio no se muestreó durante todo el año, por lo que podría esperarse que aumentara el número de especies en el área, si se considera que hay especies estacionalmente asociadas. En comparación con la Sierra de Atoyac, Guerrero (Vargas et al., 1992) y la Sierra de Manantlán en Jalisco y Colima, (Vargas et al., 1999), la Sierra Mazateca presentó un número similar de especies a pesar del menor esfuerzo de recolecta. También cabe mencionar que la familia menos representada con respecto a la Sierra de Juárez fue Nymphalidae, sierra con características de vegetación y climas similares, por lo tanto se esperaba que el número de especies fuera similar, aunque la Sierra de Juárez abarca una extensión más amplia. La familia Hesperidae, no fue considerada en los otros trabajos, por lo que se necesitan más estudios con fines de comparación, pues muchos estudios de inventarios no incluyen a esta familia.
Tabla 5 Tabla comparativa de la riqueza de especiesy subespecies por familia con otras Sierras de México. Lycaenidae y Riodinidae, se presentan juntos para poder comparar los datos con los de otros trabajos. Esfuerzo en días de recolecta: Sierra Mazateca = 35 días efectivos; Sierra de Juárez = 98; Sierra de Atoyac = 116 y Sierra de Manantlán = 121 días. * En el total no se consideran las 141 taxones de Hesperidae registradas en este estudio.
| Papilionidae | Pieridae | Lycaenidae y Riodinidae | Nymphalidae | Total* | |
|---|---|---|---|---|---|
| Sierra Mazateca, Oaxaca (este estudio) | 19 | 42 | 90 | 168 | 319 |
| Sierra de Juárez, Oaxaca (Luis-Martínez et al. 1991) | 37 | 50 | 124 | 241 | 452 |
| Sierra de Atoyac, Guerrero (Vargas-Fernández et al. 1992) | 20 | 37 | 129 | 153 | 339 |
| Sierra de Manantlán, Jalisco y Colima (Vargas-Fernández et al. 1999) | 24 | 37 | 122 | 132 | 315 |
La localidad de Loma de Cacao fue la que presentó la mayor riqueza (con 236 taxones) para las seis familias incluidas en este trabajo, lo cual la coloca dentro de las localidades de Oaxaca con mayor riqueza comparada con Soyolapan el Bajo (243 taxones), Puerto Eligio (231 taxones) y Candelaria Loxicha (260 taxones).
Conclusiones
En la Sierra Mazateca se registra un total de 460 taxones de la superfamilia Papilionoidea. De ellas, 347 son nuevos registros para la Sierra Mazateca.
Se recolectó el 71.3% (380) de las especies-subespecies estimadas para la zona, con base en el modelo de Clench.
La riqueza en el gradiente altitudinal disminuye al aumentar la altitud a partir de los 900 msnm, lo cual coincide con otras sierras de México.
La localidad con mayor riqueza (236 especies) fue Loma de Cacao, a los 470 msnm.

