











 nova página do texto(beta)
nova página do texto(beta) Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO 
 Permalink
PermalinkINTRODUCTION
Paleontology (particularly vertebrate paleontology) benefitted immensely from the advent of isotopic dating at accessible price that occurred in the early sixties. It allowed calibration of the Geochronologic Time-table (Time-scale), which up to then was based on bioevents, such as the first/last occurrences of significant selected taxa (e.g. fusulinids, ammonites, planctonic foraminifers and terrestrial tetrapods, largely mammals). This, in turn, made possible a more precise long range correlation of geologic events s.l. across distant regions/continents, thus producing a qualitative leap in the understanding of the history and evolution of the Earth System, and of the living beings that populate it.
The application of isotopic dating techniques started early in México, with the foundation in 1962 of the Laboratorio de Geocronometría, Instituto de Geología, Universidad Nacional Autónoma de México, which mainly focused on dating Precambrian, Paleozoic and Mesozoic events. The Mixteca region, northwestern Oaxaca state in the Sierra Madre del Sur Morphotectonic Province (MP hereafter) was one of the first, where isotopic dating of Cenozoic events was attempted during the development of the senior author's doctoral dissertation in the University of Texas at Austin (Ferrusquía-Villafranca, 1971), through the cooperation of Professors Leon Long and Fred McDowell.
The systematic effort to isotopically date actually or potentially vertebrate-bearing Cenozoic continental sequences was carried out by means of a far-reaching multidisciplinary and multi-institutional research project. This study produced K-Ar ages in selected areas from the Mixteca region, Oaxaca (~30 Ma), Valle de Oaxaca (~20 Ma in the northern part and ~15 Ma in the southeastern part), Oaxacan Subisthmian region (~15 Ma), Oaxacan Isthmian region (~15 Ma), and northwestern Chiapas (~15 Ma). Such data allowed to calibrate three local faunas from Oaxaca (Suchilquitongo, 20 Ma; Matatlán, ~15 Ma) and one from Chiapas (Ixtapa, 15 ~Ma). The geologic and paleobio-logical significance of such discoveries will be discussed elsewhere.
In the last decades, similar efforts to date other vertebrate-bearing sequences have been carried out yielding important results, such as an Early Jurassic sequence in Tamaulipas, Sierra Madre Oriental MP (Fastovsky et al., 1998, 2005), a Late Cretaceous sequence in Baja California, Península de Baja California MP (Renne et al., 1991), an Early Paleogene sequence and fauna in Guanajuato, Central Plateau MP (Aranda-Gómez and McDowell, 1998), Late Neogene sequences from selected areas of the Trans-Mexican Volcanic Belt MP and neighboring regions of the Central Plateau MP (Carranza-Castañeda, 2006 and references therein) and lately in the Sierra Madre Oriental MP. The geologic and paleobiologic significance will be discussed in the section dealing with each fauna.
It should be noted that the geologic part of all these results varies enormously due to: (1) differences in the formality/depth given to the treatment of each sequence. (2) Intrinsic differences of each sequence, so that some bear vertebrates or dated rock material at more than one stratigraphic level. (3) More than one research team may have studied a given sequence. These facts may lead to confusion or ambiguity. To avoid these discrepancies, we deem it appropriate to describe each sequence in an uniform way and to evaluate the nomenclatorial status of the units with vertebrates and/or dated rock material. Both items influence the dependability of analyzed results.
The possibility to test the North American Land Mammal Age (NALMA) System's aptness to date faunas from 28 sites scattered throughout Mexico, is an added bonus. Such possibility is at present taken for granted, notwithstanding these facts: (1) it is based on the occurrence of bioevents (certain mammal taxa) in temperate (30°-50°N Latitud) North America (i.e., the contiguous United States; cf. Woodburne, 2004). (2) The occurrence of some taxa in Central Mexico and Alaska that does not conform to it (cf. Bell et al., 2004). Given the above, we aim at fulfilling the following objectives:
To critically present and review the geologic and vertebrate paleontologic information on the sites where this dual isotopic/ paleontologic approach has been applied during the last fifty years in this country.
To test by this review if the North American Land Mammal Age System is suitable to date vertebrate faunas in Mexico.
To contribute to a better understanding of this country's vertebrate faunistic evolution, and to a lesser extent, its geologic evolution as well.
METHODOLOGY
In Figure 1 and Table 1 we present the results of an exhaustive bibliographic search for isotopically dated lithostratigraphic units that bear studied vertebrate fossil assemblages, or that are stratigraphically related to such units, thus furnishing their minimal/maximal ages. Across the country, we detected 28 pre-Pleistocene sites that fulfill this dual criterion, from which reliable analyses can be made.
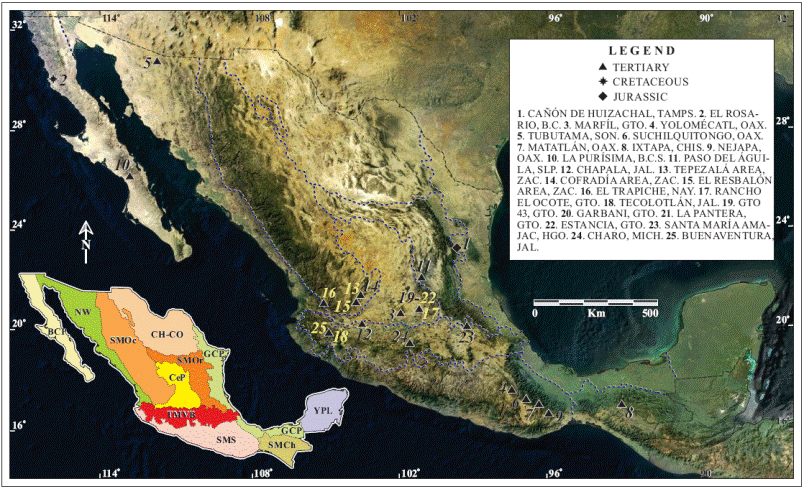
Figure1. Morphotectonic Provinces Map of Mexico showing the location of sites with isotopically dated lithostratigraphic units that bear vertebrates, plotted on a satellite image. [Source of image: CONABIO (2003)].
Table 1 Isotopically dated Jurassic, Cretaceous and Tertiary lithostratigraphic units from Mexico that bear studied vertebrates.
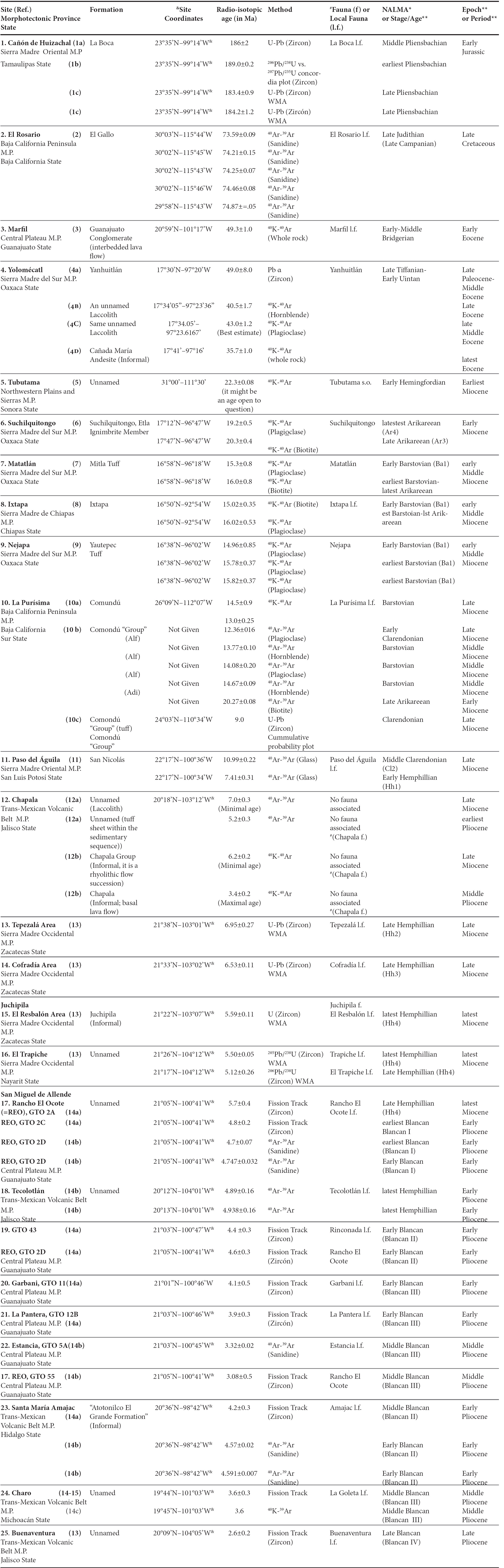
ABBREVIATIONS: Alf: Andesitic lava flow. Adi: andesitic dike. BCH: biochronologic reference [indicated by number in brackets beside the site name]. RCH, radio-isotopic reference [indicated by number in brackets beside the locality name]. NALMA: North American Land Mammal Age. WMA: weighted mean age.
SYMBOLS: *Geochronometric NALMA boundaries after Woodburne (2004), particularly Cifelli et al. (2004, Judithian, Late Cretaceous), Robinson et al. (2004, Wasatchian-Duchesnian, Eocene), Tedford et al. (2004, Arikareean-Clarendonian, Early-Middle Miocene), Bell et al. (2004, Blancan-Rancholabrean, Late Pliocene- Pleistocene). **Geochronometric Period, Stage/Age, and Epoch boundaries after Gradstein et al. (2004). & Latitude-Longitud of RCH sampling site not stated by author; coordinates inferred from information presented in the corresponding article. #Attributed fauna, i.e., a fauna or faunal assemblage known to occur in the area/locality, but not present in the dated/sampled section .
REFERENCES: [RCH1]: 1a, Fastovsky et al. (1998); 1b, Fastovsky et al. (2005); 1c, Rubio-Cisneros et al. (2011). [BCH1]: Clark and Hopson (1985), Clark et al. (1994), Fastovsky et al. 1995). [RCH2]: Renne et al. (1991); [BCH2]: Molnar (1974), Morris (1972, 1973a-b, 1981); Lillegraben (1972, 1976). [RCH3]: Aranda-Gómez and McDowell (1998). [BCH3]: Fries et al. (1955), Ferrusquía-Villafranca (1989, 2004, 2005). [RCH4]: 4a. Ferrusquía-Villafranca (1976). 4b. Martiny et al. (2000). 4c. Cerca et al. (2007). 4d. Santamaría-Díaz et al. (2008). [BCH4]: Jiménez-Hidalgo et al. (2011). [RCH5]: Arriaga-Meléndez et al. (1986). [BCH5]: Ferrusquía-Villafranca (1990a). [RCH6]: Ferrusquía-Villafranca and McDowell (1991). [BCH6]: Ferrusquía-Villafranca (1990a, 2003). [RCH7]: Ferrusquía-Villafranca and McDowell (1991), [BCH7]: Ferrusquía-Villafranca (1990a, 2003). [RCH8]: Ferrusquía-Villafranca (1996). [BCH8]: Ferrusquía-Villafranca (1990b, 2003). [RCH9]: Ferrusquía-Villafranca (2001). [BCH9]: Ferrusquía-Villafranca (1990b, 2003). [RCH10]: 10a. Mc Lean et al. (1987). 10b. Umhoefer et al. (2001). 10c. Fletcher et al. (2007). [BCH10]: Ferrusquía-Villafranca (1990a). [RCH11]: Ferrusquía-Villafranca et al. (2015). [BCH11]: Ferrusquía-Villafranca et al. (2014). [RCH12a]: Israde-Alcántara et al. (2010).[BCH12a]: Israde-Alcántara et al. (2010 and refs. therein), Ferrusquía-Villafranca et al.2010). [RCH12 b]: Rosas-Elguera and Urrutia- Fucugauchi (1998), cited by Lucas (2008). [BCH12b]: Lucas 2008 and refs. therein), Ferrusquía-Villafranca et al. (2010). [RCH13]: Carranza-Castañeda et al. (2013). [BCH13]: Carranza-Castañeda et al. (2013). [RCH14]: 14a, Kowallis et al. (1998); 14b, Flynn et al. (2005). 14c. Israde-Alcántara and Garduño-Monroy (1999). [BCH14]: 14a, Carranza-Castañeda (2006 and refs. therein). [BCH15: Repenning (1962), Miller and Carranza (1984), Carranza-Castañeda (2006 and refs. therein).
The sites are plotted on a morphotectonic provinces (MP) map of México (cf. Ferrusquía-Villafranca et al., 2010), because the geological significance of the sites-information could be perceived better there, than in a political division-state map. The sites are listed in Table 1, which includes eight information items (columns); they are explained as follows:
Geographic-Geologic: The first column indicates the morpho-tectonic province and the state to which the site belongs, each received a consecutive number; a site may include a single locality or a cluster of spatially close localities, which are designated area sites. The chief references (both isotopical and paleontological) are given too (bracketed number after the site-name) and spelled out as foot notes. The second column shows the fossil-bearing unit name from which isotopic ages were obtained; the unit's condition (named or unnamed) is stated; the named units are classified as formal or informal, depending on whether or not their proposal fulfills the requirements of the North American Stratigraphic Code (NACSN, 2005) or of the pertinent code at the time of their proposal. The third column gives the site coordinates as stated in the corresponding paper or as inferred from information given there; this last mode is expressed by the sign & as an exponent.
Isotopic Geochronologic: The fourth column bears the numerical ages. The fifth column states the dating method (K-Ar e.g. Bonhomme et al., 1975; 40Ar-39Ar, e.g. van der Pluijm et al., 2006; U-Pb, e.g. Gehrels et al., 2008; fission track, e.g. Wagner and Van den Haute, 1992) and the mineral used; in some instances, the latter was not stated in the corresponding paper.
Paleontologic-Geochronologic: The sixth column lists the names of the fossil assemblages studied, they may be a fauna (f., an assemblage collected from several spatially close localities that lie in a given area or region), local fauna (l.f., an assemblage collected from a single locality, or spatially very close localities) and single occurrence [s.o., vertebrate remain(s) that belong to a single individual/taxon]. Such assemblages are usually named after a geographic s.l. feature. The seventh column shows the biochronological age of each assemblage, which apart from the Jurassic, is stated in terms of the North American Land Mammal Ages (acronymed NALMA) as presented in Woodburne (2004), the standard reference for mammalian biochronology. The eighth column carries the geochronologic information expressed in epochs and periods sensu Gradstein et al. (2004).
Footnotes at the end of Table 1: they explain symbols and provide bibliographic references to the isotope geochronology data (RCH), and the biochronological (BCH) data; both are keyed to the bracketed number on the right of the first column. Notice that in some instances, two or more sites may carry the same bibliographic reference number; this indicates that in the corresponding paper, more than one site was studied. On the other hand, the same site could have been independently studied by two or more different research teams, accordingly we have cited all papers involved, giving them independent bibliographic reference numbers; in the text we discuss and parsimoniously assess them.
Terms, abbreviations and considerations
The boundaries of epochs/stages are taken from Gradstein et al. (2004). The sedimentary petrographic nomenclature follows Folk (1974) and Boggs (1995), the pyroclastic terminology used is from Fisher and Schmincke (1984), the sedimentary facies nomenclature follows Miall (2006, 2010) for the fluvial facies and Collinson (1996) for the lacustrine facies. In some instances, old descriptions were rediscribed/interpreted in current terminology and/or conceptions. The Morphotectonic Province boundaries were taken from Ferrusquía-Villafranca et al. (2010).
Morphotectonic province abbreviations
BCP, Baja California Peninsula; NW, Northwestern plains and sierras; SMOc, Sierra Madre Occidental; CH-CO, Chihuahua-Coahuila plateaus and ranges; SMOr, Sierra Madre Oriental; GCP, Gulf Coastal Plain; CeP, Central Plateau; TMVB, Trans-Mexican Volcanic Belt; SMS, Sierra Madre del Sur; SMCh, Sierra Madre de Chiapas; and YPL, Yucatán Platform.
Other Considerations
(a) the Pleistocene sites were not included. (b) The lithostratigraphic vertebrate-bearing units that have been numerically dated by magneto- stratigraphic methods, have not been included either because the paper is solely focused on units dated by isotopic methods. (c) Appendix, Part A includes the geologic maps of the sites, Figures A1 through A19. Appendix, Part B includes the taxonomic composition of the faunas and faunal assemblages, Tables B1 through B12. (d) The total number of sites investigated is 28 (they are reported as such in the text and listed in Table 1). However, these facts may lead to different total numbers: (d.1) two or more sites may lie so close to each other, that they could not be discriminated, that is why in Figure 1, only 25 sites are plotted. (d.2) Several sites carry two sets of dated materials and vertebrate faunas, that is why in Table 1, only 23 sites are numbered. (d.3). Finally, only 27 sites are amenable to the North American Land Mammal Age system of dating (Woodburne, 2004); the Jurassic and older faunas could not be independently dated/corroborated by current magnetostratigraphic techniques.
RESULTS AND DISCUSSION
Table 1 is a summary of the geographic, geologic and geochronologic information on the detected sites/localities where both isotopically dated lithostratigraphic units and studied vertebrate fossils co-occur; they are treated below following a normal geochronologic order. For each site, the information on the geological setting is presented and discussed first; its isotopic dating and vertebrate fossil assemblage is offered and discussed next, finishing with the regional significance of both the geologic and paleontologic data obtained.
Jurassic
Cañón de Huizachal site
Tamaulipas State, Sierra Madre Oriental MP, Figure 1, Number 1; Table 1, Number 1; Figures A1 and A2
The geology of this place is known for long time (Carrillo-Bravo, 1961 and references therein), however, detailed information on the Valle del Huizachal is scarce (Mixon, 1963a, 1936b; Fastovsky et al., 2005; Rubio-Cisneros and Lawton, 2011). The latter two contributions will be briefly discussed due to their significant differences regarding the conceptions and cartographic boundaries of the units involved.
Both agree that the red bed sequence there belongs to the Huizachal Group, which includes La Boca Formation (lower part) and La Joya Formation (upper part). The first overlies a steeply bedded/banded rhyolitic succession sparsely intercalated by basaltic flows/breccias and terrigenous clastic strata (shales to conglomerates). Fastovsky et al. (2005) designated this succession Volcanic and Epiclastic Suite (VES), an informal unit (cf. NACSN, 1983, 2005) and interpreted it as recording a major angular unconformity due to tectonic activity prior to Huizachal deposition. Alternatively, Rubio-Cisneros and Lawton (2011) interpreted it as a rhyolitic dome and adjacent deformation zone developed in the host rock unit, an interpretation that we follow in this work.
La Boca Fm. is a ~54 m thick pyroclastic/epiclastic-volcarenitic succession consisting of two texturally/compositionally different parts, the lower one being largely pyroclastic (red, moderately altered, thinly bedded, felsic, crystal-vitric, ash-fall and ash-flow tuffs). Both fossils and zircon crystals for dating were taken from this part. The upper part is mainly epiclastic (fine to coarse-grained fluvial facies) and un-fossiliferous; Rubio-Cisneros and Lawton (2011) dated zircon crystals from this part (168-163 Ma). La Joya Fm. is a fluvial succession with a basal conglomerate (Rubio-Cisneros and Lawton, 2011).
The unit underlying the Huizachal Group does not crop out in the area, but it is known that in northern Mexico, red bed sequences like the Huizachal Group unconformably overlie crystalline basement rock units of diverse ages (Proterozoic to Late Paleozoic) and attributes (cf. Barboza-Gudiño et al., 2008 and references therein). The Huizachal Group is overlain by the Late Jurassic Zuloaga Fm., a predominantly carbonate sequence that marks the onset of marine deposition in northeastern Mexico (Salvador, 1987).
Dating and regional significance. The ages reported by Fastovsky et al. (1998, 2005) and Rubio-Cisneros and Lawton (2011) are moderately different (see Table 1). Fastovsky et al. (2005) preferred 189±0.2 Ma, earliest Pliensbachian [not Middle Toarcian, as they reported (cf. Gradstein et al., 2004)], as the maximal age of La Boca's lower part. On the other hand, Rubio-Cisneros and Lawton (2011) convincingly argued for slightly younger ages (184.2±1.2 Ma and 183±0.9 Ma, Late Pliensbachian).
At any rate, the Huizachal Group registers an Early to Middle Jurassic volcanism (from the Nazas Volcanic Arc; Barboza-Gudiño et al., 2008) coeval to fluvial deposition occurring in an extensional setting related to the dismembering of Pangea, and prior to the onset of the wide Late Jurassic marine transgression that extensively covered southern North America (including parts of Mexico; cf. Salvador, 1987).
La Boca local fauna is the earliest terrestrial vertebrate assemblage from Mexico, it includes mammal-like reptiles, early dinosaurs, pterosaurs, early diapsids, sphenodonts and early mammals (Clark et al., 1994; Clark and Hopson, 1985; Fastovsky et al., 1995, 1998; Reynoso, 1996), and closely resembles Early-Middle Jurassic faunas from North America (Kayenta Fm.), Eastern Asia (Lufeng Fm.) and South Africa (Clarence Fm.). The U-Pb dating results constrain the age to the late Middle Jurassic, thus refining the biochronologic assessment. The widespread geographic distribution of the taxa/faunas involved indicates that the Breaking up of Pangea-Vicarious Event separated taxa/ faunas once sharing the same area of distribution.
Cretaceous
El Rosario site
Baja California State, Baja California Peninsula MP, Figure 1, Number 2; Table 1, Number 2; Figure A3.
This site is well known long ago (cf. Kilmer, 1963; Gastil et al., 1975; Busby, 2004) however, published detailed geologic information (maps included) is scarce (cf. Renne et al., 1991; Fulford and Busby-Spera, 1993). The Cretaceous sedimentary cover in Baja California unconformably rests on the Aptian-Albian Alisitos Fm. or on the Mesozoic Batholith; this cover includes three units, readily observed in the area: Bocana Roja Fm. (lower, continental, coarse-grained, unfossiliferous), El Gallo Fm. (middle, continental to transitional, fine-clastic, vertebrate-fossiliferous) and El Rosario Fm. (upper, marine, carbonatic and fine-siliciclastic). Late Cenozoic-Quaternary alluvial deposits unconformably rest on this cover.
In turn, El Gallo Formation consists of three parts: La Escarpa member (lower, ~150 m thick, fluvial facies), El Disecado member (middle, ~1,150 m thick, fluvial and subtidal facies, vertebrate-fossiliferous, intercalated at various levels by rhyolitic, crystal-vitric tuff sheets) and El Castillo member (upper, continental, meso to coarse-grained, it does not crop out in the area; Renne et al., 1991).
Dating and regional significance. The 40Ar-39Ar ages obtained from the tuff sheets have a small spread (74.87±0.05 Ma to 73.59±0.09 Ma, Renne et al., 1991; Table 1), placing this unit and its fauna in the Late Campanian; further, these authors also dated explosive volcanic activity occurring concurrently with fluvial (and subsidiary tidal) deposition within the rapidly subsiding Late Cretaceous El Rosario Basin, a small portion of the Peninsular Ranges Forearc Basin (Renne et al., 1991). On the other hand, Busby (2004) in her regional tectonic study, convincingly argues for regarding El Rosario as a forearc basin itself, and so names it (Busby, 2004, fig. 3).
El Rosario l.f. (Lillegraven, 1972, 1976; Morris, 1972, 1973a, 1973b, 1981; Molnar, 1974) includes hadrosaurian and theropod dinosaurs as well as mammals (non-therian and therian), it resembles Judithian faunas of western North America (e.g. that from the Judith River Fm.; Cifelli et al., 2004) and thus it was assigned to the Judithian NALMA (Cifelli et al., 2004); the isotopic dating corroborated/refined this. El Rosario l.f. is the southwestern most Cretaceous assemblage of North America.
Eocene
Marfil site
Guanajuato State, Central Plateau MP, Figure 1, Number 3; Table 1, Number 3; Figure A4.
This site is located in the Guanajuato Mining District, which holds the world class silver deposits of Guanajuato and, is geologically known long ago (cf. Edwards, 1955; Echegoyen-Sánchez et al., 1970, whose detailed lithostratigraphy is followed here).
The Esperanza Formation (a Late Mesozoic metamorphic complex with Late Cretaceous-Early Tertiary granitoid intrusions) constitutes the oldest unit and is unconformably overlain by the Early Eocene Guanajuato Conglomerate, a ~2,000 m thick fluvial sequence formed by two members. The lower is ~1,300 m thick, largely consisting of fine grained, arkosic-volcarenitic sediments (overbank facies); it is little fossiliferous, and is also sparsely intercalated by andesitic lava flows. The upper member unconformably overlies the lower, it is a ~700 m thick coarse-grained fluvial sequence (alluvial fan and debris flow facies) expressed as deep red, clast to matrix supported conglomerate strata; it records rapid source-area uplift.
The Guanajuato Conglomerate is non-conformably overlain by a thick volcanic succession that includes upward these units: Losero Fm. (rhyolitic ash-flow tuffs, and tuffaceous sandstones), Bufa Rhyolite (a ~30 m thick felsic ignimbrite sheet that yielded K-Ar ages of 37.5±3 Ma; Gross, 1975), Calderones Fm. (?Oligocene, a ~600 m thick, largely pyroclastic pile), Cedro Fm. (?Pliocene, a ~50 m thick, largely basaltic lava flow stack), Capulín Conglomerate (?Pliocene, a ~120 m thick, poorly consolidaded fluvial sequence) and Quaternary deposits (alluvium, colluvium and soils) unconformably overlie the older units.
Dating and regional significance. A lower member lava flow yielded a 40K-40Ar age of 49.3±1.0 Ma (Aranda-Gómez and McDowell, 1998; Table 1), placing the Guanajuato Conglomerate in the Early Eocene, which is confirmed by the Bufa Rhyolite age. Previously though, Fries et al. (1955) reported Late Eocene-Early Oligocene vertebrates from this unit (now known to occur ~400 m above the dated flow), which afford the first objective dating of the taphrogenic event immediately succeeding the Laramide Orogeny in Mexico, as well as the associated magmatism (both volcanic and intrusive). The geologic importance of this fact needs no further stressing.
The Marfil l.f. (Appendix Part B, Table B1; Fries et al., 1955; Black and Stephens III, 1973; Ferrusquía-Villafranca, 1989, 2004, 2005) includes reptiles (an iguanid), birds (a falconiform) and mammals [five orders (Condylarthra, Carnivora, Rodentia, Perissodactyla and Uncertain-Paleanodonta), six families, eight genera]. The condy- larth Hyopsodus and perissidactyl Helaletes are characteristic Early Bridgerian taxa (cf. Robinson et al., 2004); largely on this basis this fauna is assigned to the Bridgerian NALMA (Robinson et al., 2004). The lava flow age further corroborates this assignment. Finally, the fauna shows an overall North American biogeographic affinity, but endemism is apparent; some of the rodents (e.g. Marfilomys) show vague affinities with the early caviomorphs of South America (cf. Patterson and Wood, 1982).
Yoloméctal site
Oaxaca State, Sierra Madre del Sur MP, Figure 1, Number 4; Table 1, Number 4; Figure A5.
Detailed geologic information for this place is lacking. The summary presented below draws from Ferrusquía-Villafranca (1976, adjacent northern area), López-Ticha (1985) and Santamaría-Díaz et al. (2008), the latter two are regional works that include this site. The oldest unit is the Middle Jurassic Tecocoyunca Group, unconformably overlain by the folded/faulted/uplifted Late Cretaceous Teposcolula Limestone and Yucunama Marl, which form a north-south trending range. The Cenozoic sequence includes these units (from the bottom up): ?Yanhuitlán Fm. (Eocene, fluvio-lacustrine, fine-grained facies prevail; it is sparsely fossiliferous), Llano de Lobos Tuff (Oligocene, a thick pyroclastic sheet), Yucudaac Andesite (Oligocene, a thick flow stack), San Marcos Andesite (Oligocene, a less thick lava flow stack), and Chilapa Fm. (Oligocene, intertongues the latter further north, it is largely lacustrine with abundant silicified limestone).
Dating and regional significance. Near Sayultepec (~20 km south- east), a tuff sheet interbedded in the Yanhuitlán Fm. yielded a Pb α age of 49.0±8.0 Ma (Ferrusquía-Villafranca, 1976). On the other hand, Martiny et al. (2000) reported a 40K-39Ar age of 40.5±1.7 Ma (Late Eocene) from an andesitic laccolith that intruded the Yanhuitlán Fm. near Marcos Pérez (~10 km NE of the area), thereby yielding a minimal Late Eocene age for this formation. Later, Cerca et al. (2007) reported a 40K-39Ar age of 43.2±1.2 Ma (Late Eocene) from the same laccolith. Finally, Santamaría-Díaz et al. (2008, p. 500, tab., figs. 2-3) reported a 40K-39Ar age of 35.7±1.0 Ma (latest Eocene) for the Cañada María Andesite, from a place situated ~4.5 km west of Tamazulapam (a town located ~18 km north of Yolomécatl). The Cañada María Andesite is an informal unit that overlies the Yanhuitlán Fm., thus providing a minimal age for the latter unit.
Summing up, in spite of their numerical differences, these four different ages (notwithstanding the large error of the first) place the Yanhuitlán Fm. in the Eocene. It remains to be proven whether the so called Yanhuitlán Fm. at Sayultepec, Marcos Pérez, Santa Catarina Tayata and Yolomécatl sites actually is the same unit. Whichever the case, such ages help to date the onset of the taphrogenic event subse-quent to the laramidic deformation in western Oaxaca, whereby the Cretaceous units became folded, faulted and uplifted, and horst and graben structurs developed.
The Yolomécatl l.f. (Appendix Part B, Table B2; Jiménez-Hidalgo et al., 2011) include five orders [Carnivora (one fam.), Creodonta (one fam.), Rodentia (two fam.), Perissodactyla (three fam.) and Artiodactyla (six fam.)] and 13 families; it has a Late Eocene outlook (?Duchesnian-?Chadronian), which is corroborated by the isotopic ages discussed above. This fauna increases the Early Tertiary North American fauna extent from central (Marfil l.f.) to southeastern Mexico (Oaxaca).
Early and Middle Miocene
Tubutama site
Sonora State, Northwestern plains and sierras MP, Figure 1, Number 5; Table 1, Number 5; Figure A6.
It is located in one of the basins that bear Tertiary fluvio-lacustrine sequences associated to volcanic emplacements. Arriaga-Meléndez et al. (1986) described the area, in the context of an economic geology (Borates) project. The pre-Cenozoic includes a Jurassic lithodem in- truded by a Cretaceous granitoid, overlain by a Late Cretaceous clastic marine succession, in turn covered by an up to ~1,120 m thick Cenozoic sequence consisting of six informal units (described upwards).
Unit 1. A latest Eocene-earliest Oligocene, ~150 m thick stack of andesitic and rhyolitic flows, pyroclastic breccias. Unit 2. A ~70 m thick, stratal pile of fine-grained arkosic sandstone and siltstone (lake facies) capped by a ~20 m thick, K-rich, palagonitized basaltic andesite flow (vitric-textured by underwater emplacement). Unit 3. A ~350 m thick, threefold stratal package: the lower part consists of red to orange, fine-grained sediments [largely clay and silt (lake facies)], intercalated by volcarenitic conglomerate lenses (fluvial, channel facies). The middle part consists of colemanite (CaB3O4(OH)3.H2O), gypsum, siltstone and fine grained sandstone strata (lake facies). The upper part largely consists of fresh water limestone. Unit 4. A ~ 200 m thick stratal pile lithologically similar to Unit 3, but more frequently includes limestone strata, and has little or no colemanite. Unit 5. A ~250 m thick stratal body formed by a red polymictic, matrix-supported, volcarentic conglomerate (debris flow and tallus facies). Unit 6. The Baucarit Fm. unconformably overlies the previous units; it consists of a Late Miocene basalt flow, covered by a ~100 m thick, volcarenitic, conglomeratic stratal body (fluvial facies). Finally, Quaternary alluvium deposits unconformably rest on this sequence.
Dating and regional significance. The palagonitized basalt flow yielded a 40K-40Ar age of 22.3 Ma (Arriaga-Meléndez et al., 1986, who cited as source an unpublished report for the Consejo de Recursos Minerales by Damon; Table 1), which places it in the earliest Miocene. Given that palagonitized basalts are not well suited for K-Ar dat- ing, this age is questionable. Within Unit 3, ~80 m above the basalt flow, we collected remains of the camel Stenomylus tubutammensis (Ferrusquía-Villafranca, 1990a). Stenomylus spans the Late Arikareean-Late Hemingfordian NALMA interval (Honey et al., 1998), however, intrinsic evidence from S. tubutamensis led the senior author to prefer an Early Hemingfordian age (Ferrusquía-Villafranca, 1990a), which is consistent with the isotopic age. S. tubutamensis is one of the south-ernmost stenomyline records in North America. Geologically, these isotopic and biochronologic ages date the beginning of basin develop- ment in northwestern Sonora.
Suchilquitongo site
Oaxaca State, Sierra Madre del Sur MP, Figure 1, Number 6; Table 1, Number 6; Figure A7.
The local Cenozoic sequence is located in the northern arm of the Valle de Oaxaca Graben (Ferrusquía-Villafranca and Ruiz-González, 2007), a Southeastern Mexico's major structural feature, which is bound to the west by the Late Proterozoic Oaxacan Complex and, to the east by the Late Paleozoic-Mid Mesozoic Sierra de Juárez Metamorphic Complex (also named Sierra de Juárez Mylonitic Complex; SGM, 2000) and partly, by folded and faulted Cretaceous marine carbonate units (SGM, 2007).
The Cenozoic includes andesitic lava flows at the base, and the Suchilquitongo Fm. (Wilson and Clabaugh, 1970), a ~750 m thick fluvio-lacustrine sequence formed by fine-grained subarkosic/tuffaceous sandstone, siltstone and claystone set in thin/medium strata (overbank facies), intercalated by thick strata of volcarenitic conglomerate (channel facies) and siliceous, fresh water limestone (lacustrine facies). Thin sheets of silicic ash-flow and ash-fall tuffs are interbed- ding sedimentary strata at various levels; the one that crops out in and around Etla constitutes the Etla Ignimbrite member, which commonly dips 15 °NE. The vertebrate fossils (Suchilquitongo l.f.) were collected from the fine-grained fluvial facies, ~80 m above this member (Ferrusquía-Villafranca, 2003). The Suchilquitongo Fm. records fluvial and lacustrine deposition in an actively subsiding basin. Quaternary deposits and soils unconformably overlie the previous units.
Dating and regional significance. The Etla Ignimbrite member yielded 40K-40Ar ages of 20.3±0.4 Ma and 19.2±0.5 (Ferrusquía-Villafranca and McDowell, 1991; Table 1), which place the Suchilquitongo Formation in the Early Miocene; additionally these ages allow us to date the initial development of the Valle de Oaxaca Graben (Ferrusquía-Villafranca and Ruiz-González, 2007, 2011), because: (a) this is the first and only emplacement of a thick pyroclastic sheet in the northern arm of this graben structure. (b) The pyroclastic sheets are confined to the graben (because none is present in the bounding horsts). Finally, these isotopic ages also contribute to date the Late Tertiary volcanism in the Sierra Madre del Sur M.P. (cf. Morán-Zenteno et al., 1999, 2000). The same applies for the Matatlán and Nejapa sites.
The Suchilquitongo l.f., (Appendix Part B, Table B3; Ferrusquía-Villafranca, 1990b, 2003) includes three orders [Carnivora (one fam.), Perissodactyla (two fam.) and Artiodactyla (four fam.)], seven families and 11 genera; its taxonomic makeup resembles that of the Late Arikareean-Early Hemingfordian faunas of temperate North America (e.g. Harrison, Marsland and Sheep Creek; cf. Tedford et al., 2004), accordingly its age falls within the Late Arikareean-Early Hemingfordian NALMAs (Tedford et al., 2004). This assignment is consistent with the presented isotopic ages. Finally, the Suchilquitongo l.f. from Oaxaca stands midway between the chrono-correlative faunas of North America (southern United States) and the Cucaracha l.f. from the Panama Canal Zone (Whitmore and Stewart, 1965; MacFadden et al., 2010) thus, corroborating the southward extension/continuity of the North America mammal fauna, during the Early Miocene.
Matatlán site
Oaxaca State, Sierra Madre del Sur MP, Figure 1, Number 7; Table 1, Number 7; Figure A8.
This Cenozoic sequence occupies the southeastern arm of the Valle de Oaxaca (Ferrusquía-Villafranca and Ruiz-González, 2007), unconformably overlying folded and faulted carbonate sequences of Albian and Late Cretaceous ages. The Cenozoic includes the Matatlán Fm. (Ferrusquía-Villafranca, 2003, fig. 13.4), a ~300 m thick, fluvio-lacustrine sequence consisting of four components: (a) fine-grained, cross-bedded tuffaceous sandstone and clayey siltstone set in thin to medium strata (fluvial, largely overbank facies). (b) Clast-supported, pebble to cobble volcarenitic conglomerate set in thick strata (alluvial fan and/or channel facies). (c) Laminar to thinly bedded clayey siltstone and silty claystone locally showing ripple marking (lacustrine facies). (d) Thin sheets of friable, felsic vitric ash-fall tuff interbedded at various stratal levels (pyroclastic facies).
This unit registers fluvial and lacustrine deposition in a rapidly subsiding basin with coeveal to intermittent pulses of silicic explosive volcanism (Ferrusquía-Villafranca and Ruiz-González, 2007). The fine-grained fluvial facies bears vertebrates (the Matatlán l.f.). The Matatlán Fm. intertongues and partly overlies the Mitla Tuff, a ~150-200 m thick volcanic succession formed by welded, vitric crystal to crystal vitric rhyolitic to rhyodacitic ash-flow tuff and pyroclastic surges. Quaternary deposits and soil crown the Cenozoic sequence.
Dating and regional significance. The Mitla Tuff yielded 40K-40Ar ages of 16.0±0.8 Ma and 15.3±0.8 Ma (Ferrusquía-Villafranca and McDowell, 1991; Table 1), thus placing it in the early Middle Miocene; that is, some 5 Ma younger than the Etla Ignimbrite of the Valle de Oaxaca Graben's northern arm, which contains a largely similar sedimentary/pyroclastic fill. Further, this age dates the final development of this graben (Ferrusquía-Villafranca and Ruiz-González, 2007, 2011) because (a) there are no volcanic successions covering the Mitla Tuff within or outside the graben. (b) There are no sedimentary deposits subsequent to such tuff and/or the Matatlán Fm., disclosing that the graben subsidence had ceased by this time (Middle Miocene), and with it the availability of accommodating space.
The Matatlán l.f. (Appendix Part B, Table B4; Ferrusquía-Villafranca, 1990b, 2003; Bravo-Cuevas and Ferrusquía-Villafranca, 2006, 2010) includes five orders [Carnivora (two fam.), Rodentia (one fam.), Proboscidea (one fam.), Perissodactyla (two fam.) and Artiodactyla (six fam.)], 12 families and 15 genera. Its taxonomic composition is comparable to that of the Early Barstovian faunas of temperate North America (e.g. Pawnee Creek, Deep River, Virgin Valley and Mascall; cf. Tedford et al., 2004), thus it is assigned to the Early Barstovian NALMA (Tedford et al., 2004); this biochronologic dating is further corroborated by the isotopic age discussed above. Finally, the fact that all Matatlán taxa show North American biogeographic affinities indicates that, at least during the Middle Miocene, the North American mammal fauna extended uninterruptedly southward down to Oaxaca.
Ixtapa site
Chiapas State, Sierra Madre de Chiapas MP, Figure 1, Number 8; Table 1, Number 8; Figure A9.
The local Cenozoic sequence unconformably overlies Late Cretaceous marine units; it includes a Paleocene-Middle Miocene, ~9,000 m thick, largely marine succession (Ferrusquía-Villafranca, 1996), deposited in a "pull-apart" basin (Mandujano-Velázquez and Keppie, 2009), placed in the Transcurrent-Fault Province of Chiapas (Sánchez-Montes de Oca et al., 1979); such basin is here interpreted as a fault-bend basin (cf. Busby and Ingersoll, 1995).
The Ixtapa Fm. unconformably overlies the previous succession; it is a ~3,500 m thick fluvio-lacustrine and subordinately lagoonal stratal sequence consisting of five components: (a) fine-grained, tabular cross-bedded, tuffaceous to volcarentic sandstone, clayey siltstone and silty friable claystone (chiefly overbank facies). (b) Clast-supported, pebble to cobble calcilithitic conglomerate (mainly channel facies). (c) Very fine-grained tuffaceous to volcarenitic, parallel-bedded tuffaceous to volcarenitic sandstone, clayey siltstone and silty claystone set in thin to laminar strata (lacustrine/pond facies). (d) Similarly made up strata bearing marine and continental palynomorphs (coastal lagoon facies; Martínez-Hernández, 1992). (e) Felsic ash-flow and ash-fall tuffs set in medium to thick strata, which make up about one third of this unit. The Ixtapa Fm. records fluvial and subordinately coast lagoonal deposition in a rapidly subsiding basin, coetaneous with explosive silicic volcanic activity. This unit is overlain by the Punta de Llano Formation (a Middle Pleistocene, pyro-epiclastic succession) and by other Quaternary deposits and soils.
Dating and regional significance. The tuff part of the Ixtapa Fm. yielded early-Middle Miocene 40K-40Ar ages of 16.02±0.53 Ma and 15.02±0.35 Ma (Ferrusquía-Villafranca, 1996; Table 1). Such ages date the structural deformation that generated the Transcurrent Fault Province of Chiapas.
The Ixtapa l.f. (Appendix Part B, Table B5) is a small assemblage of vertebrates collected from overbank facies beds lying below (~ 90 m) and above (~200 m) the dated tuff strata; it includes two orders [Proboscidea (one fam.) and Perissodactyla (tw o fam.)], three families and three genera (Langenheim and Frost, 1963; Ferrusquía-Villafranca, 1990c, 2003; Bravo-Cuevas and Ferrusquía-Villafranca, 2008). This fauna's makeup is comparable to that of the Early Barstovian faunas of temperate North America (e.g. Pawnee Creek, Mascall and Virgin Valley; cf. Tedford et al., 2004), hence it is assigned to the Early Barstovian NALMA (cf. Tedford et al., 2004), which is further corroborated by the isotopic ages discussed above. The Ixtapa l.f. discloses that during this time, the North American mammal fauna uninterruptedly extended southwards down to central-western Chiapas.
Nejapa site
Oaxaca State, Sierra Madre del Sur MP, Figure 1, Number 9; Table 1, Number 9; Figure A10.
This local Cenozoic sequence unconformably overlies a ?Paleozoic metamorphic complex and consists of the following units (Ferrusquía-Villafranca, 2001), mentioned in stratigraphic order:
Limón Conglomerate (Eocene, ~900 m thick, of phyllarenitic and calcilithitic composition). Yautepec Tuff and associated units (Middle Miocene, discordant over the Limón Cgl., a ~1,200 m thick, succession of felsic, largely welded, ash-flow tuffs, lapilli tuffs and surge emplacements; it partly intertongues and underlies El Camarón Fm. and laterally, it also intertongues the Huitihuini Rhyolite. The latter is a stack of lava flows with marked fluidal texture; it is intruded by the San José de Gracia unit, which forms stocks, apophyses and dykes of latitic to diabasic composition).
El Camarón Fm. is a Middle Miocene, ~250 m thick fluvio-lacustrine sequence consisting of three components: (a) fine-grained, tuffaceous to volcarenitic sandstone, clayey siltstone and silty claystone set in thin to medium strata (overbank facies; vertebrate fossils were chiefly collected from it). (b) A pebble to cobble volcarenitic conglomerate set in thick strata (mainly channel facies). (c) Fine-grained, parallel-bedded sandstone, clayey siltstone and silty claystone set in laminar to thin strata (lacustrine/pond facies). This unit registers fluvial/lacustrine deposition coeval to intense magmatic activity. Finally, Quaternary deposits (alluvium, colluvium) and soils unconformably cover the Tertiary sequence.
Dating and regional significance. The Yautepec Tuff yielded 40K-40Ar ages of 15.82±0.37 Ma, 15.78±0.37 Ma and 14.96±0.8 Ma (Ferrusquía-Villafranca, 2001; Table 1), thus placing this and its intertongued units (El Camarón Fm. and Huithuini Rhyolite) in the early Middle Miocene. These ages evidence that the Yautepec Tuff from the Oaxacan Subisthmian region and the Mitla Tuff from the Valle de Oaxaca, some 51 km apart, are coetaneous.
Additionally, it should be noted that the coeval Ixtapa, Perros (from the Isthmus of Tehuantepec; Ferrusquía-Villafranca, 1999), Yautepec and Mitla Tuffs disclose widespread Middle Miocene (ca. 15 Ma) volcanic/magmatic activity in a ~380 km long arc-like "belt," located at an average distance of 95 km north from the present position of the Middle American Trench/Cocos Plate subduction zone, and extending from central Oaxaca, over the Isthmian region to central-western Chiapas.
Such magmatic age-pattern does not support the east-ward Chortis Block displacement model of Southeastern Mexico's Cenozoic tectonic evolution (cf. Schaaf et al., 1995), nor the Cocos Plate leading edge model of Keppie and Morán-Zenteno (2005), thus paving the way for other alternative models (cf. Schulze et al., 2004; Nieto-Samaniego et al., 2006; Morán-Zenteno and Keppie, 2009; Silva-Romo and Mendoza- Rosales, 2008).
The Nejapa fauna (Appendix Part B, Table B6; Ferrusquía-Villafranca, 1990b, 2003; Bravo-Cuevas and Ferrusquía-Villafranca, 2006, 2010) includes five orders [Carnivora (two fam.), Rodentia (one fam.), Proboscidea (one fam.), Perissodactyla (two fam.) and Artiodactyla (eight fam.)], 14 families and 20 genera. This fauna is comparable to the Early Barstovian faunas of temperate North America (e.g. Pawnee Creek, Deep River, Virgin Valley and Mascall; cf. Tedford et al., 2004); thus it is assigned to the Early Barstovian NALMA (cf. Tedford et al., 2004), which is congruent with the isotopic ages discussed above. The Najapa fauna is more diverse than the Matatlán fauna, likewise it exclusively consists of North American taxa, a fact that confirms the southward extent of the North American land mammal fauna down to Southern Mexico, at least during the Middle Miocene.
Finally, it should be noted that the Miocene mammal fauna of Oaxaca includes lineages with both little derived and more derived taxa, which suggests that cladogenesis might have occurred there, i.e. southern tropical North America, and not only in temperate North America, as it is currently held; the Equini (Perissodactyla) and Kiptoceratinae (Artiodactyla) are two examples.
La Purísima site
Baja California Sur State, Baja California Peninsula MP, Figure 1, Number 10; Table 1, Number 10; Figure A11.
Following McLean and Hausback (1984), and McLean et al. (1985, 1987), the local Cenozoic succession includes the Late Oligocene San Gregorio Fm. (~70 m thick, diatomite, phosphatic shale/sandstone and thin vitric, biotite-bearing tuff sheets; 40K-40Ar dated as of 27-23 Ma), which is unconformably covered by the Middle Miocene, diachronous San Isidro/Comondú Fomations. The first (also spelled Ysidro) consists of marine, light yellowish gray, fine-grained sandstone, siltstone and shale, white coquina and a yellowish pebble conglomerate. It is very fossiliferous, its fauna is referred to the Vaqueros and Temblor Marine Stages, which in turn are nearly coetaneous with the Late Hemingfordian-Ealy Barstovian NALMAs (Barnes, 2002).
On the other hand, the Comondú unit has a long and involved nomenclatorial history [from Beal (1948), who first used the term to the present (cf. Umhoefer et al., 2001; Fletcher et al., 2007)] consequently, its conception, definition and usage varies enormously from one author to another (see review by Umhoeffer et al., 2001). The latter authors upgraded the Comondú Formation to group rank, their proposal, however, did not meet the requirements of the North American Stratigraphic Code (NACSN, 1983). Therefore, such change of rank remains informal. Subsequent use [e.g. Fletcher et al. (2007, p.1323) use Comondú Formation; fig.7] does not make it formal. Umhoefer et al. (2001) did not propose either the formations that their "Comondú Group" would include; instead, they propound these three informal units:
Lower clastic unit (200-300 m thick, coarse-grained, volcarenitic clastics sparsely intercalated by rhyolite tuff sheets and a few mafic lava flows; this unit has complex facies relations). Middle breccia and lava flow unit (largely dacitic to andesitic) and Upper lava flow and breccia unit (also largely dacitic to andesitic). The geologic interpretation of Umhoefer et al. (2001)is expressed in a regional map (1:500,000 scale), which does not include the La Purísma area; therefore, such interpretation is not directly applicable here. Further, these authors interpreted the Lower clastic unit as continental (fluvial and eolic), which is in contrast to other views that regarded it as largely marine or transitional (cf. McLean and Hausback, 1984; McLean et al., 1985, 1987). Certainly, in the La Purísima area our direct observations disclosed that the Comondú Formation includes both marine and transitional facies (Ferrusquía-Villafranca, 1990a).
In La Purísima area, a Middle-Late Miocene unnamed, mesa-forming basalt flow stack that yielded 40K-40Ar ages between 14.6- and 7.0 Ma (Pallares et al., 2008), unconformably overlies the Comondú Fm. Such stack is in turn overlain by a ?Pliocene unnamed unit (a hummocky relief-forming basalt flow stack). Quaternary basalt flows, cinder cones and alluvial deposits unconformably overlie the older units.
Dating and regional significance. Tuff sheets from the Comondú Fm. yielded Middle Miocene. 40K-40Ar ages of 13.0 and 14.5 Ma (McLean et al., 1987; Table 1). However, Umhoefer et al. (2001, p. 140) on the basis of their own 40Ar-39Ar ages and a critical appraisal of previous ages, indicate that at Loreto (~80 km ESE from La Purísima), the Lower clastic unit was laid down between ~21 and ~19 Ma (Early Miocene), but they also acknowledge that neither its lower nor upper boundaries are well constrained (due to the lack of suitable dating material at or close to them).
However, the reported age differences of the Comondú "Formation/ Group" at La Purísima and Loreto areas could adequately be explained by these facts: (a) insufficient constraint of this unit's lower and upper boundaries. (b) The diachroneity of this unit. (c) The Late Miocene U-Pb ages (19% of zircon crystals from a sample of the Comondú "Formation/Group" collected in a locality placed 29 km west of La Paz and ~310 km SSE from La Purísima, yielded ages as young as 9 Ma) reported by Fletcher et al. (2007, p. 1313, fig. 7). (d) The possibility that the so called Comondú unit could not be the same rock body at these three places. It follows that much detailed geologic work remains to be done to correctly settle this issue.
At any rate, from a locality where the San Isidro and Comondú Formations intertongue, one of us recovered terrestrial and marine mammals (Ferrusquía-Villafranca, 1990a): Euoplocyon praedator (Carnivora, Canidae) and Desmostylus hesperus (Desmostylya, Desmostylidae) respectively. Euplocyon's biochronologic range spans the Late Hemingfordian-Early Barstovian (Tedford et al., 2004), but it is best known from the Early Barstovian Green Hills fauna of California (Woodburne et al., 1990). On the other hand, the marine mammal Desmostylus is known in North America from the Early and Middle Miocene (McKenna and Bell, 1997), an interval roughly corresponding to the Late Arikareean-Barstovian NALMAs (cf. Tedford et al., 2004). It follows that their co-occurrence would more probably fall in the Late Hemingfordian-Early Barstovian time interval. However, the isotopic ages obtained from the Comondú tuff sheets and overlying mesa-basalts at La Purísima, allow us to constrain the interval to ca. 14 Ma, which falls in the Early Barstovian NALMA (Tedford et al., 2004) and La Purísima 1.f. is assigned to this age. Finally, it should be noted that this site is well suited to investigate direct correlation of marine and terrestrial stratal units and fossil assemblages in western North America (Ferrusquía-Villafranca, 1990a), and on this regard alone, it is very important.
Late Miocene
Paso del Águila site
San Luis Potosí State, Sierra Madre Oriental MP, Figure 1, Number 11; Table 1, Number 11; Figure A12.
The local Cenozoic succession unconformably overlies Late Cretaceous marine units; it consists of two Oligocene volcanic units in the lower part and the San Nicolás Fm. (Ferrusquía-Villafranca et al., 2015) that overlies them. The latter is a ~1,100 m thick, largely fluvial sequence that includes five components: (a) immature, calcilithitic, frequently cross-bedded sandstone, clayey siltstone and silty claystone set in thin to medium strata (channel and overbank facies). (b) Clast-supported, pebble to cobble, chiefly calcilithitic conglomerate set in medium to very thick strata (channel facies). (c) Very fine-grained, parallel-bedded calcilithitic sandstone, siltstone and claystone set in thin strata (overbank facies), it is sparsely fossiliferous. (d) Subordinate amounts of very fine-grained, parallel-bedded sandstone, clayey siltstone and claystone set in thin to very thin strata, interbedded by fresh water limestone (lacustrine/pond facies). (e) A few thin sheets of felsic, vitric, ash-fall tuff, interbedded at various stratigraphic levels (pyroclastic facies). This unit records largely fluvial -and subordinately lacustrine-deposition in a rapidly subsiding basin, coetaneous to intermittent silicic volcanic activity. The San Nicolás Formation is unconformably overlain by Quaternary deposits and soils.
Dating and regional significance. The interbedded San Nicolás tuff sheets yielded Late Miocene 40Ar-39Ar ages of 10.99±0.22 Ma and 7.41±0.31 Ma (Ferrusquía-Villafranca et al., 2014; Table 1). These ages contribute to date a Late Tertiary, short-lived episode of continental deposition in a western graben of the Sierra Madre Oriental M.P., the only one known from this province so far. This example proves that the holistic study of sedimentary fills in other graben structures would improve the understanding of the Late Cenozoic geologic/tectonic evolution of the Sierra Madre Oriental.
On the other hand, The Paso del Águila l.f. (Appendix Part B, Table B7; Ferrusquía-Villafranca et al., 2014) includes two orders [Perissodactyla (one fam.) and Artiodactyla (two fam.)], three families and three genera of mammals, and a reptile (Chelonia). This fauna resembles the Late Clarendonian-Early Hemphillian faunas of temperate North America (e.g. McGehee Farm, Mixon, and Dove Springs; cf. Tedford et al., 2004). In fact, the perissodactyl Pliohippus potosinus displays a similar degree of cheek teeth development (in size and morphology) than Late Clarendonian-Early Hemphillian Pliohippus species (cf. Kelly, 1998), hence leading us to assign this fauna to this interval, an interpretation further corroborated by the isotopic ages discussed above. The lack of any South American taxon in this fauna strongly suggests that the Panamian land connection was still not completed then.
Chapala site
Jalisco State, Trans-Mexican Volcanic Belt MP (TMVB hereafter), Figure 1, Number 12; Table 1, Number 12; Figure A13.
This site is located in the Chapala Rift (first described by Clemens, 1963), in the western part of the TMVB, particularly on the northern shore of Lake Chapala. Practically, there is no published detailed geologic information on the area, particularly for the sedimentary sequence that bears the vertebrate fossils. The exception is the geological map of INEGI (1976), which only discriminates lithic units without a time frame. On the other hand, the tectonic and volcanologic literature on this part of the TMBV is abundant (cf. Ferrari et al., 2012, and references therein). The geologic summary given below, chiefly draws from Clemens (1963), INEGI (1976, 1988) and SGM (1992).
The basal unit is a rhyolitic body, which seemingly yielded K-Ar ages of 6.2 Ma and 3.4±0.2 Ma (Lucas, 2008, p. 40, fig. 7). However, the author does not state the source of the first age and attributes the second to Delgado-Granados et al. (1995), Rosas-Elguera et al. (1996) and Ferrari et al. (2000). Actually, the younger age is from a basalt (Rosas-Elguera and Urrutia-Fucuguchi, 1998, tab. 1). The same authors also published a 6.26±1.21 Ma age from another basalt, that might be the source of the first age cited above.
This basal unit is non-conformably overlain by the vertebrate-bearing Chapala Fm., an informal unit proposed by Clemens (1963) without satisfying the pertinent requirements of ACSN (1961).
The Chapala Fm. is a ~600 m thick sequence of thickly bedded claystone, siltstone and partly diatomaceous shale, thinly bedded fresh water silty limestone (lacustrine facies); sparse thin interbeds of felsic ash-fall tuff (pyroclastic facies); and lesser amounts of cross-bedded volcarenitic sandstone and clast-supported conglomerate (fluvial facies); it commonly dips 20°-30°NE. (lithic description updated by the authors). Quaternary alluvial deposits overlie this unit.
Dating and regional significance. Israde-Alcántara et al. (2010, p. 88) reported a 40Ar-39Ar age of 7.0±0.3 Ma (Table 1) from a laccolith that intruded the sedimentary sequence (i.e., the Chapala Formation), thus placing it at least in the Late Miocene; however, there are no vertebrate fossils known from this part of the sequence. These authors also reported a 40Ar-39Ar age of 5.2±0.3 Ma (Table 1) of a tuff sheet associated to diatom-bearing strata, which places the Chapala Formation in the earliest Pliocene. Again, no vertebrates are known from this or associated beds in the sequence. On the other hand, such ages are concordant with the current interpretation of the tectonic evolution of the western TMVB (cf. Delgado-Granados, 1992, 1993; Rosas-Elguera et al., 1996; Michaud et al., 2000; Ferrari et al., 2000).
Finally, the presence of pre-Pleistocene mammals of Blancan age in Chapala is known long ago (cf. Downs, 1958a, 1958b; Ferrusquía-Villafranca, 1978; Lucas, 2008), however, their precise stratigraphic provenance is not known, nor have they been associated to isotopic ages. The Pleistocene mammal fauna from this site is summarized and discussed in Ferrusquía-Villafranca et al., 2010.
Tepezalá area site
Zacatecas State, Sierra Madre Occidental MP, Figure 1,
Number 13; Table 1, Number 13; Figure A14.
Cofradía area site
Zacatecas State, Sierra Madre Occidental MP, Figure 1,
Number 14; Table 1, Number 14; Figure A14.
El Resbalón area site
Zacatecas State, Sierras Madre Occidental MP, Figure 1, Number 15; Table 1, Number 15; Figure A14.
These sites are located in the southern Sierra Madre Occidental M.P., within the Juchipila Basin, one of the several NNE-SSE trending grabens or half grabens present there (cf. INEGI, 1974; SGM, 1991) developed in the Oligocene Upper Volcanic Complex (sensu McDowell and Keizer, 1977; McDowell and Clabaugh, 1979). Isotopic dating of tuff sheets of the horsts associated to the graben structures of the Juchipila Basin yielded ages between 25.9±2.5 Ma and 24.9±2.7 Ma (latest Oligocene, fission track method; Webber et al., 1994), which would be the maximal age of graben development.
The sedimentary graben fill includes the Late Miocene-Early Pliocene Juchipila Fm. (Lahiere, 1982; López, 1991; a unit never formally proposed), a ~90 m thick fluvio-lacustrine sequence formed by parallel-bedded, fresh water limestone and very fine-grained clastics (lacustrine facies), interbedded by frequently cross-bedded volcarenitic sandstone and siltstone set in thin to medium bedded strata (mainly overbank and channel facies; vertebrate fossils were collected from this facies). Thin sheets of felsic ash-fall tuff (pyroclastic facies) interbed the sedimentary sequence at various levels. Finally, the Juchipila Fm. is unconformably overlain by Quaternary gravel deposits.
Dating and regional significance. The tuff sheets in these sites yielded U-Pb zircon ages ranging from 6.95±0.11 Ma (in Tepezalá, Carranza-Castañeda et al., 2013; Table 1), to 6.53±0.11 Ma (in La Cofradía, Carranza-Castañeda et al., 2013) and to 5.59±0.11 Ma (in El Resbalón, Carranza-Castañeda et al., 2013; Table 1), which gives latest Miocene-Early Pliocene ages for the Juchipila Fm. These ages also contribute to date Late Cenozoic pulses of volcanic activity and structural deformation in the southern Sierra Madre Occidental M.P., as well as continental depositions in their grabens/half grabens.
The local faunas of these sites constitute the Juchipila fauna (Appendix Part B, Table B8), which includes five orders [Xenarthra (one fam.), Carnivora (one fam.), Proboscidea (one fam.), Perissodactyla (two fam.) and Artiodactyla (four fam.)], nine families and 16 genera (Carranza-Castañeda et al., 2013). Its taxonomic content closely resembles that of the Late Hemphillian faunas of temperate North America [e.g. Draw, Nebraska (Skinner et al., 1977), Wikieup, Arizona (Morgan and White, 2005), Upper Bone Valley, Florida (Hulbert, 1987)], which allows to assign it to the early Late Hemphillian (Hh2) NALMA (cf. Tedford et al., 2004). The above mentioned isotopic age corroborates this interpretation.
Finally, in Mexico, the Juchipila fauna correlates with coeval faunas of Guanajuato, Nayarit and Jalisco (see below), which also include South American xenathrans, thus heralding the Great American Biotic Interchange (GABI hereafter), a phenomenon known long ago (Scott, 1937; Ferrusquía-Villafranca, 1978), but much later named so (cf. Stehli and Webb, 1985).
El Trapiche site
Nayarit State, Sierra Madre Occidental MP, Figure 1, Number 16;
Table 1, Number 16; Figure A15.
Rancho El Ocote (=REO)-GTO2A site; REO-GTO 2C
Both in: Guanajuato State, Central Plateau MP, Figure 1,
Number 17; Table 1, Number 17; Figure A16.
Tecolotlán site
Jalisco State, Trans-Mexican Volcanic Belt MP, Figure 1, Number 18; Table 1, Number 18; Figure A17.
Published detailed geologic information for these sites/areas is scarce, and no lithostratigraphic units have been proposed. The brief summary presented below draws from SGM (1991, 2006, 2014).
These sites are located in the Late Cenozoic NNE-SSW elongate basins set in graben structures developed in the southern part of the CeP (San Miguel de Allende Graben), the southwestern most part of the SMOc (El Trapiche Graben), and the western most part of the TMVB (Tecolotlán Graben). It should be noted that under close examination, the Trapiche Graben shown by Carranza-Castañeda et al. (2013, fig. 2, seemingly a DEM -Digital Elevation Model-without explicit latitude and longitude coordinates) corresponds to a topographic depression, actually a valley with steep slopes and without topographic discontinuities (faults) that would characterize it as a graben structure. In contrast, the San Miguel de Allende and Tecolotlán Grabens are clearly bounded by faults (Carranza-Castañeda et al., 2013, fig. 3; SGM, 2014).
The sedimentary fill of these grabens and topographic depressions is formed by fluvio-lacustrine sequences a few dozens of meters thick, consisting of four components: (a) fine to coarse-grained, frequently cross-bedded, commonly volcarenitic sandstone, siltstone and claystone set in thin to medium strata (fluvial facies). (b) Clast-supported, pebble to cobble conglomerate set in thick strata (fluvial, chiefly channel and fan facies). (c) Parallel-bedded, fine siltstone and silty claystone, and unfrequent calcareous claystone beds bearing ostracods and gastropods set in thin strata (lacustrine facies). (d) Thin sheets of felsic ash-fall tuffs that interbed the sequence at various levels (pyroclastic facies).
It should be noted that these sequences are locally designated as follows: (a) TnAr-Cgp unit (Tertiary Neogene sandstone-polymictic conglomerate; SGM, 1999b) in the San Miguel de Allende area. (b) Tpl.Cgp-Ar unit (Tertiary Pliocene-polymictic conglomerate and sandstone; SGM, 2014) in the Tecolotlán area. (c) In the Trapiche area, the local sequence is so small that it does not show up in the 1:50,000 scale geological map (cf. INEGI, 1973; SGM, 2006). Finally, all these sequences record fluvial and/or lacustrine deposition taking place coetaneously with pulses of explosive silicic volcanism.
Dating and regional significance. The tuff sheets yielded the following isotopic ages: (a) 5.7±0.4 Ma (GTO2A site, Kowallis et al., 1998; fission track method; Table 1). (b) 5.50±0.05 Ma to 5.12±0.26 (Trapiche site, Carranza-Castañeda et al., 2013; U-Pb zircon method; Table 1). (c) 4.989±0.16 Ma to 4.89±0.16 Ma (Tecolotlán site, Flynn et al., 2005; 40Ar-39Ar method). (d) 4.8±0.2 Ma (REO-GTO2C site, Kowallis et al., 1998; fission track method; Table 1). Such ages place these sequences in the Late to latest Miocene-earliest Pliocene; they also contribute to date Late Cenozoic volcanism, faulting and continental deposition in the regions/areas of these sites.
The combined mammalian assemblage of these local faunas (Appendix Part B, Table B9) includes five orders [Xenarthra (one fam.), Carnivora (four fam.), Proboscidea (one fam.), Perissodactyla (two fam.) and Artiodactyla (four fam.)], 13 families (all may not occur in a given site) and 19 genera (Carranza-Castañeda, 2006; Carranza-Castañeda et al., 2013). Its taxonomic makeup is comparable to that of Late Hemphillian faunas of temperate North America [e.g. Modesto Reservoir, California (Wagner, 1976), Edson Quarry, Kansas (Harrison, 1983), Coffee Ranch (Dalquest, 1969)], therefore it is assigned to the latest Hemphillian NALMA (cf. Tedford et al., 2004). The isotopic ages cited above corroborate these finding. As in the faunas from Zacatecas, the salient fact is the presence of South American immigrant taxa (Megalonychidae), which evidence GABI's initial phase. Finally, it should be stressed that the San Miguel de Allende sequence also carries at its upper levels numerous Blancan mammal taxa, thus it is well suited to investigate Late Cenozoic biochronologic succession and faunal turnover in central Mexico. To a lesser extent, this also applies to the Tecolotlán sequence.
Pliocene
GTO 43 site
Guanajuato State, Central Plateau MP, Figure 1, Number 19;
Table 1, Number 17; Figure A16.
Rancho El Ocote, GTO 2D site
Guanajuato State, Central Plateau MP, Figure 1, Number 19;
Table 1, Number 17; Figure A16.
Garbani, GTO 11 site
Guanajuato State, Central Plateau MP, Figure 1, Number 20;
Table 1, Number 20; Figure A16.
La Pantera, GTO 12B site
Guanajuato State, Central Plateau MP, Figure 1, Number 21;
Table 1, Number 21; Figure A16.
Estancia, GTO 5A site
Guanajuato State, Central Plateau MP, Figure 1, Number 22;
Table 1, Number 22; Figure A16.
Rancho El Ocote, GTO 55 site
Guanajuato State, Central Plateau MP, Figure 1, Number 17; Table 1, Number 17; Figure A16.
These six sites are located in the San Miguel de Allende Graben, whose sedimentary fill was already described elsewhere (Rancho el Ocote and related sites above).
Dating and regional significance. The tuff sheets associated to the fossiliferous strata yielded the following ages: (a) 4.747±0.032 Ma (Rancho El Ocote, GTO 2D site, Flynn et al., 2005; 40Ar-39Ar method; Table 1), 4.7±0.032 Ma (same site and reference as above) and 4.6±0.3 Ma (same site, Kowallis et al., 1998; fission track method; Table 1). (b) 4.4±0.3 Ma (GTO 43 site, Kowallis et al., 1998; same method; Table 1). (c) 4.1±0.5 Ma [Garbani, GTO 11 site, same reference and method as in (b); Table 1]. (d) 3.9±0.3 Ma [La Pantera, GTO 12B site, same reference and method as in (b); Table 1]. (e) 3.32±0.02 Ma (Estancia, GTO 5A site, Flynn et al., 2005; 40Ar-39Ar method; Table 1). (f) 3.08±0.5 Ma (GTO 55 site, Kowallis et al., 1998; fission track method; Table 1). These ages place the local sequences in the late Early-Middle Pliocene and also contribute to date Late Cenozoic structural deformation and fluvio-lacustrine deposition in the southern part of the Central Plateau M.P.
The combined assemblage from these local faunas, herewith designated San Miguel fauna (Appendix Part B, Table B10; cf. Carranza-Castañeda, 2006 and references therein) includes: seven orders [Xenarthra (one fam.), Lagomorpha (one fam.), Rodentia (two fam.), Carnivores (two fam.), Proboscidea (one fam.), Perissodactyl (one fam.) and Artiodactyla (three fam.)], 11 families and 25 genera; its taxonomic makeup is comparable to that of temperate North American Blancan faunas [e.g. Vallecito Creek and Fish Creek, California (Downs and White, 1968; Cassiliano, 1999) Buckeye Creek, Nevada (Kelly, 1994) and Hagerman, Idaho (Mc Donald et al., 1996 and references therein)]. Hence, the San Miguel Fauna is assigned to the Early and Middle Blancan NALMA (cf. Bell et al., 2004). The isotopic ages cited above corroborate the biochronologic age information.
The presence of two South American immigrant families (Megalonychidae and Hydrochoeridae) in Guanajuato, evidences that GABI was then in full swing. On the other hand, on the basis of the above cited ages, Flynn et al. (2005) argued that: (a) the Hemphillian/ Blancan Boundary at Guanajuato should be placed at ~4.7-4.8 Ma. (b) The Guanajuato South American record predates for some 1.5-1.2 Ma GABI's initial record in temperate North America. However, this can be questioned, because there are earlier Late Miocene records of megalonychids there (cf. Woodburne et al., 2006 and references therein). Other facts also suggests the need to revise GABI's initiation, such as the presence of Late Miocene North American mammals in South America (e.g. the long known cyonasuan carnivores (Pascual, 1972) and the seemingly earlier proboscidean record (Campbell et al., 2010). However, such kind of revision is beyond the scope of this paper.
Finally, given that the Rancho El Ocote stratal succession at lower levels also carries Hemphillian mammals, it is well suited for studying biochronologic succession and faunal turnover, as mentioned above.
Santa María Amajac site
Hidalgo State, Trans-Mexican Volcanic Belt MP, Figure 1,
Number 23; Table 1, Number 23; Figure A18.
Charo site
Michoacán State, Trans-Mexican Volcanic Belt MP, Figure 1, Number 24; Table 1, Number 24; Figure A19.
The description of the first site is largely taken from Segerstrom (1956, 1962), and SGM (1995a). The local sequence includes the Late Tertiary volcanic Pachuca Group, which is unconformably overlain by the Atotonilco El Grande Formation, an informal unit proposed by Segestrom (1956) without fulfilling the then accepted requirements (Ashley et al., 1933); such unit remains informal, because in spite of its subsequent use (cf. de Cserna et al., 1987; Arellano-Gil et al., 2005); no one has ever formally proposed it since that time.
The Atotonilco El Grande Fm. is a ~500-600 m thick volcanic and epiclastic body that intertongues the Pliocene Navajas Rhyolite. The epiclastic part is a fluvio-lacustrine sequence, which besides the expected channel and overbank deposits, also includes fresh water limestone (lacustrine facies), lignite and peat (palludal facies). It carries vertebrates, and is interbedded by thin rhyolitic ash-fall tuff sheets. This unit is overlain by Quaternary deposits.
On the other hand, the epiclastic part of this formation was described in more detail by Arellano-Gil et al. (2005) and according to these authors, it is a 151.4 m thick sequence of volcarenitic [their "graywackes" (grauvacas)] conglomerate, sandstone (coarse to fine-grained) and siltstone set in medium to thick strata, sparsely intercalated by tuff sheets. The conglomerate may form lenses or tabular bodies that show cross bedding (channel facies). The siltstone may show lamination and bear well preserved leaves, diatoms and ostracods (lacustrine facies).
As for the Charo site, the published detailed geologic information is also scarce. The following summary is taken from INEGI (1983), SGM (1995b), Silva-Mora (1995) and Israde-Alcántara and Garduño-Monroy (1999). The site is located in north-east Michoacán, some 15 km ENE of Morelia. It is located in a small Late Cenozoic graben, where a ~100 m thick, fluvio-lacustrine sequence was deposited; it non-conformably overlies the Miocene Mil Cumbres Complex (locally its age ranges from 8±0.2 to 13.0 Ma; K-Ar method; Israde-Alcántara and Garduño-Monroy, 1999).
The sedimentary sequence includes four components: (a) tabular cross-bedded volcarenitic sandstone and clayey siltstone set in thin to medium strata (channel and overbank facies, a few vertebrate fossils are present here). (b) Clast-supported, volcarenitic conglomerate forms lenses throughout the sequence (channel facies). (c) Parallel-bedded siltstone, claystone and diatomite (lacustrine facies), which constitute nearly one half of the sequence. (d) A few interbedded thin felsic ash-fall tuff sheets. Quaternary fine-grained clastic deposits that bear Pleistocene mammals, unconformably overlie the Pliocene sequence (Carranza-Castañeda, 2006).
Dating and regional significance. The ash-fall tuff sheets in Amajac have yielded the following isotopic ages: (a) 4.591±0.007 Ma (Flynn et al., 2005; 40Ar-39Ar method; Table 1). (b) 4.57±0.02 Ma [same reference and method as in (a), Table 1]. (c) 4.2±0.3 Ma (Kowallis et al., 1998; fission track method; Table 1). On the other hand, in Charo only a single tuff sheet was independently dated twice: (a) 3.6±0.3 Ma (Kowallis et al., 1998; fission track method). (b) 3.6 Ma (Israde-Alcántara and Garduño-Monroy, 1999; K-Ar method; Table 1). All these ages place the Amajac and Charo sequences in the Early-Middle Pliocene (although, that of Amajac is slightly older), and correlate them with sequences of the San Miguel de Allende area, as discussed above. These ages also contribute to date Late Cenozoic volcanism and structural deformation in the areas where the sites are located.
The combined Santa María Amajac-Charo mammalian assemblage (Appendix Part B, Table B11; Repenning, 1962, and Carranza-Castañeda, 2006 and references therein) includes six orders [Lagomorpha (one fam.), Rodentia (two fam.), Carnivora (one fam.), Proboscidea (one fam.), Perissodactyla (one fam.) and Artiodactyla (three fam.)], nine families and 14 genera; its taxonomic composition resembles that of the Blancan faunas from temperate North America [e.g. Hagerman, Idaho (McDonald et al., 1996); Taunton and White Bluffs, Washington (Repenning et al., 1995)]. Therefore, this assemblage is assigned to the Blancan NALMA (cf. Bell et al., 2004). The isotopic ages cited above corroborate these findings. Finally, this combined assemblage is less diverse than the San Miguel fauna, and as yet, it does not include South American immigrant taxa.
Buenaventura site
Jalisco State, Trans-Mexican Volcanic Belt MP, Figure 1, Number 25; Table 1, Number 25; Figure A17.
This site is located in the Tecolotlán Basin, which was already described in site #18, thus we just add that the Hemphillian mammal-bearing sequence is unconformably overlain by another fluvio-lacus-trine sequence carrying Blancan mammals. It is sparsely interbedded by thin, felsic ash-fall tuff sheets. We are focusing here on the upper sequence.
Dating and regional significance. Samples from the tuff sheets yielded a fission track age of 2.6±0.6 Ma (Kowallis et al., 1998; Table 1), thus placing it in the Late Pliocene. The Buenaventura l.f. (Appendix Part B, Table B12) is small and includes only two orders [Perissodactyla (one fam.) and Artiodactyla (two fam.)], three families and three genera. It resemblances the Late Blancan faunas of temperate North America (e.g. 111 Ranch, Arizona, Galusha et al., 1984), therefore, we assign it to the Blancan NALMA (Bell et al., 2004). The above mentioned fission track age, corroborates this interpretation. The possibility to investigate biochronologic succession in this basin was already mentioned.
SUMMARY AND CONCLUSIONS
We described/discussed the isotopic and paleontologic information of 28 sites across the country, their distribution in space and time is very uneven: BCP, one Cretaceous and one Miocene sites. NW, one Miocene site. SMOc, four Miocene sites. CeP, one Eocene and eight Miocene/Pliocene sites. SMOr, one Jurassic and one Miocene sites. TMVB, two Miocene and three Pliocene sites. SMS, one Eocene and three Miocene sites. SMCh, one Miocene site. Patchiness/skewness aside, the results are quite significant.
Mesozoic. The Pliensbachian Huizachal site (SMOr) is located in the rift system that split central-eastern Pangea, records associated volcanism and bears Mexico's oldest vertebrate fauna, which is fairly diverse and befittingly shows laurentic and gondwanic biogeographic affinities. The Rosario site (BCP) evidences Late Campanian fluvial (and subsidiarily tidal) deposition coetaneous to volcanism in the Peninsular Ranges Forearc Basin (or El Rosario Forearc Basin); its Judithian fauna shows western North American biogeographic affinities, as well as some degree of endemism (particularly among the dinosaurs).
Cenozoic. The Cenozoic sites lie in several graben structures and record episodes of deposition as well as tectonic and magmatic activity. Eocene-Early Miocene: The Marfil (CeP) and Yolomécatl (SMS) sites are located in basins associated to post-Laramide relaxation and magmatism; their Bridgerian and ?Duchesnian/?Chadronian faunas evidence the southward extent of the North American Paleogene vertebrate fauna. Some taxa however, show strong endemism. The Tubutama (NW) and Suchilquitongo (SMS) sites record deposition associated to tectonic and magmatic activity (taking place by 20 Ma); their faunas show North American biogeographic affinities and endemism seemingly related to isolation.
Middle Miocene. The La Purísima (BCP), Matatlán (SMS), El Camarón (SMS) and Ixtapa (SMCh) sites record deposition related to tectonic and magmatic activity (occurring at ca. 15 Ma). The Valle de Oaxaca Suchilquitongo and Matatlán sites disclose southward migration of such activity. Elsewhere in Southeastern Mexico, this process generated a volcanic belt extending from the southeastern Valle de Oaxaca to northern Chiapas. Their faunas evidence the southern extent of North America's fauna at that time. Some taxa (e.g. Equini and Kyptoceratinae) though, show that their cladogenesis (partly at least) took place in tropical southern North America, rather than in temperate "northern" North America, as it is currently thought.
Late Miocene and Pliocene. The Paso del Águila (SMOr), El Resbalón, Cofradía, Tepezalá, El Trapiche (SMOc), Rancho El Ocote (CeP) and Tecolotlán (TMVB) sites record deposition coeval to tectonic and volcanic activity (ca. 9.5-5 Ma). Their mammal assemblages includes two chronofaunas: Clarendonian-Hemphillian (one record, the first in Mexico) and Hemphillian (eight records, they carry some of the oldest registers of South American taxa in North America, thus heralding the full swing of the Great American Biotic Interchange). The nine Pliocene sites (CeP, 6; TMVB, 3) record deposition associated to tectonic and volcanic activity (taking place around 4.5-3.5 Ma), and their Blancan chronofauna includes numerous South American taxa (which indicates that GABI was at its peak then.
In spite of the large geographic spread (15 ° of latitude, spanning temperate and tropical zones/regions of diverse climate regimes and biomic frameworks) and elapsed time (from Late Cretaceous to Pliocene) displayed by the 27 investigated sites that bear both isotopically dated material and studied terrestrial vertebrate faunas, the North American Land Mammal Age System yielded biochronologic ages consistent with the isotopic ages. This cumulative evidence lends objective support to the empirical practice of assuming the idoneity/ suitability of such system to adequately date the mammal faunas of Mexico, regardless the fact that it was developed from stratigraphic successions of terrestrial mammal faunas that lived in temperate North America (largely southern Canada and the contiguous United States). This is indeed a major contribution to vertebrate paleontology in this country.
Concluding remarks. This brief review on the combined isotopic/ vertebrate paleontologic approach applied in Mexico during the last fifty years, an effort developed through short term independent projects, shows that it has nonetheless yielded valuable, albeit patchy geologic/paleontologic results. Clearly, the stage is set to undertake long-range, multidisciplinary/interinstitutional projects across the country, whose outcome could be a many-fold knowledge increase that would qualitatively improve our understanding on the geologic/ biotic history and evolution of Mexico.

