











 text new page (beta)
text new page (beta) Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO 
 Permalink
Permalink

ANTECEDENTES
La preeclampsia (PE) es una patología con alta morbimortalidad a nivel mundial1,2, esta entidad fisiológica fue descrita desde la primera mitad del siglo XVII3 y, sin embargo, se desconoce su etiología. Gracias al estudio intenso de la PE se comprenden muchos de sus aspectos fisiopatológicos, por lo que se ha mejorado el manejo terapéutico, así como el diagnóstico temprano y la disminución de secuelas. Desde muy temprano en el estudio de la PE se reconoció que la inflamación, así como la activación del sistema inmunológico participaban en el origen y perpetuación de la enfermedad. Se ha determinado que la placenta es uno de los órganos de choque donde la inflamación y la respuesta inmunológica generan el daño y que se traduce en el cuadro clínico característico, es por esto que los mecanismos celulares y humorales de la respuesta inmunológica podrían ser blancos terapéuticos para tratar la PE. El presente artículo describirá la participación de elementos de la respuesta inmunológica que participan en la fisiopatología de la PE. Además, se mencionarán los posibles blancos terapéuticos para la PE.

El daño en el endotelio como origen de la PE
Los tres signos clásicos que definen la PE (hipertensión, proteinuria y edema) indican que el sistema vascular está involucrado en la fisiopatología de la enfermedad, y quizá también participa en su etiopatogenia. No sabemos si los cambios vasculares en placenta desencadenan la desregulación de los sistemas, o si es la desregulación de esos mismos sistemas lo que afecta el desarrollo fisiológico vascular en placenta y resulta en la PE. Hay reportes que muestran defectos de angiogénesis en pacientes con PE lo que apoyaría la tesis de que la PE es la consecuencia de la primigenia afectación vascular, sin embargo, también se han reportado defectos en la coagulación o del complemento que afectan negativamente la función vascular y que finalmente se expresarían como PE. Por lo que, lo que denominamos como PE puede ser la expresión patológica de origen diverso, donde en algunos casos:
Fue una organogénesis y función vascular alterada (especialmente en placenta) lo que provocó el desarrollo de PE.
Fue la patología en otros sistemas que afecto la función vascular y favorece la perpetuación de la PE.
O fue una participación de ambos mecanismos.
El sistema inmunológico está altamente regulado en el embarazo, a esta regulación la denominamos “tolerancia inmunológica del embarazo”. Desde el principio del estudio de la tolerancia en el embarazo se evaluó la participación de la placenta como elemento regulador, y más recientemente la participación de componentes celulares y humorales de la respuesta innata en el embarazo. Varios son los mecanismos que establecen y mantienen la tolerancia en el embarazo fisiológico o que por contraste podrían estar desregulados en la PE. El análisis clínico y el tiempo en el que sucede la fisiopatología de PE, nos ha permitido distinguir dos tipos de presentación:
La que sucede antes de la 34ª semana de gestación (SDG) que denominamos PE de inicio temprano (EOPE, por sus siglas en inglés).
La que sucede después de la 34ª SDG que denominamos PE de inicio tardío (LOPE, por sus siglas en inglés).
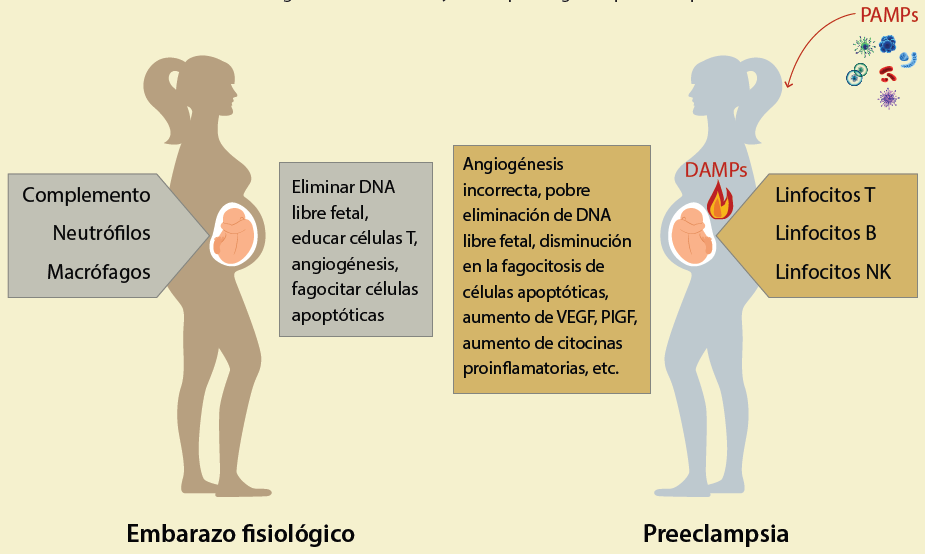
En ambos escenarios clínicos los mecanismos inmunológicos operan favoreciendo el establecimiento del estado patológico. El presente artículo comentará sobre algunos de los mecanismos de la respuesta innata y adquirida que son afectados en el estado de PE (figura 1), aunque sin hacer la distinción entre inicio temprano o tardío.

Elementos solubles y celulares de las respuestas innata y adaptativa participan coordinadamente para establecer y mantener la tolerancia inmunológica, o son desregulados por efecto de DAMPs o PAMPs lo que permite el establecimiento de la preeclampsia.
Imagen vectorial: Freepik
Figura 1 Componentes de respuesta inmunológica que participan en la “tolerancia inmunológica del embarazo” y la fisiopatología en preeclampsia
El sistema inmunológico tiene respuestas innatas como adquiridas, ambas respuestas actúan a diferentes tiempos durante la gestación; además, se han descrito tanto elementos celulares como humorales en estas respuestas4. Los mecanismos inmunológicos que mantienen la tolerancia inmunológica del embarazo son redundantes y operan con intensidad variable a lo largo del embarazo, sin embargo, la falla o limitación de estos mecanismos de tolerancia en la PE favorece el desarrollo de la patología, y puede resultar en desenlaces fatales para el binomio feto-materno.
Varios son los componentes celulares y humorales de la respuesta innata y adquirida que han sido estudiados en pacientes con PE y se ha determinado que su participación es decisiva para esta enfermedad. Se puede suponer que la regulación tanto de las respuestas “viejas” evolutivamente (respuesta innata) como las más “recientes” en la evolución (respuesta adquirida) son necesarias para evitar el desarrollo de PE. Se ha propuesto que células de la respuesta innata pueden participar muy temprano en la remodelación e invasión trofoblástica de tejido placentario, permitiendo el desarrollo estructural placentario fisiológico o en su defecto favorecer el desarrollo anómalo y el posterior estado fisiopatológico de la también llamada “toxemia del embarazo”. En contraste, la participación de la respuesta adquirida parece tener efectos menos estructurales y más funcional/regulador que observamos durante el desarrollo normal en la gestación. Así que la participación de la respuesta inmunológica en PE es la suma de los defectos en la respuesta innata como de la adquirida. La inadecuada respuesta inmunológica en PE afecta negativamente el desarrollo de otros sistemas como el de coagulación, el renal, cardiovascular o el endocrinológico entre otros. Esto permite entender que el tratamiento de la PE debe ser integral, con una visión holística del padecimiento y que el manejo médico de la paciente con PE requiere un equipo multidisciplinario, que actué armónicamente, para así alcanzar el éxito terapéutico con las menores secuelas posibles para el binomio madre-feto o madre-recién nacido.
La respuesta innata y la PE
El sistema del complemento está integrado por más de una treintena de proteínas séricas que de manera secuencial se activan una a otra. La descripción inicial de las funciones del complemento se centró en su capacidad para lisar microrganismos, y de esta manera funcionaba como el complemento al anticuerpo para lisar bacterias. Hoy conocemos que la actividad lítica es una más de las muchas y variadas funciones efectoras y reguladoras que el complemento cumple como integrante de la respuesta inmunológica. La función del complemento está altamente regulada en el embarazo y se han descrito factores solubles y receptores de membrana que limitan la actividad del complemento. Tales factores reguladores/inhibitorios de la activación del complemento son quienes hacen posible el estado de tolerancia en el embarazo. El componente fundador del sistema es el denominado C3, este componente hace posible la activación y amplificación del sistema. Por otro lado, los inhibidores de la activación del complemento pueden ser:
Solubles, que incluyen el inhibidor de C1 (C1INH), la proteína de unión a C4b (C4BP), el factor H, la clusterina y la vitronectina.
Reguladores en membrana, que incluyen CD35, CD46, CD55 y CD59.5
Se ha reportado que pacientes con PE expresan una menor actividad de varias de estas moléculas inhibidoras. En contraste con lo que sucede en el embarazo fisiológico, en la PE es la activación del complemento lo que se favorece, con una mayor activación de C3 y una menor inhibición/regulación tanto de factores solubles como de membrana. Por ejemplo, el binomio de factor H y factor I actúan juntos para inactivar a C3b y así inhibiendo la actividad de la C3 convertasa de la vía alterna se ha determinado que las células del trofoblasto expresan fisiológicamente el factor H, sin embargo, en algunos pacientes con PE el factor H materno no es funcional5; todas estas observaciones en pacientes con PE pueden explicar que una menor regulación del sistema de complemento puede llevar a desarrollar PE.

También se han detectado una elevada cantidad de productos de activación del complemento en pacientes con PE tanto EOPE como LOPE6, la causa de esta activación no es clara, aunque, se sabe que el desarrollo de inflamación en etapas tempranas de la placentación fisiológica es necesario7; además, se ha determinado que la remodelación de tejido placentario y la remoción de células apoptóticas por fagocitosis y opsonización en placenta puede activar “inespecíficamente” la activación del complemento, lo que aumentaría la respuesta inflamatoria. Asimismo, se sabe que C4BP se une a la cromatina y facilita la eliminación de DNA fetal libre, en contraste con pacientes con PE, se ha determinado un incremento de DNA fetal libre en sangre periférica, este DNA libre tiene un efecto proinflamatorio que apoya el establecimiento y perpetuación del estado de PE. Además, la sobreactivación del complemento en PE puede ser un estímulo para células endoteliales y otras células participantes de la inflamación que sintetizan la tirosina cinasa 1 de tipo FMS soluble (sFlt-1, por sus siglas en inglés), esta cinasa es una antagonista soluble del factor de crecimiento vascular-endotelial (VEGF, por sus siglas en inglés) y del factor de crecimiento de placenta (PlGF, por sus siglas en inglés), en pacientes con PE se han detectado una elevada concentración sérica de sFlt-1 derivada de placenta, lo que resultaría en la alteración del modelamiento vascular, de la angiogénesis y la restricción del crecimiento placentario5,8. El complemento puede ser un gran amplificador de la desregulación que se observa en PE, y no solo favoreciendo su activación sino limitando su inhibición. El complemento tiene puntos de activación compartidos con el sistema de coagulación, o de la cascada de bradicininas, por lo que se tiene una gran diversidad de puntos de regulación que pueden estar alterados en PE y que solo el estudio profundo podrá decirnos cual sería la mejor manera para tratarlos médicamente.
El desarrollo fisiológico de la placenta incluye el desarrollo de apoptosis y la liberación de vesículas extracelulares desde el sincitiotrofoblasto9. En condición fisiológica, las vesículas derivadas de la placenta incluyen a exosomas, micropartículas, microvesículas y cuerpos apoptóticos, todas estas vesículas en conjunto limitan la inflamación a través de citocinas, sin embargo, en PE el efecto de las vesículas extracelulares provoca daño endotelial, inflamación e hipercoagulación clásicamente observada en esta patología10.

Son necesarios más ensayos para entender con mayor profundidad la regulación de la respuesta inmunológica por efecto de las vesículas extracelulares de origen placentario en el embarazo fisiológico y fisiopatológico, sin embargo, podemos concluir que este mecanismo suma un nivel de complejidad al estudio de la respuesta inmunológica en el embarazo y que podría representar un eslabón que conecta los mecanismos que operan en la fase soluble con aquellos que lo hacen como respuestas celulares.
Las respuestas celulares innatas también ocurren en el contexto de la tolerancia inmunológica del embarazo. Por ejemplo, los neutrófilos provocan la diferenciación de células T a fenotipos reguladores como GARP+CD127lowFoxP3+,11 estas células T GARP+CD127lowFoxP3+ obtienen este fenotipo por acción de neutrófilos y secretan IL-10, IL-17 y VEGF in vitro. Utilizando un modelo con ratón se determinó que la eliminación de neutrófilos durante la preñez resulta en una limitada invasión trofoblástica y un pobre desarrollo de arterias espirales en la decidua, interesantemente, estos cambios en el tejido recuerdan a lo observado en PE. También, se determinó que neutrófilos de mujeres con PE inducen un bajo porcentaje de células Th1, por lo que la función limitada en los neutrófilos puede favorecer la desregulación del sistema y condiciona la placentación defectuosa en PE11. Además, se ha determinado que los neutrófilos pueden activarse por acción de lípidos oxidados secretados por la placenta dañada, e invaden la vasculatura materna, liberando especies reactivas de oxígeno (ROS, por sus siglas en inglés), TNF-α y mieloperoxidasa (MPO) que dañan el endotelio y facilitan el desarrollo de PE12. También, se ha observado que el número de neutrófilos en pacientes con PE aumenta, aunque paradójicamente su actividad fagocítica esta disminuida13. Finalmente, los neutrófilos en pacientes con PE tienden a expresar más fácilmente trampas extracelulares de neutrófilos (NETs, por sus siglas en inglés), que favorecen el estado inflamatorio, el daño endotelial y el desarrollo de eventos trombóticos, por lo que se favorece la fisiopatología observada en la PE. En conclusión, los neutrófilos de forma directa o indirecta facilitan el desarrollo de PE, sin embargo, se ha reportado que la vida media de los neutrófilos es corta (de horas) por lo que este factor debe ser evaluado en estudios posteriores y conocer si los cambios observados en los neutrófilos son causa o solo una asociación con la PE. Hacen falta más estudios para determinar el papel de neutrófilos en la PE.
Los macrófagos también participan en el establecimiento de la tolerancia en el embarazo, se ha determinado que entre el 20 y 30% de los leucocitos en la interfaz materno-fetal son macrófagos, y pueden expresar fenotipos tanto proinflamatorios (células M1) como antiinflamatorios (células M2)14. La población de macrófagos presente en la decidua expresa el fenotipo M2 en condición fisiológica y frecuentemente están localizados cerca de las arterias espirales, sin embargo, en PE es más frecuente encontrar a macrófagos con fenotipo M1 que producen TNF y óxido nítrico (NO)15. Dada la abundante presencia de macrófagos en decidua y su gran potencial como fuente de citocinas proinflamatorias, es probable que la génesis de algunos de los casos de PE sea inicialmente un estímulo infeccioso que active macrófagos, esta activación sería dependiente de patrones moleculares asociados a patógenos (PAMPs, por sus siglas en inglés), sin embargo, no siempre el componente infeccioso está presente, por lo cual se ha pensado que se puede desarrollar la denominada “inflamación estéril”, la cual es inducida por patrones moleculares asociados a daño/peligro (DAMPs, por sus siglas en inglés). Algunos de estos DAMPs, como: ácido úrico, ácido palmítico, vesículas extracelulares, DNA libre o ácidos grasos libres alcanzan concentraciones elevadas especialmente en PE16. Estos PAMPs o DAMPs provocan que macrófagos secreten citocinas proinflamatorias como IL-1β o IL-18. Debemos recordar que, para obtener la forma madura de IL-1 IL-1β, la caspasa-1 debe realizar un corte proteolítico sobre la pro-IL-1 IL-1β. La caspasa-1 a su vez es producto del inflamosoma NLRP3, este complejo molecular es clave como inductor de la inflamación estéril, ya que activa a la caspasa-1 dando lugar a inflamación exacerbada y piroptosis (un tipo de muerte celular por inflamación). Recientemente, se ha reportado que varios de los DAMPs producidos en altas concentraciones en PE activan al inflamosoma NLRP3 en macrófagos deciduales, esta serie de activación facilita la desregulación del proceso inflamatorio en PE. Además, se ha determinado que en condiciones fisiológicas los macrófagos deciduales fagocitan células apoptóticas y esto limita su función fagocítica, sin embargo, cuando estas células fagocitan células necróticas en PE, se activan y diferencian hacia fenotipos M1 proinflamatorios, lo que podría activar excesivamente al inflamosoma NLRP3. Hacen falta más estudios para entender mejor el proceso fisiopatológico y establecer una terapéutica médica para controlar la activación de macrófagos M1 en pacientes con PE.
Las células NK funcionan como un eslabón entre la respuesta celular innata y la adquirida, son linfocitos con gran potencial citotóxico que inicialmente se pensó eran perjudiciales para el mantenimiento de la gestación, hoy se reconoce que las células NK deciduales (dNKs) son indispensables como inductoras y modeladoras del proceso fisiológico de decidualización17. Las células NK son por mucho los linfocitos más abundantes en la interfaz materno-fetal, más del 50% de los linfocitos en decidua son NKs18. Las células dNK tienen menor actividad citotóxica y mayor capacidad para secretar citocinas como VEGF o PlGF que las NK en sangre periférica (pNKs)19, las dNK modelan la vasculatura arterial en placenta al secretar VEGF o PlGF y participan activamente en el control fisiológico de la gestación18,20, sin embargo, cuando esta capacidad secretora es menor o se expresa en tiempos incorrectos se obtiene un modelaje vascular patológico similar al observado en la PE.
La funcionalidad de las dNK es regulada por el factor de crecimiento transformante (TGF), esta citocina también diferencia a células T hacia un fenotipo regulador (células Treg), ambos tipos celulares aportan la regulación que favorece la tolerancia inmunológica en el embarazo. Se ha reportado un número de dNK aumentado en PE y se puede observar que la concentración de TGF esta elevada, además de una excesiva acumulación de Treg en decidua, por lo que de forma paradójica las células dNK limitan su actividad angiogénica, esta actividad de dNK afecta la vasculatura nuevamente favoreciendo el desarrollo de PE21. También, las células dNK pertenecen al grupo de células linfoides innatas (ILC, por sus siglas en inglés), este grupo de células participan en varios procesos de organogénesis y modelado tisular, por lo que su estudio puede aportar conocimiento decisivo para entender la fisiopatología de la PE y planear estrategias terapéuticas para esta enfermedad.
La figura 2 esquematiza la participación de los mecanismos inmunológicos efectores y reguladores innatos en el establecimiento de la tolerancia inmunológica del embarazo o el proceso fisiopatológico en la PE.
Elementos solubles y celulares de las respuestas innata actúan fisiológicamente a varios niveles; eliminan señales peligrosas (DNA fetal libre), educan células reguladoras (inTs), controlan la angiogénesis, entre otras funciones, sin embargo, cuando se desregulan (como es el caso de PE) una alteración puede llevar a la desregulación de la siguiente, como en un “efecto domino”.
La respuesta adaptativa y la PE
La respuesta inmunológica adaptativa se desarrolla al activar a células B y T22,23. Estas células reconocen con alta afinidad antígenos específicos. Los linfocitos B y T se originan en la médula ósea donde se diferencian a sus formas maduras, llevando un proceso complejo de expansión clonal y diferenciación celular muy controlada para eliminar las clonas autoreactivas. Es importante recordar que los linfocitos T adquieren el compromiso a diferenciarse como células T desde su estadía en la médula ósea, aunque el proceso completo de diferenciación se cumple en el timo. Tanto las células B como las T, alcanzan su estado maduro al expresar en su membrana celular receptores específicos contra el antígeno. El BCR en el caso de células B, y el TCR en el caso de las células T. De forma general podemos mencionar que los linfocitos B pueden ser de tipo B1 (muchas de ellas expresan el marcador CD5) o tipo B2 (que son las células B convencionales que encontramos en sangre periférica). Ambos tipos celulares B desarrollan respuestas humorales al diferenciarse a células plasmáticas productoras de anticuerpos, sin embargo, podemos mencionar que las células B1 montan respuestas contra antígenos independientes de la ayuda por células T, y las células plasmáticas que se generan entonces son de vida corta (un par de semanas a meses), y que liberan IgM desde zonas extrafoliculares desde el ganglio linfático o el bazo. De forma complementaria a esta respuesta, las células B2 dan origen a células plasmáticas de vida larga (algunas viven por el resto de la vida del animal o persona que las desarrolla), y para obtener este fenotipo de células plasmáticas se requiere que las células B2 obtengan ayuda de linfocitos T, esta ayuda proviene generalmente de linfocitos T foliculares (células Tfh), por lo que la diferenciación de células plasmáticas que se desarrolla por esta vía de cooperación B-T incluye el cambio de isotipo (de IgM a IgG, IgA o IgE). Este “switch de isotipo” depende de las citocinas en el microambiente en el folículo linfoide del ganglio o del bazo. También, recientemente se ha estudiado el fenotipo y funcionalidad de las denominadas células B reguladoras (linfocitos Breg), estas células son capaces de presentar antígeno y limitar la activación de un gran número de células24, es interesante que las Breg parecen cumplir su función en presencia de inflamación, esto podría ser de gran interés en el caso de la PE donde hay un proceso inflamatorio crónico e intenso.

Recientemente la función de las células B en PE ha sido extensamente estudiada y se han planteado nuevos acercamientos terapéuticos para tratar esta enfermedad25. Se determinó que células aisladas de membranas amnióticas, coriónicas y vellosidades coriónicas, entre otras, pueden apoyar la diferenciación de células B hacia un fenotipo regulador, estas células aisladas de tejido placentario y cordón umbilical son llamadas “células perinatales”, y son mayoritariamente células del estroma mesenquimal. Estas células perinatales ejercen una función inhibitoria de la activación y proliferación de células B26, este efecto no necesita contacto célula a célula, por lo que señales solubles (como citocinas u otros factores) son las encargadas de dirigir la diferenciación de células B27. También, las células perinatales modulan la diferenciación de células plasmáticas. Es probable que las células perinatales a su vez requieran de un estímulo que permita dirigir la diferenciación de células B a Breg, y un estímulo que puede activar a las células perinatales es la presencia de oligodesoxinucleótidos CpG (CpG ODN, por sus siglas en inglés), aunque se desconoce a detalle los mecanismos por los cuales las células perinatales regulan a células B, sin embargo, se ha observado que las células Breg derivadas de células perinatales expresan IL-10 y de esta manera ejercen su actividad inhibitoria sobre respuestas proinflamatorias. Es por estos antecedentes que el estudio de células perinatales es de mucho interés para propósitos terapéuticos en PE.

Como se menciona antes, son dos las principales poblaciones de células B, las B1 tienen su origen en el hígado fetal, y se pueden identificar por la expresión de CD5, CD27 y CD43, mientras que las células B2 tienen su origen postnatal en la médula ósea. Durante el embarazo fisiológico las células B maduran a células plasmáticas productoras de anticuerpos asimétricos, estos anticuerpos son univalentes28, es decir, uno de sus sitios de reconocimiento antigénico está ocupado por carbohidratos y no es funcional29, estos anticuerpos univalentes reconocen antígenos paternos pero no activan complemento, fagocitosis o citotoxicidad25, por lo que es un mecanismo más que favorece el estado de tolerancia inmunológica; se ha reportado que en pacientes con PE la presencia de anticuerpos asimétricos disminuye, por lo que habría mayor activación de complemento y potencialmente mayor daño vascular. Además, se ha observado que durante la PE las células B frecuentemente no expresan el fenotipo regulador, pero, en contraste, las células B en PE maduran a células plasmáticas que secretan anticuerpos contra el receptor de tipo 1 de la angiotensina (AT1R, por sus siglas en inglés), estos anticuerpos inyectados en ratones reproducen las características clínicas de la PE de humanos como son: hipertensión, proteinuria, anormalidades placentarias y restricción de crecimiento fetal30. La presencia de otros autoanticuerpos (antiproteína C, antiproteína S o antitiroides) apunta a que en la PE se desarrolla una respuesta autoinmune. El origen de los autoanticuerpos podrían ser las células B CD19+CD5+, ya que el número de estas células normalmente disminuye en sangre periférica de mujeres con embarazo fisiológico, pero permanece elevado en mujeres con PE31. Además, se ha reportado que células CD19+CD5+ producen autoanticuerpos contra AT1R cuando son cultivados con suero de pacientes con PE. Estos hallazgos en el campo de estudio de células B permiten evaluar de mejor manera a los modelos animales que expresan una patología similar a la PE en humanos. LaMarca et al., reportaron que el uso de rituximab puede eliminar células B y reducir la presión sanguínea, así como la concentración de anticuerpos anti-AT1R en ratones32,33, por lo que es posible que en un futuro cercano se utilice rituximab con intención terapéutica en pacientes con PE, aunque la seguridad del uso de este medicamento en el embarazo debe ser minuciosamente analizado y monitorizado.

Mientras las células B desarrollan repuestas humorales, las células T cooperadoras (linfocitos T CD4+ cooperadores, Th) apoyan la diferenciación de células B o ejercen citotoxicidad (linfocitos T CD8+ citotóxicos). Los linfocitos T son muy diversos fenotípica y funcionalmente. Al clasificar a las células T por la composición del TCR observamos dos poblaciones: los linfocitos Tab, que son más del 90% de las células en sangre periférica, y los linfocitos Tgd, que representan solo del 2 a 3% de las células circulantes en sangre periférica. Las células Tab a su vez podemos clasificarlas como CD4+ o CD8+. Las células T se originan de un progenitor común linfoide en médula ósea, que luego migra a timo donde pueden completar la diferenciación a células T maduras. Los linfocitos T CD4+ dan origen a fenotipos Th1 productoras de interferón-gamma (IFN-γ, por sus siglas en inglés) entre otras citocinas, o fenotipo Th2 productoras de Interleucina-4 (IL-4) entre otras citocinas. Algunas células T CD4+ también expresan CD25 y el factor de trascripción FoxP3 y ejercen funciones reguladoras (células Treg), estas Treg son importantes para el mantenimiento de la preñez en modelos animales y el embarazo fisiológico en humanos. Por ejemplo, pacientes con infertilidad y abortos repetidos expresan bajo porcentaje de células Treg. En contraste, las células T productoras de IL-17 son denominadas células Th17 que favorecen el desarrollo de la inflamación y respuestas Th1 potencialmente peligrosas para el desarrollo de la gestación.

El trofoblasto placentario no activa a células T, pues el trofoblasto no expresa moléculas clase I-A o B del complejo principal de histocompatibilidad (MHC, por sus siglas en inglés), en contraste, las células del trofoblasto expresan moléculas clase I-C del MHC, la expresión de esta molécula permite que las células T puedan ser tolerantes contra antígenos paternos en la interfaz materno-fetal apoyando así el desarrollo del embarazo. Además, células T CD8+ activadas son eliminadas en el trofoblasto vía la función de PD-1 y PDL-1, lo que disminuye el riesgo de efecto citotóxico contra antígenos paternos. También, la expresión de moléculas clase II del MHC (como HLA-DR) es limitada o ausente en el trofoblasto reforzando su carácter regulador negativo de la activación de células T maternas potencialmente peligrosas para el tejido fetal. En contraste, en el estado de PE se ha determinado que en la interfaz materno-fetal hay la expresión aberrante de HLA-DR, lo que posibilita la activación de células T y el consecuente daño al tejido placentario incluido el endotelio o el vascular34. Además, se ha determinado que NKs participan en el cambio de fenotipos Th1 en el primer trimestre del embarazo a Th2 del segundo trimestre35, indicando que la regulación de las celulares dependen de diversos fenotipos que trabajan coordinadamente para apoyar la gestación fisiológica. Hasta el momento se desconoce si este tipo de interacción NK-Th1/Th2 está alterada en pacientes con PE.

Otro tipo de células que es importante de analizar en pacientes con PE son las células Th17. Se ha reportado un aumento en la proporción de Th17 circulantes en mujeres con PE36, lo que apoyaría el proceso inflamatorio. Se ha determinado que, en varias enfermedades autoinmunes, las células Th17 pueden reclutar gran cantidad de neutrófilos, y que estas células son grandes generadoras de estrés oxidativo que se observa en PE. Células Th17 que promueven inflamación al liberar IL-17 y linfocitos Th1 también promueven la inflamación al producir TNF-α e IL-6, la actividad de células Th17 y Th1 puede generan daño endotelial que favorece la progresión de la PE37. También, el estímulo con IL-17 es importante factor para la expresión de autoanticuerpos contra el AT1R38, y el aumento de células proinflamatorias puede favorecer la disminución de células reguladoras en PE; de hecho, se ha reportado una disminución en la proporción de células Treg productoras de IL-10. El amplio espectro de regulación que ofrece IL-10 hace que esta molécula sea un candidato para usarlo con intención terapéutica en pacientes con PE, aunque hacen falta muchos ensayos con modelos animales para determinar la seguridad de esta maniobra con intención terapéutica. La figura 3 esquematiza la participación de los mecanismos inmunológicos efectores y reguladores celulares en el establecimiento de la tolerancia inmunológica del embarazo o el proceso fisiopatológico en la PE.

Células de las respuestas innata y adaptativa participan redundantemente en la tolerancia inmunológica, el incremento o disminución de alguna de estas poblacionescelulares afecta la función y número de algunas otras y así se establece la fisiopatología en PE. Recientemente el control de células B ha llamado la atención como“blanco terapéutico”, pues se piensa que la PE es una enfermedad autoinmune que debe ser tratada al menos parcialmente con ese enfoque. Imagen vectorial: Freepik
Figura 3 Componentes de respuesta inmunológica adquirida que participan en la “tolerancia inmunológica del embarazo” y la fisiopatología en preeclampsia

CONCLUSIÓN
En el embarazo se establece uno de los escenarios más complejos que enfrenta el sistema inmunológico, en ese estado se desarrollan múltiples mecanismos que conforman la tolerancia inmunológica de este estado, tales mecanismos actúan de forma armónica durante tiempos específicos en la ontogenia, esta red compleja que soporta la tolerancia inmunológica puede ser afectada por infecciones o señales de peligro que desregulan el sistema.
Tanto los componentes humorales como celulares de las respuestas innata o adquirida pueden ser afectados por la estimulación con PAMPs o DAMPs. Se ha reportado tanto el aumento como la disminución de fenotipos celulares asociados con el estado de PE, por ejemplo, una disminución de células Breg o un aumento en la frecuencia de células Th17. Además, la disminución de factores solubles, como el factor H para el complemento o el aumento de citocinas proinflamatorias puede llevar a la desregulación del sistema y el establecimiento del estado de preeclampsia.
Tanto factores solubles como celulares que participan en la respuesta inmunológica pueden dañar el endotelio o la vasculatura en placenta lo que favorece el desarrollo de la PE.
Finalmente, recientemente se ha postulado que dada la presencia de autoanticuerpos que provocan la expresión de muchas de las características clínicas de la PE, esta enfermedad podría ser considerada como una patología por autoinmunidad, la pérdida de la tolerancia inmunológica en PE podría ser una de las piedras angulares en la fisiopatología que lleve a proponer nuevos acercamientos terapéuticos exitosos para el control de esta enfermedad.

