











 text new page (beta)
text new page (beta) English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO 
 Permalink
PermalinkThe crab genus Chorilia Dana, 1851, is made up of 3 species, all previously considered subspecies of Chorilia longipes Dana, 1851 (Garth, 1958; Ng, Guinot, & Davie, 2008). The nominal species, C. longipes is found along the west coast of North America, while C. japonica (Miers, 1879) is known from the western Pacific, Japan and Korea. The third species, C. turgida Rathbun, 1924, was described as C. longipes turgida based on a male of 50 mm carapace length (CL) collected off San Diego, California, USA, in 359 fathoms (ca 665 m).
During sampling operations off the west coast of the Baja California Peninsula, Mexico, aboard the R/V "El Puma" of the Universidad Nacional Autónoma de México (UNAM), a small series of majoid crabs belonging to Chorilia were collected with a benthic sledge. This material represents new records and a considerable distribution range extension of this species south of the USA-Mexico border. All the specimens are kept in the Regional Collection of Marine Invertebrates (EMU) of UNAM, in Mazatlán, Mexico.
Chorilia turgida Rathbun, 1924
Material examined . TALUD XVI-B. St. 17 (29°57'18" N, 116°1'30" W), May 29, 2014, 1 M (CW, 15.3 mm), 734-774 m (EMU-10525). St. 18 (30°39'18" N, 116°25'54" W), May 25, 2014, 740-785 m, 2 M (CL 20.5-23.1 mm) (EMU-10526), 7 M (CW, 9.8-16.5 mm) and 6F (CW, 13.4-21.8 mm) (EMU-10527). 6 M (CW, 13.9-30.8 mm) and 8F (CW, 13.9-28.1 mm) (EMU-10528), 1 M (CW, 25.7 mm) and 2 F (CW, 23.7-25.6 mm) (EMU-10635). St. 19 (30°38' N, 116°31'40" W), May 25, 2014, 1F (CW 13.4 mm), 1385-1,433 m (EMU-10529).
Remarks . The only specimens recorded for the Pacific coast of Mexico are from the vicinity of the Cortés Bank (a broken specimen and a young male), close to the USA border (Garth, 1958: table 54). The material examined fit well with the characters provided by Garth (1958) to separate turgida from longipes , including the more inflated branchial region, the more spinous carapace, the sharp hepatic spine, and the more diverging rostral spines (Fig. 1). Garth (1958) considered both forms as almost segregated along a latitudinal gradient, longipes representing the northern form and turgida the southern one. Garth (1958) further noted that distributions of both "sub-species" overlap in the southern portion of their range, and that the presence of the typical form, i.e., longipes , a species associated with cooler (northern) water than turgida , might be related to the presence of upwellings in the south. Wicksten (2012) considered only one species in her list, but noted that two sub-species are considered along the west coast of the USA, from Alaska to California: C. longipes longipes from Alaska to Santa Catalina Island, and C. l. turgida , from Monterey Bay to off San Diego.

Figure 1 Chorilia turgida Rathbun, 1924: A, fresh specimen, male, CL 30.8mm, dorsal view (EMU-10528); B, same, preserved, detail of hepatic and branchial regions.
The material examined represents new records for C. turgida from the west coast of Mexico, where it was previously reported from the Cortés Bank (Garth, 1958) (32°25'30.00" N, 119°03'30.00" W), and a distribution range extension to 29°57'18" N, 116°01'30" W (Fig. 2). It was collected in a depth range of 734-1,433 m. Wicksten (2012) gave a depth range for C. longipes of 33-1,200 m noting that it is usually found in deeper water in the southern portion of its distribution range. Garth (1958) provided a list of precise records for both "forms": from 10 to 298 fathoms (ca 18-550 m depth) for C. "longipes "longipes (the typical form), and from 32 to 650 fathoms (ca 60-1,200 m depth) for C. "longipes " turgida . Present records therefore increase the deeper distribution limit of C. turgida and reinforce the idea that this species occurs in deeper water in its southern range (i.e., to 1,433 m depth). Other parameters measured at the collection sites from western Mexico are the epibenthic temperature (3.16-5.48 °C) and dissolved oxygen concentration (0.25-0.90 ml/l O2), which indicate the association of C. turgida with cold water and mild to severe hypoxic conditions, at depths which correspond to the lower boundary of the Oxygen Minimum Zone (OMZ) that occurs in the area (Helly & Levin, 2004).
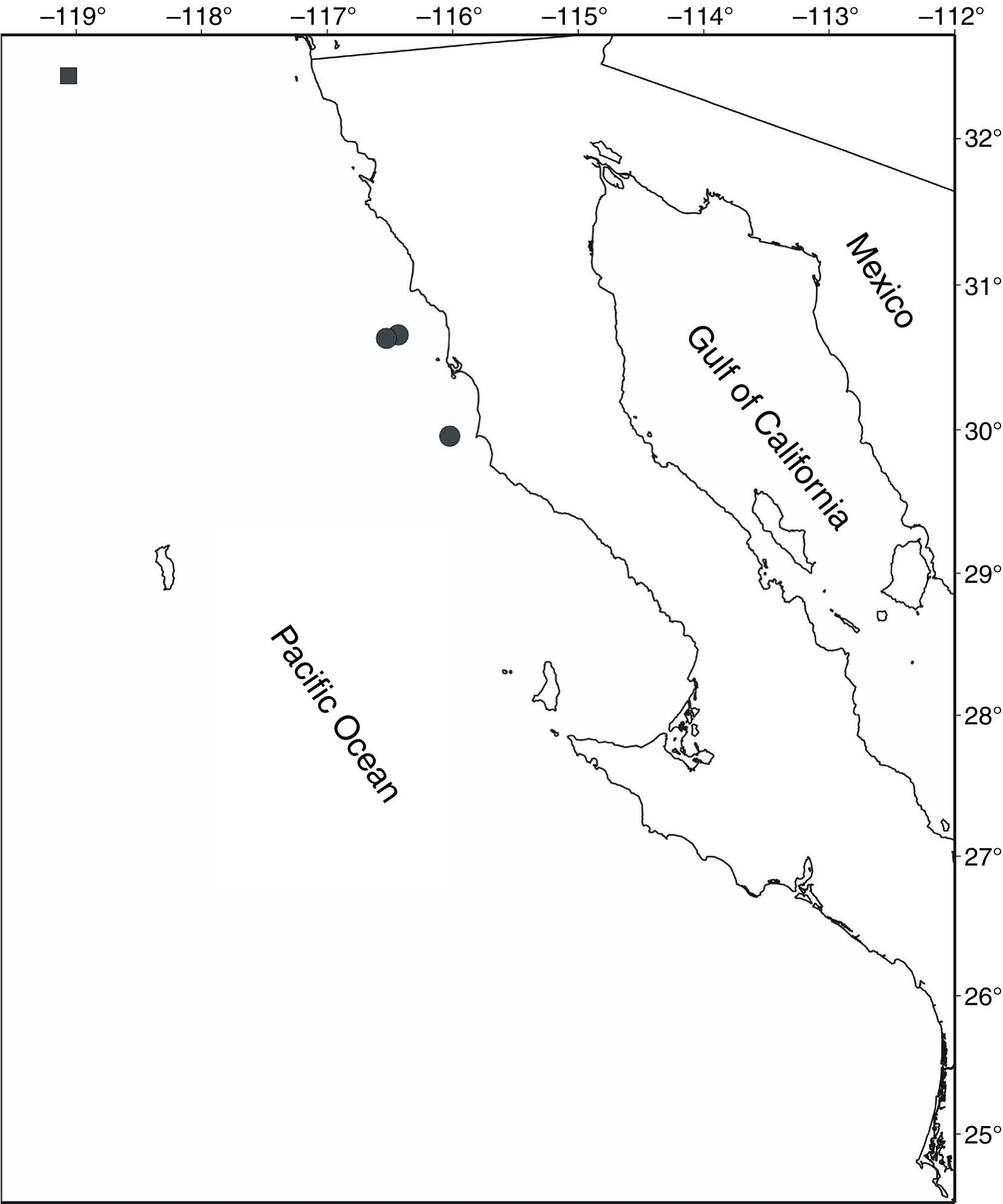
Figure 2 Distribution of Chorilia turgida Rathbun, 1924 off the west coast of Baja California, Mexico. (■) Record of Garth (1958); (●) present study records.
Wicksten (2012) noted that there is a need to analyze morphologic variations within the "longipes " complex considering the water temperature and the geographic location of the specimens available, with a view to determine whether reproductively isolated populations exist or if variations between the two "forms" correspond to ecophenotypes. With the new records of C. turgida under the OMZ, and considering that the OMZ core is particularly wide between 30° N and 60° N (>200 m) and features intermediate hypoxic values <0.5 ml/l but generally >0.2 ml/l (Helly & Levin, 2004), the question arises as to whether this species occurs both above (from about 500 to 600 m and shallower) and below the OMZ (from about 1,100 to 1,200 m depth and below) (Helly & Levin, 2004), and might eventually tolerate very low oxygen concentrations in intermediate depths (1,100 m > depth > 500 m). If the case, this species could have a continuous bathymetric distribution and the OMZ core would not represent a physiological barrier to vertical dispersion, as it occurs in other regions of the eastern Pacific (Hendrickx & Serrano, 2013). Records of C. turgida from off California were mostly in the depth range of 178-650 fathoms (ca 330-1,200 m) (Garth, 1958). This depth range includes the upper boundary of the OMZ core (using 0.5 ml/l as the upper and lower boundaries of dissolved oxygen concentrations), the OMZ core itself, and another area just below the lower boundary of the OMZ (Helly & Levin, 2004), thus supporting the idea of a continuous bathymetric distribution across the OMZ core.
Color . From a freshly collected specimen (Fig. 1A). Carapace pinkish-white, spines salmon. Rostral horns with band of light salmon in proximal half and tip. Chelipeds salmon, with some areas pinkish-white, fingers white with flush of salmon. Ambulatory legs with irregular patches of light salmon, proximal segments and dactylus whitish. The color roughly corresponds to the description by Garth (1958: 266): "usually white, but occasionally bright flesh pink".
Ship time (TALUD XVI-B) was provided by the Coordinación de la Investigación Científica, UNAM, and partly supported by Conacyt (project 179467). The TALUD project received support for laboratory activities from Conacyt (project 179467). The author thanks all scientists, students and crew members who took an active part in the TALUD XVI-B cruise. Photographs of Fig. 1 were taken by José Salgado-Barragán. Mercedes Cordero Ruiz is acknowledged for editing the final version of the manuscript and preparing fig. 1, and Ana K. Barragán Zepeda (Conacyt project 179467 assistant) for her laboratory assistance. I acknowledge the use in figure 1 of Maptool, a product of SEATURTLE.ORG (www.seaturtle.org).

