










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkActa zoológica mexicana
On-line version ISSN 2448-8445Print version ISSN 0065-1737
Acta Zool. Mex n.82 Xalapa Apr. 2001
Artículo
Distribución altitudinal de la mastofauna en la Reserva de la Biosfera "El Cielo", Tamaulipas, México
Jorge A. Vargas-Contreras1 y Arturo Hernández-Huerta2
1Instituto de Ecología y Alimentos, Universidad Autónoma de Tamaulipas, 13 Blvd. A. López Mateos No. 928, Cd. Victoria, Tamaulipas, CP 87040 MEXICO.
2Instituto de Ecología, A. C., Apdo. Postal 63, Xalapa, Veracruz, CP 91000 MEXICO.
Recibido: 15 de junio 2000
Aceptado: 14 de noviembre 2000
RESUMEN
La Reserva de la Biosfera "El Cielo" alberga uno de los acervos más importantes de la biodiversidad en el estado de Tamaulipas, México. En esta reserva se pueden encontrar cuatro tipos de vegetación (bosque tropical subcaducifolio, bosque mesófilo de montaña, bosque de encino-pino y matorral xerófilo), a los que están aunados una fauna característica. Este estudio determinó las especies de mamíferos asociadas a cada tipo de vegetación. Se detectó que los mamíferos presentan una distribución diferencial en un gradiente altitudinal en cinco patrones: cuatro especies de distribución amplia, siete especies de distribución media, 31 especies de distribución reducida, 12 especies de distribución salteada y 42 especies de distribución específica. Además, el bosque tropical subcaducifolio contiene el mayor número de especies con distribución específica, seguido por el bosque mesófilo de montaña, matorral xerófilo y bosque de encino-pino con diez, nueve y cuatro, respectivamente. Se empleó el índice porcentual de Simpson para calcular la similitud mastofaunística entre hábitats y se obtuvo que el bosque tropical y el bosque mesófilo presentan un mayor porcentaje (72.54%) de especies en común. En tanto que el bosque de encino pino y el matorral xerófilo comparten menos especies con otros hábitats y sus valores de similitud fluctúan de 28.57 a 58.33%. Así, los mamíferos de esta reserva están asociados en tres grupos: los del bosque tropical subcaducifolio-bosque mesófilo de montaña, los del bosque encino pino y los del matorral xerófilo.
Palabras clave: Gradiente altitudinal, mamíferos, patrones de distribución, Reserva de la Biosfera "El Cielo", Tamaulipas, México, similitud faunística.
ABSTRACT
The Biosphere Reserve "El Cielo" is one of the most important areas in Tamaulipas, Mexico, holding a rich biodiversity. This reserve embraces tropical semideciduous forest, cloud forest, pine-oak forest and dry scrub, which possess a unique fauna. Here, we determined which mammals were associated to each vegetation type. We observed five distributional patterns for mammals along an elevational gradient: four species with a wide distribution, seven species with a medium distribution, 31 species with a reduced distribution, 12 species with leap frog distribution, and 42 species with a specific distribution. Furthermore, tropical semideciduous forest have more species (19) with specific distribution that cloud forest (10), dry scrub (9), and pine-oak forest (4). Simpson's index was used to measure species similarity among habitats. A high percentage of shared species was found between tropical deciduous forest and cloud forest (72.54%), in comparison with other habitats; whose values varied between 28.57 and 58.33%. Meanwhile, pine-oak forest and dry scrub shared few species with other habitats and their values of similarity are between 28.57 and 58.33%. Thus, the Biosphere Reserve "El Cielo" mammals' are clustered in three groups: tropical semideciduous forest-cloud forest, pine-oak forest, and dry scrub mammals.
Key words: Altitudinal gradient, Biosphere Reserve "El Cielo", Tamaulipas, distribution patterns, mammals, Mexico, faunistic similarity.
INTRODUCCIÓN
La diversidad de especies se encuentra distribuida heterogéneamente. En muchos grupos, el número de especies disminuye al aumentar la latitud o la altitud (Pianka 1966, Kucera 1978, Humphrey y Bonaccorso 1979, Graham 1983). También, el paisaje puede jugar un papel importante; la complejidad del hábitat, determinada por una mayor variación ambiental, sea climática, topográfica o en tipos de vegetación, propicia que los hábitats heterogéneos contengan una mayor riqueza de especies, tanto florística como faunística, en comparación con aquellos hábitats homogéneos. Para describir los patrones espaciales de la biodiversidad, se han reconocido tres niveles principales de diversidad: diversidad α o número de especies en un área pequeña homogénea (riqueza puntual), diversidad β o tasa de recambio de especies entre hábitats contiguos y diversidad γ, que se refiere a la diversidad de ecosistemas en una región determinada (Orians 1994). Estas expresiones de la diversidad son particularmente conspicuas en los gradientes altitudinales.
Además, entre los mecanismos que promueven los patrones de distribución se mencionan el cambio de parámetros abióticos (temperatura, humedad, precipitación y altitud), la reducción del área efectiva, la complejidad y la productividad biológica de los hábitats, entre otros factores bióticos correlacionados (MacArthur y Wilson 1967, MacArthur 1972, Terborgh 1977, Graham 1983, Heaney et al. 1989, Rosenzweig 1992, Schluter y Ricklefs 1993, Tilman y Pacala 1993, Sánchez-Cordero en prensa). Sin embargo, los estudios sobre este tema son aún incipientes en México.
Entre los trabajos que resaltan la diversidad faunística en gradientes altitudinales en América se pueden mencionar los siguientes: sobre anfibios y reptiles (Martin 1955, 1958), aves (Terborgh 1971, Rotenberry 1978, Navarro 1992, Palomera et al. 1994, Gram y Faaborg 1997), roedores (Aguilar 1977, Santillán 1978, Barry et al. 1984), murciélagos (Humprey y Bonaccorso 1979, Graham 1983, León-Paniagua 1986), roedores y murciélagos (Sánchez-Cordero en prensa), y varios grupos de vertebrados (Martin 1955), entre otros. Dichos patrones pueden indicar la forma en que está conformada la diversidad en una determinada región; así, por ejemplo, en el caso de México, es la diversidad β la que permite que tenga mayor número de especies de mamíferos que Brasil (WCMC 1992, Arita 1993, Arita y León-Paniagua 1993), a pesar de ser un país 4.3 veces menos extenso.
En México el grupo de los mamíferos es diverso con aproximadamente 449 especies terrestres (Ceballos y Navarro 1991). Para el estado de Tamaulipas se han registrado el 31.40% de ese total (Ramírez-Pulido et al. 1983), concentrándose gran parte de esa riqueza en la porción suroeste del Estado, debido a la influencia de ambientes tropicales y a la complejidad orográfica de la Sierra Madre Oriental. La extraordinaria riqueza biológica presente en esta parte de Tamaulipas, especialmente en la región de Gómez Farías, ha motivado el interés de diversos investigadores. Este interés se refleja en un amplio historial de investigaciones para la región, que se remonta a principio de los 50's (Hernández-X. et al. 1951). En 1985 el gobierno del estado de Tamaulipas decretó la creación de la Reserva de la Biosfera "El Cielo", la cual oficialmente cubre una superficie de 144,530 ha, ayudando a proteger cerca del 60% de la región de Gómez Farías (Sosa et al. 1997).
En el área de la Reserva de la Biosfera "El Cielo", son pocos los trabajos que se han desarrollado sobre la distribución zonal de vertebrados. Martin (1955, 1958) determinó faunas de vertebrados asociadas al bosque mesófilo de montaña. Otros trabajos desarrollados en esta zona, que sólo mencionan las especies, el sitio de colecta, el hábitat o el número de especies, corresponden a Hooper (1953), De la Torre (1954), Álvarez (1963), Baker y López (1968), Reddell y Mitchel (1971), Martínez (1979), Hall (1981) y Hernández (1989).
Este estudio pretende determinar el grado de similitud mastofaunística a través de un gradiente altitudinal en la Reserva de la Biosfera "El Cielo", Tamaulipas, México, los patrones de distribución altitudinal y la asociación con la vegetación.
MATERIALES Y MÉTODOS
Área de estudio
La Reserva de la Biosfera "El Cielo" (RBC) se localiza al suroeste del estado de Tamaulipas (Fig. 1) y presenta cinco tipos de vegetación (Fig. 2; Sosa 1987): bosque tropical subcaducifolio (BTS, entre 200-800 m de altitud), que cuenta con dos estratos arbóreos, de los cuales el superior alcanza los 22 m y tiene como especies más comunes a Bursera simaruba (BURSERACEA), Brosimum alicastrum, Ficus spp. (MORACEA), Croton niveus (EUPHORBIACEA), Mirandaceltis monoica (ULMACEAE), Guazuma ulmifolia (STERCULIACEA) y Enterolobium cyclocarpum (MIMOSACEA); bosque mesófilo de montaña (BMM, 800-1,400 m), cuyo dosel puede llegar a los 30 m, y presenta tres subestratos arbóreos, teniendo como especies indicadoras a Liquidambar styraciflua (HAMAMILIDACEAE), Quercus sartorii, Q. germana (FAGACEAE), Podocarpus reichei (PODOCARPACEAE), Cercis canadensis (CAESALPINACEAE) y Acer skutchii (ACERACEAE); bosque de encino-pino (BEP, 1,400-1,800 m), ocupa una gran extensión en el área y tiene como especie más comunes a Q. sororia, Q. glaucenses (FAGACEAE), Pinus montezumae y P. teocote (PINACEAE); bosque de pino-encino (BPE, > 1,800 m), cuyos árboles más altos pueden superar los 20 m, y en el que dominan P. montezumae, P. patula, P. teocote (PINACEAE), Q. glaucenses (FAGACEAE) y Ostrya virginiana (BETULACEAE); y por último, el matorral xerófilo (MX, 1,300-1,600 m), en el que se combinan Quercus spp. (FAGACEAE), con Arbutos (ERICACEA), Yucca (AGAVACEAE), Cercocarpus (ROSACEAE) y Bauhinia (CAESALPINACEAE) (Martin 1958, Sosa 1987).
Los climas predominantes en estos tipos de vegetación son: semicálido-subhúmedo con lluvias en verano,(A)C(W1)(W), para el BTS; semicálido-húmedo, (A)C(m)(W), con abundantes lluvias en verano, para el BMM y el BEP; templado subhúmedo, C(W1), con lluvias en verano, para el BPE; y semiseco-semicálido con lluvias en verano para el MX (INEGI 1983, García 1988). La caracterización climática de la superficie que ocupan los tipos de vegetación, denota marcadamente las zonas cálidas y templadas como lo indica García (1988). Por otra parte, la RBC se ubica cerca de la zona transicional entre las regiones biogeográficas mayores, la neártica y la neotropical (Smith 1939).
Muestreo
Los registros de mamíferos fueron obtenidos de Nelson (1909), Goldman (1919), Miller (1924), Setzer (1949), Hooper (1953), Goodwin (1954, 1961), De la Torre (1954), Davis (1959), Packard (1960), Álvarez (1963), Anderson y Nelson (1965), Villa (1966), Baker y López (1968), Mollhagen (1971), LaVal (1973), Hall (1981) y Moreno-Valdez et al. (1997); de colectas de campo durante enero de 1990 a octubre de 1996 e información de bases de datos de colecciones mastozoológicas del Instituto de Biología de la Universidad Nacional Autónoma de México (IBUNAM), del Instituto de Ecología y Alimentos de la Universidad Autónoma de Tamaulipas (CM-IEA) y del Museo de Historia Natural de la Universidad de Kansas (MNHUK).
La toma de datos de campo comprendió 100 días. En el muestreo directo se emplearon 13 trampas "Tomahawk" de tamaño mediano en un transecto de aproximadamente 700 m (con 1,300 horas/trampas = 1,300 h/t): el BTS con 455 h/t, el BMM con 585 h/t y el BEP con 260 h/t; siete redes ornitológicas (2.13 x 12.81 m) (600 horas/red = 600 h/r): el BTS con 210 h/r, el BMM con 210 h/r, el BEP con 90 h/r y el MX con 90 h/r; 100 trampas "Sherman", cubriendo 1 ha (1,300 h/t): el BTS con 260 h/t, el BMM con 455 h/t, el BEP con 390 h/t y el MX con 195 h/t; recorridos por caminos y veredas para el avistamento y recolectas de mamíferos (un promedio de 300 h): el BTS con 90 h, el BMM con 90 h, el BEP con 60 h y el MX con 60 h. El tiempo que permanecieron abiertas las trampas son: las "Tomahawk" y "Sherman" de las 18:00 a 07:00 h, las redes de las 18:30 a 00:30 h y en los recorridos se invirtió tres horas en el día o la noche. Los ejemplares preparados y consultados están depositados en las colecciones mastozoológicas del IBUNAM, CM-IEA y MNHUK (Apéndice 2). Basándose en estos datos, se obtuvieron listados de las especies de mamíferos por hábitat (tipos de vegetación). El arreglo sistemático de las especies de mamíferos está basado en Ramírez-Pulido et al. (1996).
Con respecto a los tipos de vegetación que se han determinado para la reserva, no hay datos concluyentes. Diversos autores dan cifras distintas, Martin (1958) menciona ocho: matorral espinoso, matorral xerófilo, chaparral, bosque de pino-encino húmedo, bosque de encino-pino seco, bosque mesófilo de montaña, bosque tropical subcaducifolio y bosque tropical perennifolio-subperennifolio; Valiente (1984) cita cinco: selva mediana subperennifolia-perennifolia, selva mediana-baja subcaducifolia, selva baja caducifolia, bosque ripario y vegetación acuática; Sosa (1987) alude a otros cinco: matorral xerófilo, bosque de pino, bosque de encino-pino, bosque mesófilo de montaña y bosque tropical subcaducifolio; Suzán y Fragoso (1989) citan cuatro: matorral xerófilo, bosque de encino-pino, bosque mesófilo de montaña y bosque tropical subcaducifolio. Por último, Mora (1993) menciona cinco tipos de vegetación: bosque tropical subcaducifolio, bosque mesófilo de montaña, bosque de pino-encino, matorral xerófilo y vegetación acuática. Por tal razón y para fines prácticos de análisis de la diversidad de mamíferos en la RBC, seguimos a Suzán y Fragoso (1989) quienes consideran cuatro tipos de vegetación, sin discriminar entre distintas unidades de vegetación y ecotonos: bosque tropical subcaducifolio, bosque mesófilo de montaña, bosque de encino-pino y matorral xerófilo.
Para conocer la heterogeneidad faunística (diversidad beta) del área de estudio, es importante determinar el grado de similitud entre los hábitats (Hair 1987, Magurran 1988). Para esto se empleó el índice de similitud de Simpson, que es un índice porcentual y se basa en información binaria (ausencia-presencia). Su fórmula es: RN2 = 100 (s)/N2, donde s = número de taxa compartidos, N1 = número de taxa de fauna mayor y N2 = número de taxa de la fauna menor, interpretándose como la proporción de taxa compartidos en relación a la fauna más pequeña (Sánchez y López 1988). Se usó este índice porque es más sensible que otros a la presencia de especies compartidas y sus valores no dependen del tamaño de las faunas bajo estudio.
Se construyó una matriz de similitud que contiene información de riqueza total de especies por hábitat, número de especies compartidas y valores de similitud entre hábitats. El propósito de este análisis es verificar si los mamíferos voladores, no voladores, de talla pequeña (< 100 g) y de talla mediana a mayor (> 100 g) están formando grupos. Con el método de agrupamiento pareado no ponderado (Crisci y López 1983), se elaboraron los dendrogramas de similitud y con el valor crítico del 66.66% (Sánchez y López 1988) se determinó los posibles grupos mastofaunísticos de la RBC.
Con respecto a la división de mamíferos de talla pequeña y de talla mediana a grande, cabe mencionar que Robinson y Redford (1986) y Medellín (1994) consideran cuatro categorías basadas en la escala logarítmica de base 10: pequeños (< 100 g), medianos (entre 100 y 1,000 g), grande (entre 1,000 y 10,000 g) y muy grande (>10,000 g). Para nuestro análisis, sólo se consideran dos divisiones, pequeños (< 100 g) y medianos a grande (> 100 g), debido a que la fauna del segundo grupo es pequeño.
Con base en los registros sobre ausencia-presencia, en cada uno de los hábitats considerados, se determinaron los siguientes cinco patrones de distribución para las especies de mamíferos de la RBC. Distribución amplia, cuando una especie se encuentra a lo largo de todo el gradiente, desde los 200 a más de 1,800 m de altitud. Distribución media, cuando ocupa tres ambientes, de los 200 a 1,800 m o de los 800 a más de 1,800 m. Distribución reducida, cuando la especie sólo ocupa dos ambientes contiguos. Distribución específica, si las especies tienen distribución restringida a un solo hábitat. Distribución salteada, si la especie no está presente en hábitats adyacentes (Fig. 2).
RESULTADOS
Se registraron un total de 96 especies de mamíferos en la RBC (Apéndice 1) y siete más con distribución potencial que se menciona en la literatura para áreas cercanas. La riqueza de especies de mamíferos está representada en ocho órdenes, concentrándose el mayor número de especies en Chiroptera (44), Rodentia (28) y Carnívora (17) y, el menor número, en Xenarthra (1) y Lagomorpha (2) (Fig. 3).

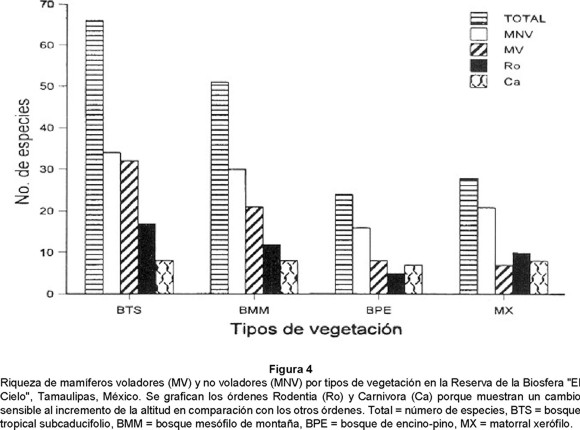
Las 96 especies con distribución real están representadas de la siguiente manera: 66 especies para el BTS, 51 para el BMM, 24 para el BEP y 28 para el MX (Fig. 4). En términos generales, la riqueza de mamíferos no voladores (59 especies) es mayor en un 13.46% que los mamíferos voladores (44), situación que se repite en cada tipo de vegetación (Fig. 4). En cuanto al tamaño, considerando la masa corporal, se encontró mayor riqueza de mamíferos de talla pequeña (62) que los de talla mediana a grande (34).
En esta zona, está representada aproximadamente el 75% de la riqueza mastofaunística de Tamaulipas, el 81.81% de las especies de murciélagos y el 67.81% de los mamíferos no voladores del Estado. En la Sierra de San Carlos, localizada a 152 km al norte de la RBC y el único otro lugar en Tamaulipas con información detallada sobre sus mamíferos, se han registrado 54 especies de mamíferos silvestres (Schmidly y Hendricks 1984); de ese total sólo 14 especies (cinco de quirópteros y nueve de no voladores) no están reportadas para la RBC. La mayoría de los mamíferos de esta sierra son especies de ambientes templados y desérticos, las cuales tienen una amplia distribución en el Estado.
El 81.25% (78 especies) de las especies reportadas para la RBC cuentan con verificación reciente de su presencia. Con respecto a los patrones de distribución, se tiene que el 56.25% de las especies están en más de dos hábitats, de las cuales Desmodus rotundus, Urocyon cinereoargenteus, Silvilagus floridanus y Peromyscus pectoralis, habitan todos los tipos de vegetación que aquí se citan. El 70.36% de este último porcentaje se distribuye en dos o tres hábitats y el 22.22% están en diferentes hábitats, pero no muestran un patrón definido. Es decir, siete especies están presente en tres hábitats adyacentes (patrón de distribución medio; BTS -BMM - BEP = Myotis californica, Peromyscus levipes, P. ochraventer, Mustela frenata y Panthera onca; BMM - BEP - MX = Puma concolor y Ursus americanus), 31 en dos hábitats (patrón de distribución reducido) y 12 se registraron en hábitat no contiguos (patrón de distribución salteado). Por otra parte, el 43.75% del total de especies se localizan en un sólo hábitat (patrón de distribución específico). De este último número, el BTS es el ambiente que presenta mayor número de especies exclusivas con 19, le siguen el BMM con 10 y el MX con nueve, en tanto que el BEP sólo presenta cuatro especies exclusivas (Cuadro 1, Apéndice 1).

Los hábitats con mayor número de especies compartidas, considerando la mastofauna en general, son el BTS y BMM con 72.54% de similitud. En cambio el MX es el hábitat con baja similitud con respecto a los otros, de 28.57 a 50% (Cuadro 2). Dado que la fauna de estos hábitats tienen una proporción de similitud menor al punto crítico (66.66%; Sánchez y López 1988), se puede considerar que en la zona de estudio existen tres grupos mastofaunísticos (BTS-BMM, BEP y MX) (Fig. 5).


Al analizar por separado los mamíferos voladores, no voladores, pequeños y medianos a grande, se encontró que la topología de los dendrogramas son diferentes. El caso de los mamíferos voladores es similar al obtenido cuando se analiza la fauna en general. Con respecto a los mamíferos no voladores y de talla pequeña, en el dendrograma se distinguen dos grupos (BTS-BMM-BEP y MX). Mientras que el dendrograma de los mamíferos medianos a grandes se observan cuatro grupos (BTS, BMM, BEP y MX) (Fig. 5).
Debido a que la RBC es un área protegida, la mayoría de los registros recientes sobre la presencia de algunas especies de mamíferos medianos y grandes, se basan en observaciones directas (avistamientos o capturas en trampas "Tomahawk") o evidencias indirectas, como rastros y excretas. Especies como el cabeza de viejo (Eira barbara), la comadreja (M. frenata), el tejón (Nasua narica) y el temazate (Mazama americana), se ha observado en estado silvestre. La zorra gris (U. cinereoargenteus), el tlacuachín (Marmosa mexicana), los tlacuaches mayores (Didelphis virginiana y D. marsupialis) y el conejo (S. floridanus), se capturaron en trampas para animales vivos. Los ataques al ganado y los rastros de sus pisadas conformaron las evidencias de la presencia del jaguar (P. onca); el oso negro (U. americanus) es frecuentemente observado por los pobladores locales, además, sus huellas y los daños que causa a los cultivos de maíz, principalmente en el otoño, indican que su presencia también es frecuente; los rastros y daños esporádicos causados al ganado indican la presencia del puma (P. concolor). Además, existen evidencias de que estas últimas especies de carnívoros, junto con la onza (Herpailurus yagouarondi), el tigrillo (Leopardus wiedii) y el cabeza de viejo (E. barbara), han sido cazados recientemente en la región. Para el caso de mamíferos de tallas pequeñas, particularmente murciélagos, hemos capturado entre los más raros a Choeronycterys mexicana, Euderma phyllote que no había sido reportado previamente en el área de estudio, Nycticeius humeralis es una especie fácil de identificar en el campo y por ello se liberó, Corynorhinus mexicanus y Antrozous pallidus.
DISCUSIÓN
Los trabajos realizados por Álvarez (1963) y Arita (1993), mencionan la distribución real y/o potencial de los mamíferos de Tamaulipas; reportando 120 y 139 especies, respectivamente. Hernández (1989) analizó la riqueza de mamíferos en la RBC, proporcionando un listado de 92 especies. En este artículo se informa de 96 especies registradas y otras siete que probablemente ocurren o estuvieron presentes en la región, 12 más de las mencionadas por Hernández (1989). Considerando los cambios taxonómicos que se han sugerido para el complejo Peromyscus boylii (Houseal et al. 1987), y con base en su área de distribución, se reconoce en la reserva la presencia de Peromyscus levipes que anteriormente estaba contemplada como subespecie de P. boylii (Carleton 1989, Musser y Carleton 1993).
Como se mencionó anteriormente los tipos de vegetación se ubican a diferentes altitudes, y aparentemente el MX se sobrepone altitudinalmente entre el BMM y el BEP. Considerando que en ocasiones resulta difícil delimitar la frontera entre tipos de vegetación contiguos, y que ciertas zonas de la RBC han sido poco estudiadas, algunos de los mamíferos que presentan el patrón de distribución salteado, no continuo entre hábitats adyacentes, podrían al mismo tiempo tener una distribución geográfica continua, p. ej., Micronycteris megalotis, Dermanura azteca, Antrozous pallidus, Pipistrellus hesperus, P. subflavus, Myotis auriculacea, Liomys irroratus, Baiomys taylori, Peromyscus leucopus, Reithrodontomys fulvescens, Sigmodon hispidus y Odocoileus virginianus. Evidentemente los porcentajes sobre el número de especies por cada patrón de distribución pueden variar al considerar a Lasiurus ega, Nyctinomops laticaudata, N. macrotis, Perognathus flavus, Neotoma micropus, Leopardus pardalis y Odocoileus hemionus, las cuales han sido reportadas en la literatura, pero no observadas durante los muestreos de campo.El hecho de que algunas especies muestren un patrón de distribución salteado y el que especies con distribución amplia exhiban un patrón específico (e. g., D. virginiana, Tadarida brasiliensis, Canis latrans y Sciurus aureogaster), posiblemente se deba a que las áreas poco accesibles no han sido muestreadas suficientemente o que tales especies tengan abundancia muy bajas localmente.
Pianka (1966), Fleming (1973), Kucera (1978), Humphrey y Bonaccorso (1979), Graham (1983) y Sánchez-Cordero (en prensa), han mencionado que existe mayor riqueza de especies en los hábitats tropicales que en los templados. Los resultados de este estudio coinciden con esta idea generalizada, que se denota en los totales de especies por hábitats (Fig. 4). Un estudio realizado por Vargas-Contreras (1998) sobre murciélagos cavernícolas en un gradiente altitudinal en la RBC en Tamaulipas, también corrobora la idea anterior, encontrando la mayor diversidad de murciélagos en la porción tropical (56.25%) y un decremento hacia la zona templada (43.7%).
Aunque el análisis de la variación de la riqueza de mamíferos se hace a una escala espacial menor, se distingue la contribución desmesurada de los murciélagos (43.27%) al gradiente trópico-templado y su disminución conforme aumenta la altitud. Además, la riqueza de quirópteros es mayor en la parte tropical y el número de especies insectívoras (17), frugívoras (nueve), nectarívoras (tres) y hematófagas (tres) disminuye en los ambientes templados (cuatro, uno, cero y uno, respectivamente). Asimismo, encontramos especies que desaparecen conforme aumenta la altitud, o cambia el hábitat, de 11 a dos, como es el caso de Mormoops megalophylla, Pteronotus parnellii, P. personatus, Natalus stramineus, Molossus molossus y M. rufus para el BTS, Anoura geoffroyi, Lasiurus cinereus, N. humeralis y T. brasiliensis para el BMM, C. mexicanus y Myotis velifera para el BEP, Macrotus californicus y E. phyllote para el MX. En este último punto, cabe resaltar que algunas especies que hallamos pueden encontrarse en otros ambientes de menor altitud y por la forma de analizar nuestros datos, se deben tomar con mucha cautela.
Para otros grupos, como los roedores y carnívoros, existe un ligero incremento en el número de especies (Fig. 4). Esta situación es similar a la mencionada por Fleming (1973), Wilson (1974), McCoy y Connor (1980), Heaney et al. (1989), y Ceballos y Navarro (1991) para la variación del número de especies de mamíferos en el gradiente latitudinal. Para nuestro caso, la contribución de los quirópteros y mamíferos no voladores en el BTS es relativamente similar, reduciéndose en el BMM, BPE y MX; en el otro extremo, se tiene que hay un relativo aumento de la riqueza de especies de roedores y carnívoros en comparación con los quirópteros en los ambientes templados secos (MX; Fig. 4). Este patrón de distribución altitudinal coincide con otros estudios (Graham 1983, Heaney et al. 1989, Robertson 1975).
La RBC se localiza entre la zona transicional de dos regiones biogeográficas, lo cual le confiere una riqueza y recambio de especies de afinidad neártica y neotropical; concentrándose en la zona cálida la mayor diversidad de mamíferos (66 especies). Esto se refleja en las diferencias de los dendrogramas del análisis de similitud mastofanística entre hábitats, del cual se obtienen tres situaciones. (1) Un grupo está constituido principalmente por especies de afinidad tropical que habitan en el BTS y BMM, en tanto que el segundo (BEP) y el tercero (MX) su fauna está constituida en proporciones similares de taxa de afinidad neártica y neotropical. Además, los mamíferos voladores presentan el mismo patrón que la mastofauna general en el análisis de similitud faunística. (2) Los mamíferos no voladores y de talla pequeña forman dos asociaciones: la del BTS-BMM-BEP y la de MX. (3) En los mamíferos de talla mediana a grande hay diferencias significativas entre los cuatro hábitats; es decir, se unen por abajo del 64%, de esta manera cada hábitat contiene un grupo de mamíferos particular. Para todos los casos, las especies de mamíferos que habitan en el BTS y BMM siempre forman un grupo, en cambio los que se encuentran en el BEP y MX varían en el grado de similitud con respecto a los hábitats antes mencionados.
Las situaciones previamente mencionadas pueden interpretarse de la siguiente manera: en los casos 1 y 2 los mamíferos pueden estar respondiendo a factores abióticos (e. g. altitud, temperatura, precipitación, etc.); en tanto que en el caso 3, la distribución de los mamíferos medianos a grandes podría variar de acuerdo a la heterogeneidad de hábitats. Estas interpretaciones coinciden con las mencionadas por Arita (1993).
Considerando diversos estudios, la RBC figura como el área con mayor riqueza de mamíferos en Tamaulipas (Baker 1951; Hall 1951; Álvarez 1963; Baker y Webb 1966; Schmidly y Hendricks 1984) y aproximadamente una quinta parte de esta mastofauna se encuentra actualmente protegida (Cuadro 3). Entre esas especies se encuentran las endémicas para México como C. mexicanus, Sciurus alleni, L. irruratus alleni y Chaetodipus nelsoni nelsoni, y las endémicas a la zona tres, que incluye a la parte del noreste del país, como Neotoma angustapalata y P. ochraventer (Ramírez-Pulido y Mudespacher 1987, Fa y Morales 1993). Con respecto a las especies amenazadas se menciona a P. concolor, M. americana y Agouti paca nelsoni (Ceballos y Navarro 1991).

Por otra parte, el 30% de la riqueza mastozoológica en la RBC la aportan especies que están restringidas al BTS y BMM, pero en general presentan el 63% entre especies restringidas y compartidas, de aquí la importancia de conservar estos ambientes, no sólo por lo que significa para la reserva sino también para la fauna de mamíferos de Tamaulipas. En la RBC el MX no es un ambiente particularmente rico en especies, pero en este habitan nueve especies que han podido ser registradas sólo ahí. El BTS es el ambiente con mayor riqueza específica de mamíferos en la reserva y es también, junto con el BMM, uno de los ambientes que sustenta mayor número de especies vulnerables (12). Desafortunadamente también es el tipo de vegetación con mayor presión en la actualidad debido a que su cubierta vegetal está siendo transformada para establecer cultivos. Los mayores asentamientos humanos dentro de la RBC están ubicados en el BTS, donde vive cerca del 80% del total de la población de la reserva. No obstante, la mayor amenaza consiste en la reciente expansión del monocultivo del nopal (Opuntia sp.), ya que ello implica la eliminación de la vegetación natural en su totalidad y la transformación de las áreas de policultivos tradicionales, complejos y diversos, a sistemas extremadamente simples.
CONCLUSIONES
Nuestra evaluación indica que del total de especies registradas para el área de la RBC, actualmente están presentes por lo menos el 81.25% de ellas, incluyendo casi todas las especies de mamíferos de talla mediana y grande. La RBC es la única área protegida donde el oso negro (U. americanus) y el jaguar (P. onca) se encuentran juntos, coincidiendo en el BMM, que sirve como hábitat marginal, ya que ambas especies son más abundates en otros hábitats. De hecho existen conflictos por los daños que estas dos especies y el puma (P. concolor) ocasionan a los lugareños. Cabe destacar que si bien la riqueza específica de mamíferos en la RBC es alta, la abundancia relativa de la mayoría de las especies es baja, probablemente como resultado de la intensa explotación forestal que se realizó en la zona hasta mediados de los 70's y un gran incendio, ocurrió en el invierno de los 70's, que desbastó gran parte de los bosques de la reserva. De los siete taxa endémicos a México y que se distribuyen en la RBC, dos (Corynorhinus mexicanus y Sciurus alleni) únicamente se han registrado en el BPE, uno (Neotoma angustapalata) en el BMM y una subespecie (Chaetodipus nelsoni nelsoni) en el MX. Cabe señalar que existen escasos registros para estos taxa en la región y por ello ameritan atención especial.
La RBC alberga uno de los acervos más importantes de la biodiversidad en Tamaulipas. Gran parte de esa riqueza le confieren los elementos asociados a los ambientes tropicales. En el caso de los mamíferos, casi una tercera parte de las especies que viven en la RBC tienen una distribución restringida al BTS y BMM, dos ambientes que han quedado relativamente aislados, debido a las transformaciones que ha sufrido el paisaje que rodea a la reserva. Esto implica que las especies de distribución restringida a los cuatro hábitats aquí tratados tengan una probabilidad alta de extinción local. Ante esta situación es imprescindible evaluar las posibilidades de lograr una continuidad ambiental para facilitar la conectividad no solamente entre poblaciones de mamíferos que se albergan en la región de la reserva, sino también las que habitan en ambientes circundantes. Esto también implica definir los patrones espacio-temporales en la abundancia de las poblaciones de mamíferos, así como el estudio de sus interacciones intra-interespecíficas a fin de proponer estrategias para mitigar los conflictos con los pobladores locales, específicamente aquellos relacionados con los mamíferos carnívoros.
AGRADECIMIENTOS
Agradecemos a Rafael Herrera, Livia León, Joaquín Arroyo, Griselda Escalona, Víctor Sánchez-Cordero, Miguel Ortega, Arturo Mora, Vinicio Sosa, Fátima Díaz y a dos revisores anónimos por sus comentarios que aportaron mejoras sustanciales al manuscrito. A Santiago Niño y Juan Córdoba por datos proporcionados. A Fernando Cervantes del IBUNAM y Robert Timm del MNHUK por permitirnos revisar las bases de datos y especímenes de las respectivas colecciones de mamíferos. A ECOSUR-Unidad Campeche por el apoyo logístico para la culminación de este documento.
LITERATURA CITADA
Aguilar, R.V.M. 1977. Distribución altitudinal de las tuzas (Rodentia: Geomydae) en la vertiente oriental del monte Iztlacihuatl. Tesis Profesional. Escuela de Ciencias Biológicas, Universidad del estado de Morelos. 60 pp. [ Links ]
Álvarez, T. 1963. The recent mammals of Tamaulipas, Mexico. Univ. Kansas Publ., Mus. Nat. Hist. 14: 363-473. [ Links ]
Anderson, S. & C.E. Nelson. 1965. A systematic revision of Macrotus (Chiroptera). Amer. Nat. Mus. Novitates 2212:1-39. [ Links ]
Arita, H.T. 1993. Riqueza de especies de la mastofauna de México. Pp. 109-128. In: Medellín, R.A. y G. Ceballos (eds.). Avances en el estdio de los mamíferos de México. Publ. Esp. Vol. 1. Asociación Mexicana de Mastozoología, A.C. [ Links ]
Arita, H.T. & L. León-Paniagua. 1993. Diversidad de mamíferos terrestres. Ciencias, No. Especial 7:12-22. [ Links ]
Baker, R.H. 1951. Mammals from Tamaulipas, Mexico. Univ. Kansas Publ., Mus. Nat. Hist. 5:207-218. [ Links ]
Baker, R.J. & G. López. 1968. Notes on some bats of Tamaulipas. Southw. Nat. 13:361-362. [ Links ]
Baker, R.H. & R.G. Webb. 1966. Notas de acerca de los anfibios, reptiles y mamíferos de La Pesca, Tamaulipas. Rev. Soc. Mex. Hist. Nat. 27:179-190. [ Links ]
Barry, R.E., Jr., M.A. Botaje & L.B. Gratham. 1984. Vertical stratification of Peromyscus leucopus and P. maniculatus in southwestern Virginia. J. Mamm. 65:145-148. [ Links ]
Carleton, M.D. 1989. Systematic and evolution. Pp. 7-141. In: Kirkland, Jr., G.L. & J.N. Layne (eds.). Advances in the study of Peromyscus (Rodentia). Texas Tech University Press, Lubbock, Texas. [ Links ]
Ceballos, G. & D. Navarro L. 1991. Diversity and conservation of Mexican mammals. Pp. 167-198. In: Mares, M.A. & D.T. Schmidly (eds.). Latin American mammalogy: history, biodiversity and conservation. Oklahoma University Press, Oklahoma. [ Links ]
Crisci, J.V. & M.F. López A. 1983. Introducción a la taxonomía numérica. Secretaría General de la Organización de los Estados Americanos Programa Regional de Desarrollo Científico y Tecnológico. Serie de Biología, Monografía No. 26. [ Links ]
Davis, W.H. 1959. Taxonomy of the Eastern Pipistrel. J. Mamm. 40:521-531. [ Links ]
De la Torre, L. 1954. Bats from southern Tamaulipas, Mexico. J. Mamm. 35:113-116. [ Links ]
Fa, J.A. & L.M. Morales. 1993. Patterns of mammalian diversity in Mexico. Pp. 319-361. In: Ramamoorthy, T.P., R. Bye, A. Lot, and J. Fa (eds.). Biological Diversity of Mexico, origin and distribution. Oxford University Press. [ Links ]
Fleming, T.H. 1973. Numbers of mammal species in North and Central American forest communities. Ecology 54:555-563. [ Links ]
García, E. 1988. Modificación al sistema de clasificación climática de Köppen. Instituto de Geografía, UNAM. México, D.F. 217 pp. [ Links ]
Goldman, E.A. 1919. Revision of the wood rats of the genus Neotoma. N. Amer. Fauna 31:1-124. [ Links ]
Goodwin, G.G. 1954. Mammals from Mexico collected by Miriam Martin for the American Museum of Natural History. Amer. Nat. Mus. Novitates 1689:1-16. [ Links ]
---------- 1961. Flying Squirrel (Glaucomys volans) of Middle America. Amer. Nat. Mus. Novitates 2059:1-22. [ Links ]
Graham, G.L. 1983. Changes in bat species diversity along an elevational gradient up The Peruvian Andes. J. Mamm. 64:559-571. [ Links ]
Gram, W.K. & J. Faaborg. 1997. The distribution of Neotropical migrant birds wintering in the El Cielo Biosphere Reserve, Tamaulipas, Mexico. Condor 99:658-670. [ Links ]
Hair, J.D. 1987. Medida de la diversidad ecológica. Pp. 283-290. In: Mosby, H.S., R.H. Gile Jr. y S.D. Schemnitz (eds.). Manual de Técnicas de Gestión de Vida Silvestre. The Wildlife Society. USA. [ Links ]
Hall, E. R. 1981. The Mammals of North America. John Wiley & Sons. 2nd. ed. U.S.A. 1811 pp. [ Links ]
Heaney, L.R., P.D. Heideman, E.A. Rickart, R.B. Utzurrum & J.S.H. Klompen. 1989. Elevational zonation of mammals in the Central Philippines. J. Tropical Ecol. 5:259-280. [ Links ]
Hernández H., A. 1989. Importancia de la Reserva "El Cielo" para los mamíferos de Tamaulipas. BIOTAM 1:13-20. [ Links ]
Hernández-X., E., H. Crum, W.B. Fox & A.J. Sharp. 1951. A unique vegetation area in Tamaulipas. Bull. Torrey Bot. Club 78(6): 458-463. [ Links ]
Hooper, E.T. 1953. Notes on mammals of Tamaulipas, Mexico. Occas. Papers Mus. Zool. Univ. Michigan 544:1-12. [ Links ]
Houseal, T.W., I.F. Greenbaum, D.T. Schmidly, S.A. Smith & K.M. Davis. 1987. Karyotipic variation in Peromyscus boylii from Mexico. J. Mamm. 68(2):281-296. [ Links ]
Humphrey, S.R. & F.J. Bonaccorso. 1979. Population and community ecology. In: Baker, R.J., J.K. Jons, Jr., and C.D. Carter (eds.). Biology of bats of the New World family Phyllostomidae. Part.III. Spec. Publ. Mus. Texas Tech Univ. No. 16:409-441. [ Links ]
INEGI. 1983. Síntesis geográfica de Tamaulipas. Inst. Nal. Estad. Geog. e Informática. Sría. Programación y Presupuesto. México, D.F. 13 mapas escala 1:100000. [ Links ]
Kucera, C.L. 1978. The challenge of ecology. 2nd. ed. The C.V. Mosby Company. U.S.A. 326 pp. [ Links ]
LaVal, R.K. 1973. A revision of the Neotropical bats of the genus Myotis. Nat. Hist. Mus Los Angeles Co. Sci. Bull. 15:1-54. [ Links ]
León-Paniagua, L.S. 1986. Distribución altitudinal de los murciélagos en el NE del Estado de Querétaro. Tesis Profesional. Facultad de Ciencias, UNAM. México, D.F. 73 pp. [ Links ]
MacArthur, R.H. 1972. Geographical Ecology: patterns in the distribution of species. Harper and Row, New York. [ Links ]
MacArthur, R.H. & E.O. Wilson. 1967. The Theory of Island Biogeography. Princeton University Press, Princeton. [ Links ]
Magurran, A.E. 1988. Ecological diversity and its measurement. Princeton University Press Great Britain. 179 pp. [ Links ]
Martin, P.S. 1955. Zonal distribution of vertebrates in a Mexican cloud forest. Amer. Nat. 89:347-361. [ Links ]
---------- 1958. A biogeography of reptiles and amphibian in the Gomez Farias region, Tamaulipas, Mexico, Misc. Publ. Mus. Zool. Univ. Michigan 101:1-102. [ Links ]
Martínez B., J.A. 1979. Notas sobre los murciélagos de Gómez Farías, Tamaulipas, México. Tesis Profesional. Facultad de Ciencias Biológicas, Universidad Autónoma de Nuevo León, México. [ Links ]
McCoy, E.D. & E.F. Connor. 1980. Latitudinal gradients in the species diversity of North American mammals. Evolution 34:193-203. [ Links ]
Miller, G.S., Jr. 1924. List of North American recent mammals, 1923. Bull. U.S. Nat. Mus. 128:1-673. [ Links ]
Medellín, R.A. 1994. Mammal diversity and conservation in the Selva Lacandona, Chiapas, Mexico. Cons. Biol. 8:780-799. [ Links ]
Mollhagen, T. 1971. Checklist of bats in caves in the regions of the Sierra de Guatemala and Sierra de El Abra, northeastern Mexico. Assoc. Mexican Cave Studies, Bull. 4:19-22. [ Links ]
Mora O., A. 1993. Aspectos botánicos de la reserva de la biosfera "El Cielo". Pp. 45-50. In: Ruíz C., E. (ed.). Memorias del Segundo Curso Internacional sobre Entomología Tropical en la Reserva de la Biosfera "El Cielo", Tamaulipas, México. Universidad Autónoma de Tamaulipas. [ Links ]
Moreno-Valdez, A., P.A. Lavín-Murcio & O.M. Hinojosa-Falcón. 1997. El Tepezcuintle, Agouti paca (Rodentia:Agoutidae), en Tamaulipas, México. Revista Mexicana de mastozoología, 2:129-131. [ Links ]
Musser, G.G. & D. Carleton. 1993. Family Muridae. Pp. 501-755. In: Wilson, D.E. & D.M. Reeder (eds.). Mammals species of the world, a taxonomy and geographic reference. Smithsonian Institution Press. [ Links ]
Navarro, A. 1992. Altitudinal distribution of bird in the Sierra Madre del Sur, Guerrero, Mexico. Condor 94:29-39. [ Links ]
Nelson, E.W. 1909. The rabbits of North America. N. Amer. Fauna 29:1-314. [ Links ]
Orians, G.H. 1994. Global biodiversity I. Patterns and processes. Pp. 78-110. In: (Meffe, G.K., C.R. Carrol and Contributors). Principles of conservation biology. Sinauer Associates. Inc. Publishers, Sunderland, Massachusetts. [ Links ]
Packard, R.L. 1960. Speciation and evolution of the Pygmy Mice, genus Baiomys. Univ. Kansas publ., Mus. Nat. Hist. 9:579-670. [ Links ]
Palomera G., C., E. Santana C. & R. Amparán S. 1994. Patrones de distribución de la avifauna en tres estados del occidente de México. An. Inst. Biol. Univ. Nal. Autón. México. Ser. Zool. 65(1):137-175. [ Links ]
Pianka, E.R. 1966. Latitudinal gradients in species diversity: a review of concepts. Amer. Nat. 100:33-46. [ Links ]
Ramírez P., J., A. Castro C., J. Arroyo C. & F. Cervantes R. 1996. Lista taxonómica de los mamíferos terrestres de México. Occas. Paper Mus. Texas Tech Univ. 158:1-62. [ Links ]
Ramírez P., J., R. López W., C. Müdespacher Z. & I.E. Lira. 1983. Lista y bibliografía reciente de los mamíferos de México. U.A.M.-Iztapalapa. México. 362 pp. [ Links ]
Ramírez P., J. & C. Müdespacher. 1987. Estado actual y perspectivas del conocimiento de los mamíferos de México. Ciencia 38:49-67. [ Links ]
Reddell, J.R. & R.W. Mitchell. 1971. A checklist of cave fauna of Mexico. II. Sierra de Guatemala, Tamaulipas. Assoc. Mexican Cave Studies, Bull. 4:181-215. [ Links ]
Robertson, P.B. 1975. Reproduction and community structure of rodents over a transect in Southern Mexico. Unpublised Ph.D. dissertation, University of Kansas. 113 pp. [ Links ]
Robinson, J.G. & K.H. Redford. 1986. Body size, diet, and population density of Neotropical forest mammals. Amer. Nat. 128:665-680. [ Links ]
Rotenberry, J.T. 1978. Components of avian diversity along a multifactorial climatic gradient. Ecology 59:693-699. [ Links ]
Rosenzweig, M.L. 1992. Species diversity gradients: we know more and less than we thought. J. Mamm. 73:715-730. [ Links ]
Sánchez, O. & G. López. 1988. A theoretical analysis of some indices of similarity as applied to biogeography. Folia Entomol. Mex. 75: 119-145. [ Links ]
Sánchez-Cordero, V. (en prensa). Elevation gradients of diversity for bats and rodents in Oaxaca, Mexico. Global Ecology and Biogeography. [ Links ]
Santillán A., S. 1978. Distribución altitudinal de roedores en el campo experimental forestal "San Juan Tetla" Estado de Puebla, México. Tesis Profesional. Escuela de Ciencias Biológicas, Universidad Autónoma del estado de Morelos. 178 pp. [ Links ]
Schluter, D. & R.E. Ricklefs, 1993. Species diversity: an introduction to the problem. Pp. 1-12. In: Ricklefs, R.E. & D. Schluter (eds.). Species diversity in ecological communities. Historical and Geographical Perspectives. The Chicago University Press, Chicago and London. [ Links ]
Schmidly, D.J. & F.S. Hendricks. 1984. Mammals of the San Carlos Mountains of Tamaulipas, Mexico. In: Martin, R.E.& R. Chapman (eds.). Contributions in mammalogy in honor of Robert L. Packard. Special Publ. Mus. Texas Tech Univ., Lubbock, Texas. No. 22:15-69. [ Links ]
SEMARNAP. 1994. NOM-059-ECOL-1994, que determina las especies, subespecies de flora y fauna silvestre terrestres acuáticas en peligro de extinción, amenazadas, raras y las sujetas a protección especial y que establece especificaciones para su protección. Gaceta Ecológica. Secretaría de Medio Ambiente, Recursos Naturales y Pesca. VII (33):1-72. [ Links ]
Setzer, H.S. 1949. Subspeciation in the Kangaroo Rat Dipodomys ordii. Univ. Kansas Publ., Mus. Nat. Hist. 12:309-375. [ Links ]
Smith, H.M. 1939. Las provincias bióticas de México, según la distribución geográfica de lagartijas del género Sceloporus. An. Esc. Nal. Cienc. Biol. IPN. México. 2:103-110. [ Links ]
Sosa, V.J. 1987. Generalidades de la región de Gómez Farías. Pp. 15-28. En: Puig, H. y R. Bracho (eds.). El bosque mesófilo de montaña de Tamaulipas. Instituto de Ecología, A.C. México, D.F. [ Links ]
Sosa, V.J., A. Hernández & A. Contreras. 1997. Gómez Farías region and El Cielo biosphere reserve, Mexico. Pp. 161-166. In: Davis, S.D., V.H. Heywood, O. Herrera-MacBrady, J. Villalobos & A.C. Hamilton (eds.). Centres of plant diversity: a guide and strategy for their conservation. WWF-IUCN. Publications Unit. Cambridge. U.K. [ Links ]
Suzán, H. & C. Fragoso. 1989. Investigación y conservación en la Reserva de la Biosfera "El Cielo", Tamaulipas, México. BIOTAM 1:48-53. [ Links ]
Terborgh, J. 1971. Distribution on environmental gradients: theory and a preliminary interpretation of distributional patterns in the avifauna of the Cordillera Vilcabamba, Peru. Ecology 52:23-40. [ Links ]
Tilman, D. & S. Pacala. 1993. The maintenance of species richness in plant communities. Pp. 13-25. In: Ricklefs, R.E. & D. Schluter (eds.). Species diversity in ecological communities. Historical and Geographical Perspectives. The Chicago University Press, Chicago and London. [ Links ]
Valiente B., A. 1984. Análisis de vegetación de la región de Gómez Farías, Tamaulipas. Tesis Profesional. Facultad de Ciencias, UNAM. 63 pp. [ Links ]
Vargas-Contreras, J.A. 1998. Factores microclimáticos y selección del refugio diurno por murciélagos cavernícolas en Gómez Farías, México. Tesis de Maestría en Ciencias (Biología Animal) de la Facultad de Ciencias. UNAM. 110 pp. [ Links ]
Villa, B. 1966. Los murciélagos de México. Instituto de Biología, Univ. Nal. Autón. Méx. 491 pp. [ Links ]
Wilson, J.W. III. 1974. Analytical zoogeography of North American mammals. Evolution 28:124-140. [ Links ]
World Conservation Monitoring Centre. 1992. Global biodiversity: status of the Earth's living resources. Chapman & Hall, London. 594 pp. [ Links ]

