











 nueva página del texto (beta)
nueva página del texto (beta) Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
Permalink
Allium glandulosum Link & Otto, es una especie con amplia distribución geográfica, que ocupa ambientes contrastantes en Norte América, desde el sur de Estados Unidos, hasta Centroamérica (Traub 1968, Espejo-Serna & López-Ferrari 2003, Galván 2010). Dado el patrón de distribución geográfica y la presencia en diferentes condiciones ambientales de esta especie, se espera que A. glandulosum presente variación morfológica vegetativa y floral, como ocurre en otros grupos taxonómicos con patrones de distribución similares, como algunos miembros de Boraginaceae (Cavieres 2000); Cactaceae (Vázquez-Benítez et al. 2016); Pinaceae (Reyes-Hernández et al. 2005, Sáenz-Romero et al. 2012) y Taxaceae (Ramírez-Sánchez et al. 2011). En la última revisión taxonómica de Allium para México (Traub 1968), existe incertidumbre sobre los caracteres útiles para la identificación de las especies. Esta situación ha generado que actualmente existan especímenes mal identificados bajo el nombre de A. glandulosum. Esta especie carece de estudios que permitan documentar su variación morfológica a través de métodos objetivos. Los análisis morfométricos son estudios respaldados con técnicas multivariadas, que han demostrado su utilidad para la delimitación y reconocimiento de taxones con amplia variación morfológica en familias de monocotiledóneas como Amaryllidaceae (Díaz-Lifante & Andrés-Camacho 2007, El-Hadidy et al. 2012), Arecaceae (Henderson 2002), Asparagaceae (Preston 2013, Gutiérrez et al. 2017), Bromeliaceae (Pinzón et al. 2011), Orchidaceae (Pedersen 2010) y Potamogetonaceae (Kaplan & Marhold 2012). Eudicotiledóneas como Asteraceae (Villareal-Quintanilla et al. 2020), Cactaceae (Arroyo-Cosultchi et al. 2010, Sánchez et al. 2013, Vázquez-Benítez et al. 2016), Fagaceae (Martínez-Cabrera et al. 2011), Polygonaceae (Ancona et al. 2019) y Solanaceae (Castillo-Batista et al. 2017). Además de monilophytas (Hernández-Hernández et al. 2009, Pérez-Atilano et al. 2017).
Para el estudio de Allium se han utilizado métodos típicos de análisis morfométricos, como el análisis de componentes principales y de conglomerados; que se han utilizado con el objetivo de agrupar y delimitar especies relacionadas (El-Gadi & Elkington 1977, Badr & Elkington 1978, Jacobsen 1979). El-Gadi & Elkington (1977), así como Badr & Elkington (1978) utilizaron caracteres citológicos, morfológicos y químicos para estudiar las relaciones fenéticas entre especies de los subgéneros Rhizirideum (G. Don ex Koch) Wendelbo y Molium Koch (Wendelbo). Jacobsen (1979) analizó la morfología floral y vegetativa, así como la fenología, hábitat y distribución geográfica para esclarecer los límites taxonómicos entre Allium cernuum Roth y A. stellatum Nutt ex Ker Gawl. Con base en los antecedentes de trabajos morfométricos, que son relevantes para la taxonomía de Allium y otros grupos, así como la amplia distribución de A. glandulosum, el objetivo de esta investigación es determinar mediante análisis multivariados, si estructuras vegetativas y florales de A. glandulosum presentan variación morfológica significativa entre poblaciones, con el fin de determinar el o los caracteres morfológicos que permitan el reconocimiento de poblaciones de esta especie.
Materiales y métodos
Selección de poblaciones representativas. La revisión preliminar de la distribución geográfica de A. glandulosum en México, permitió obtener sitios de recolecta a partir de ejemplares depositados en los herbarios CHAPA y UAMIZ (Thiers 2023) y de registros de bases de datos (GBIF 2022, DGRU 2023). Se recolectaron 50 ejemplares en ocho estados de la república mexicana en 15 sitios en los que se identificó la presencia de A. glandulosum (Tabla 1), durante los meses de agosto y septiembre de 2020.
Tabla 1 Sitios de recolecta de material vegetal de Allium glandulosum Link & Otto.
| Estado | Municipio | Tipo de vegetación | Altitud (m snm) |
|---|---|---|---|
| Durango | Durango | Bosque de Quercus perturbado | 2080 |
| Vicente Guerrero | Matorral xerófilo | 1976 | |
| Guanajuato | San José Iturbide | Bosque de Quercus perturbado | 2308 |
| León | Bosque de Quercus | 2570 | |
| Hidalgo | Epazoyucan | Bosque de encino | 2812 |
| San Miguel Regla | Bosque de encino | 2039 | |
| México | Ozumba | Bosque de pino encino | 2365 |
| Tepotzotlán | Matorral | 2753 | |
| Texcoco | Vegetación secundaria | 2334 | |
| Puebla | Coxcatlán | Vegetación secundaria de Pinus patula | 2582 |
| Querétaro | Jalpan de Serra | Bosque de Juniperus | 1526 |
| Pinal de Amoles | Bosque de encino | 2323 | |
| San Luis Potosí | Guadalcázar | Matorral submontano | 1261 |
| Zacatecas | Saín Alto | Vegetación secundaria | 2172 |
| Villa Insurgentes | Bosque de encino perturbado | 2147 |
En el trabajo de campo se consideró la localidad tipo de la especie, registrada en las cercanías de la Ciudad de México (Espejo-Serna & López-Ferrari 1993), también se incluyeron las áreas de distribución naturales cercanas a la misma. Conjuntamente se consideraron las localidades tipo de A. kunthii G. Don. (Guanajuato, entre Santa Cruz de la Sierra y monte El Gigante) y A. longifolium (Kunth) Spreng (Querétaro, creciendo entre Arroyozarco y San Juan del Río), ya que se tiene registro que estas especies son consideradas como sinónimos de A. glandulosum (POWO; TPL; Tropicos; Espejo-Serna & López-Ferrari, 2003, Galván 2010 Ownbey 1947, 1950, Traub 1967, 1968). Finalmente, todos los especímenes recolectados en campo, fueron identificados con base en Traub (1967, 1968), para confirmar que el material recolectado pertenece a la especie bajo estudio.
Análisis morfométrico. Se midieron 20 caracteres cuantitativos de órganos vegetativos y florales (Material suplementario Tabla S1) utilizando una regla metálica convencional, un vernier digital y un microscopio estereoscópico (Carl Zeiss, ZEIZZ, Jena, Alemania), empleando un ocular con reglilla graduada en milímetros. Se determinaron 22 caracteres cualitativos (Material suplementario Tabla S1) de órganos vegetativos y florales. Todos los caracteres se obtuvieron a partir de 40 ejemplares depositados en los herbarios CHAPA, SLPM y UAMIZ (Thiers 2023), así como de 50 ejemplares recolectados en campo.
Análisis estadístico. Se obtuvieron los estadísticos descriptivos de las 20 variables cuantitativas (Material suplementario Tabla S2). A la base de datos resultante, se le realizó una prueba de correlación de Spearman, ya que los conjuntos de datos no cumplieron con el supuesto de normalidad multivariada (Winter et al. 2016). Esta prueba se utilizó para determinar si existían caracteres altamente correlacionados (> 0.60, Altman & Krzywinsky 2015) y eliminarlos de análisis posteriores (Palacio et al. 2020).
Las variables fueron transformadas con logaritmo base 10 con la finalidad de cumplir con el supuesto de normalidad multivariada, posteriormente se empleó un análisis de componentes principales (ACP), para reducir la dimensionalidad del grupo de datos e identificar las variables que contribuyen a explicar la variación morfológica de A. glandulosum (Palacio et al. 2020). A las variables con mayor peso se les aplicó un modelo lineal generalizado con distribución gaussiana (GLM), el cual permite evaluar si existen diferencias significativas entre los ejemplares de las distintas poblaciones. La elección de esta técnica estadística se basó en el hecho de que las variables no cumplieron con los supuestos de la normalidad y homocedasticidad (Bolker et al. 2009).
A través de un análisis discriminante lineal (ADL) se obtuvieron las funciones discriminantes para identificar los caracteres morfológicos vegetativos y florales que contienen la información útil para la diferenciación de las poblaciones. Este análisis se realizó empleando la librería MASS v. 7.3-58.4 (Venables & Ripley 2002). La evaluación del modelo se realizó mediante la matriz de confusión, que permite visualizar la proporción de correctas clasificaciones realizadas por el ADL (Palacio et al. 2020).
Finalmente, se realizó un análisis de conglomerados con el método de Ward (Palacio et al. 2020), empleando una matriz estandarizada con las variables cuantitativas que resultaron con alto valor informativo en el ADL y seis caracteres cualitativos (Tabla S3); el objetivo fue determinar el grado de similitud entre las poblaciones estudiadas. Todos los análisis estadísticos fueron ejecutados en el paquete estadístico R 4.2.1 (R Core Team 2020).
Resultados
Selección de poblaciones representativas. Después de la revisión morfológica, se determinó que los especímenes recolectados en la localidad tipo de A. kunthii fueron identificados como A. glandulosum. Mientras que aquellos recolectados en San Juan del Río, Querétaro, fueron determinados como A. aff. kunthii.
A partir de la revisión de los ejemplares se determinó que poblaciones identificadas como A. glandulosum presentan caracteres morfológicos similares entre sí. En todos los casos se encontró la presencia de bulbos solitarios, estambres connados basalmente, los cuales a su vez se encuentran adnados a los tépalos. Las poblaciones también comparten la presencia de nectarios septales. Así mismo, estos caracteres han sido propuestos por otros autores para determinar a A. glandulosum.
Análisis estadístico. La Tabla S2 muestra la media y la desviación estándar de los 20 caracteres cuantitativos. La prueba de correlación de Spearman reveló que existe correlación entre algunos caracteres analizados (> 0.60, Altman & Krzywinsky 2015) (Figura 1). Sin embargo, no existe una alta correlación entre los caracteres, por tanto, se consideraron todas las variables para el análisis de componentes principales (ACP).

Figura 1 Matriz de correlación de Sperman para 20 caracteres cuantitativos vegetativos y florales. LB = longitud del bulbo; DB = diámetro del bulbo; LCB = longitud del cuello del bulbo; LH = longitud de la hoja; AH = ancho de la hoja; LES = longitud del escapo; DES = diámetro del escapo; LP = longitud del pedicelo; LTE = longitud del tépalo externo; ATE = ancho del tépalo externo; LTI = longitud del tépalo interno; ATI = ancho del tépalo interno; LF = longitud del filamento; LA = longitud de la antera; AA = ancho de la antera; LO = Longitud del ovario; DO = diámetro del ovario; LE = longitud del estilo; NH = número de hojas; NF = número de flores.
El ACP reveló que 14 caracteres concentran la mayor variación en los primeros tres componentes principales con el 51.29 % de la variación acumulada (Tabla 2). El modelo lineal generalizado indicó que existen diferencias significativas en los 14 caracteres cuantitativos resultantes del ACP P < 0.05, por lo que fueron incluidos en el análisis discriminante.
Tabla 2 Coeficientes del análisis de componentes principales de 20 caracteres cuantitativos vegetativos y florales. *Caracteres de mayor contribución. Descripción de acrónimos revisar Tabla S1.
| PC1 | PC2 | PC3 | |
|---|---|---|---|
| Variación | 29.19 | 14.01 | 7.99 |
| Variación acumulada (%) | 29.19 | 43.20 | 51.29 |
| LB | 0.194 | -0.265 | 0.379* |
| DB | 0.217 | -0.217 | 0.308* |
| LCB | 0.241 | -0.016 | 0.222 |
| LH | 0.235 | -0.154 | -0.015 |
| AH | 0.288* | -0.189 | -0.109 |
| LES | 0.327* | -0.166 | 0.004 |
| DES | 0.337* | -0.124 | -0.079 |
| LP | 0.261* | -0.076 | 0.054 |
| LTE | 0.197 | 0.460* | 0.026 |
| ATE | 0.196 | 0.167 | -0.153 |
| LTI | 0.206 | 0.444* | 0.037 |
| ATI | 0.210 | 0.133 | -0.213 |
| LF | 0.155 | 0.340* | 0.378* |
| LA | 0.250* | 0.087 | -0.161 |
| AA | 0.130 | 0.026 | -0.243 |
| LO | 0.230 | 0.042 | -0.333* |
| DO | 0.201 | 0.074 | -0.324* |
| LE | 0.103 | 0.316* | 0.394* |
| NH | -0.027 | -0.058 | -0.095 |
| NF | 0.243 | -0.292* | 0.109 |
El análisis discriminante lineal (ADL) identificó que 75.04 % de la variación total se explica con las tres primeras funciones discriminantes. El ADL permitió identificar las variables que aportan mayor información para la discriminación (Tabla 3). La representación gráfica de las tres primeras funciones favoreció el ordenamiento de grupos con base en el diámetro del escapo, longitud del tépalo interno y longitud del bulbo (Figura 2). La matriz de confusión (Tabla 4) mostró que la precisión de la clasificación resultante del ADL fue de 71.53 %. Los individuos de 12 poblaciones presentaron el mayor porcentaje de clasificaciones correctas > 80 %, mientras que el resto de las poblaciones presentaron valores de clasificación < 80 %. El análisis de conglomerados basado en tres caracteres cuantitativos y seis caracteres cualitativos recuperó cuatro grupos principales (Figura 3).
Tabla 3 Coeficientes del análisis discriminante lineal, para 14 caracteres cuantitativos vegetativos y florales. *Caracteres de mayor contribución. Descripción de acrónimos revisar Tabla S1.
| Can1 | Can2 | Can3 | |
|---|---|---|---|
| Variación (%) | 39.48 | 63.21 | 75.06 |
| Variación acumulada (%) | 39.48 | 23.72 | 11.84 |
| LB | -0.024 | -0.138 | -0.158 |
| DB | -0.076 | -0.008 | -0.150 |
| LES | -0.001 | 0.004 | 0.004 |
| DES | -1.407* | 0.900 | -0.511 |
| LTE | 0.484 | -0.064 | -0.078 |
| ATE | -0.130 | -0.279 | 1.148* |
| LTI | 0.685 | 0.883 | -0.096 |
| ATI | -0.003 | 0.760 | -0.815 |
| LF | 0.320 | 0.131 | -0.446 |
| AA | 1.067* | 2.030* | -1.153* |
| LO | -1.686* | -0.692 | 1.114* |
| DO | -0.390 | 0.088 | 0.077 |
| LE | 0.175 | -0.308 | -0.246 |
| NH | 0.077 | 0.200 | -0.319 |
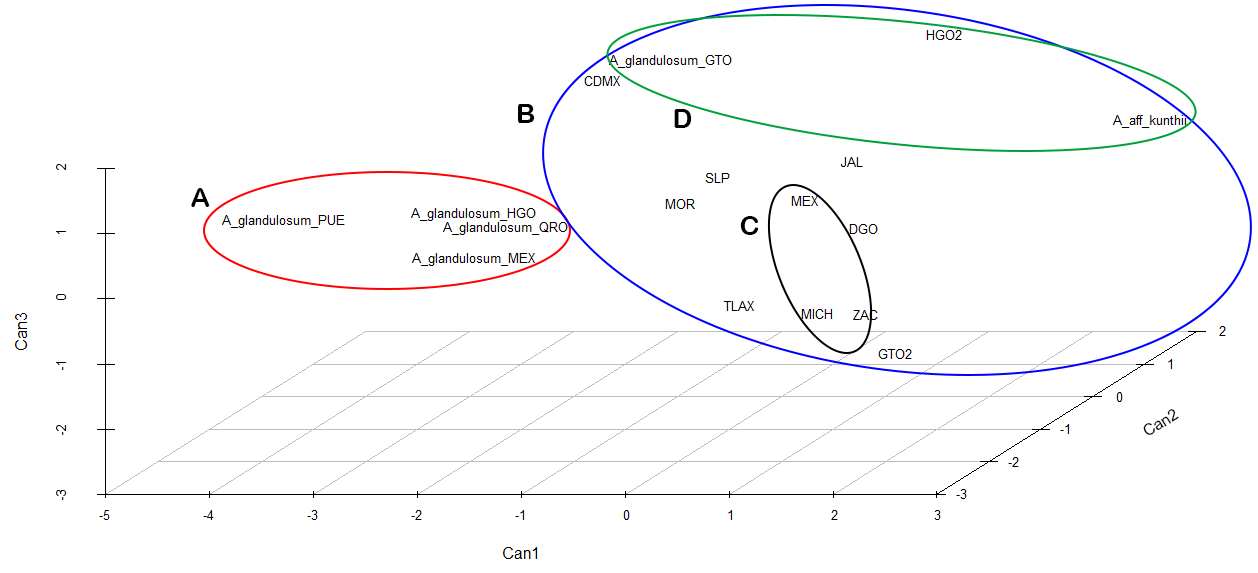
Figura 2 Gráfica del análisis discriminante lineal de poblaciones de Allium glandulosum Link & Otto estudiadas en México. Se grafican los coeficientes para las tres primeras funciones canónicas. Las elipses corresponden a los grupos de poblaciones.
Tabla 4 Porcentaje de individuos clasificados correctamente en su grupo en función del análisis discriminante canónico.
| Poblaciones | % de individuos |
|---|---|
| A_glandulosum_GTO, A_glandulosum_MEX, A_glandulosum_PUE, A_glandulosum_QRO, CDMX, DGO, MEX, MICH, MOR, A. aff kunthii, SLP, ZAC | > 80 |
| A_glandulosum_HGO, GTO, HGO, JAL, TLAX | > 65 - < 80 |

Discusión
Selección de poblaciones representativas. De acuerdo con la literatura, Link y Otto describieron a A. glandulosum a partir de plantas colectadas por Deppe en 1826, cerca de la Ciudad de México (Link & Otto 1828, Espejo-Serna & López-Ferrari 1993). En esa descripción (Link & Otto 1828) y aquellas consideradas por otros autores (Ownbey 1947, 1950, Traub 1967, 1968) esta especie se puede distinguir del resto de las especies de Allium descritas para México por caracteres morfológicos vegetativos y florales. Por ello, se priorizaron poblaciones del Valle de México como aquellas pertenecientes a la localidad tipo o cercanas a la misma, se consideraron a los especímenes recolectados en el municipio de Ozumba, Estado de México como referencia para establecer comparaciones con las demás poblaciones nombradas como A. glandulosum. Estos especímenes miden de 50-75 cm de alto, presentan bulbos solitarios, desarrollando un rizoma blanco > 10 cm de largo, terminando en un bulbilo, escapo con bordes prominentes, umbela de flores rojas con tépalos extendidos y nectarios septales. Los caracteres antes mencionados son consistentes con la descripción e ilustración del tipo de A. glandulosum (Link & Otto 1828). Las poblaciones recolectadas en los estados de Hidalgo, Puebla y Querétaro (Jalpan de Serra y Pinal de Amoles) presentaron similitudes morfológicas con la del Estado de México, también reconocidos como A. glandulosum.
En Allium se ha determinado que caracteres morfológicos como el diámetro del bulbo, ancho, forma y número de hojas, la longitud del escapo, la longitud del pedicelo y la relación bráctea floral/longitud del escapo, así como la forma del estigma y el color de la túnica del bulbo son relevantes para agrupar y distinguir especies (El-Gadi & Elkington 1977, Badr & Elkington 1978). El análisis de componentes principales (ACP) reveló que 14 caracteres cuantitativos (Cuadro 3) son los de mayor aporte al análisis. De los cuales, el diámetro del bulbo y la longitud del escapo coinciden con lo reportado previamente (El-Gadi & Elkington 1977, Badr & Elkington 1978). La longitud del bulbo, diámetro del escapo, longitud del tépalo exterior y del estilo también presentaron variación en otros géneros de Amaryllidaceae (Díaz-Lifante & Andrés-Camacho 2007, El-Hadidy et al. 2012).
La representación gráfica del análisis discriminante lineal (ADL) permitió reconocer los tres caracteres cuantitativos con mayor valor de discriminación para la formación de cuatro grupos. Este resultado es significativo ya que en el grupo A se asocian poblaciones del centro de México que preliminarmente se habían determinado como A. glandulosum, excepto una población recolectada en el estado de Guanajuato. Las poblaciones del grupo A presentan caracteres morfológicos vegetativos y flores que previamente habían sido reportados en la descripción e ilustración tipo, como son: hojas de 30 cm de largo, 2-3 mm de ancho, canaliculadas, planas en la parte superior y aquilladas en la base; escapo de bordes prominentes; pedicelos de 2.5 cm de largo; tépalos de color marrón rojizo oscuro; filamentos connados en la base; ovario con seis surcos y tres glándulas, cada una en un surco alterno del ovario (Link & Otto 1828. Adicionalmente el ADL reveló que los individuos de estas poblaciones se clasificaron correctamente en su respectiva población con un porcentaje de entre 66-99 %. Este resultado sugiere, la existencia de individuos con caracteres intermedios entre las poblaciones analizadas. En otros estudios empleando solamente caracteres morfológicos (Sánchez et al. 2013) o en combinación con caracteres florales (Gutiérrez et al. 2017), se obtuvieron resultados similares de clasificación.
La combinación de caracteres cuantitativos y cualitativos, resultó en la formación de cuatro grupos por su similitud (Figura 3). Nuestro análisis reveló que la ausencia de indumento en las hojas es importante para reconocer las poblaciones del grupo A. En este grupo, las poblaciones de Michoacán y Zacatecas son similares por presentar la mayor longitud de los tépalos ≥ 7.5 mm. Las poblaciones de este grupo se encuentran en el bosque de encino. En la agrupación B se encuentran poblaciones del centro de México que previamente se habían identificado como A. glandulosum, las cuales se diferencian de los demás grupos por presentar un diámetro del escapo > 1.8 mm. Además, son similares por la longitud del escapo > 48.0 cm, longitud del pedicelo ≥ 2.5 cm y la presencia de flores rojas. Ecológicamente estas poblaciones también muestran similitud, ya que se encuentran predominantemente en bosque de encino o pino, en donde los bulbos se encuentran considerablemente cubiertos por suelos húmedos y abundante materia orgánica, facilitando el desarrollo de estructuras de reproducción como los rizomas con bulbilos terminales.
En el grupo C se incluyeron poblaciones con longitud del bulbo ≤ 1.0 cm. Pese a que una población de Guanajuato había sido determinada como A. glandulosum, el análisis de conglomerados indicó que esta población es similar a una población de Hidalgo y con aquellos individuos determinados preliminarmente como A. aff kunthii por la longitud de la hoja < 23.0 cm y el diámetro del escapo < 1.2 mm. Estas poblaciones se encontraron en áreas abiertas y a diferencia de las anteriores, los bulbos se encontraron situados a escasos centímetros del suelo con muy poca o nula presencia de materia orgánica. En el grupo D se incluyeron individuos de poblaciones que se distribuyen en el centro y occidente de México, en diversos tipos de vegetación, desde bosques de encino hasta vegetación secundaria. Especímenes de este grupo se caracteriza por presentar una longitud del bulbo > 1.1 cm. Además, las proporciones de la longitud de la hoja y el diámetro del escapo son mayores que en el resto de los grupos.
El análisis de conglomerados indicó que A. aff kunthii difiere morfológicamente de las poblaciones de A. glandulosum (Figura 3), que se tomaron como referencia para establecer las comparaciones con el resto de las poblaciones analizadas. Sería pertinente reconsiderar la sinonimia de A. kunthii en A. glandulosum. Sin embargo, son necesarios más estudios con otro enfoque que apoyen esta propuesta.
En el pasado Link y Otto (1828) describieron a A. glandulosum a partir de material procedente a las cercanías de la Cuidad de México, y las flores de la colecta original eran de color rojo intenso. Ownbey (1947, 1950) menciona que esta forma todavía se recolecta con frecuencia en el centro del país. Además, señala que hacia el norte de México las flores son blancas, con el color confinado principalmente en la nervadura central e indica que este patrón de coloración de las flores se encuentra dentro del rango de distribución de la especie y considera que ambas formas pertenecen a una sola entidad, que debe tomar el nombre de A. glandulosum. Los resultados obtenidos coinciden con lo mencionado por Ownbey (1947, 1950), ya que individuos de flores rojas se recolectaron con frecuencia a lo largo de la Faja Volcánica Transmexicana.
Los análisis estadísticos permitieron determinar que A. glandulosum presenta una amplia variación morfológica en México, tanto en estructuras vegetativas como en las reproductivas. Sin embargo, existen caracteres como altura de los individuos, morfología del bulbo, escapo y flores que permiten reconocer poblaciones de esta especie. Futuros estudios, permitirán determinar el efecto de factores ambientales y geográficos sobre dicha variación. Ampliar y combinar este estudio con análisis anatómicos y moleculares, brindarán una visión más completa de la amplia variación de esta especie a lo largo de México. Por el momento y con el objetivo de abordar la variación de morfológica de A. glandulosum, los caracteres que permiten una discriminación entre los grupos de poblaciones pueden considerarse eventualmente útiles en análisis filogenéticos.
Material suplementario
El material suplementario puede ser consultado aquí https://doi.org/10.17129/botsci.3403

