










Services on Demand
Journal
Article
 text in
text in  English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkRevista mexicana de ciencias agrícolas
Print version ISSN 2007-0934
Rev. Mex. Cienc. Agríc vol.9 spe 20 Texcoco Apr./May. 2018
https://doi.org/10.29312/remexca.v0i20.1004
Articles
Rhizobacteria and arbuscular mycorrhizal fungi associated with poblano chili in the Sierra Nevada of Puebla, Mexico
1Faculty of Agriculture and Zootechnics. Juárez University of the state of Durango. Highway Gómez Palacio-Tlahualilo km 28, Ejido Venecia, Gómez Palacio, Durango, Mexico.
2Postgraduate College-Campus Montecillo, Mexico-Texcoco Highway km 36.5, Montecillo, Texcoco, State of Mexico. CP. 56230. (jalmaraz@colpos.mx; ronaldfc@colpos.mx; trinidad@colpos.mx; pilarrg@colpos.mx).
3Postgraduate College-Campus Puebla. Boulevard Forjadores de Puebla num. 205. Santiago Momoxpan, San Pedro Cholula, Puebla, Mexico. CP. 72760. (toswaldo@colpos.mx).
In the lower parts of the Sierra Nevada, Puebla, Mexico, small farmers grow one of the most traditional chilis in Mexican cuisine, poblano chili. The variability in agricultural practices and the characteristics of the soil lead to various microenvironments of production, causing a variation in the populations of beneficial microorganisms. The objective of the present investigation was to quantify the microbial populations [total fungi (HT), total bacteria (BT), phosphate solubilizing bacteria (BSP), nitrogen-fixing bacteria (BFN), auxin-producing bacteria (BPA) and mycorrhizal fungi arbuscular (HMA)] and fruit production in plantations of poblano chili grown in the open field. By means of serial dilutions and seeding in specific media, HT, BT, BSP, BFN and BPA were quantified. The colonization by HMA was determined by the method of thinning and staining with trypan blue. The highest populations of BT and BPA were found in site seven (Huejotzingo), soil with high phosphorus content (428.8 mg kg-1), HT, BSP and BFN were better in site one (San Matias Tlalancaleca), this site did not present the best soil characteristics, but higher altitude, which correlated positively with these microorganisms. Mycorrhizal colonization was better at site nine (Huejotzingo), in a soil with low phosphorus content (44.3 mg kg-1) and moderately alkaline pH (7.6). The yield of fruits was higher in site 8 (Huejotzingo), which presented higher content of organic matter (1.48%) and total nitrogen (0.07%) in soil.
Keywords: Capsicum annuum; nitrogen fixing bacteria; phosphate solubilizing bacteria; auxin producing bacteria
En las partes bajas de la Sierra Nevada, Puebla, México, pequeños agricultores cultivan chiles tradicionales en la cocina mexicana, el chile poblano. La variabilidad en las prácticas agrícolas y las características del suelo, conducen a diversos microambientes de producción, originando una variación de los microorganismos benéficos. El objetivo de la presente investigación fue cuantificar las poblaciones microbianas [hongos totales (HT), bacterias totales (BT), bacterias solubilizadoras de fosfato (BSP), bacterias fijadoras de nitrógeno (BFN), bacterias productoras de auxinas (BPA) y hongos micorrízicos arbusculares (HMA)] y la producción de frutos en plantaciones de chile poblano cultivado en campo abierto. Mediante diluciones seriadas y siembra en medios específicos se cuantificaron HT, BT, BSP, BFN y BPA. La colonización por HMA se determinó por el método de clareo y tinción con azul tripano. Las poblaciones más altas de BT y BPA fueron encontradas en el sitio siete (Huejotzingo), suelo con alto contenido de fósforo (428.8 mg kg-1), los HT, BSP y BFN fueron mejores en el sitio uno (San Matías Tlalancaleca), éste sitio no tiene las mejores características del suelo, pero si mayor altitud, que se correlacionó positivamente con estos microorganismos. La colonización micorrízica fue mejor en el sitio nueve (Huejotzingo), suelo con bajo contenido en fósforo (44.3 mg kg-1) y pH moderadamente alcalino (7.6). El rendimiento de frutos fue más alto en el sitio 8 (Huejotzingo), que presentó mayor contenido de materia orgánica (1.48%) y nitrógeno total (0.07%) en suelo.
Palabras clave: Capsicum annuum; bacterias fijadoras de nitrógeno; bacterias solubilizadoras de fosfatos; bacterias productoras de auxinas
Introduction
The poblano chili in the state of Puebla represents an ancient cultural tradition due to its gastronomic, economic and social importance, as it is associated with dishes such as “chiles en nogada” and “mole poblano” (Cyphers et al., 2009). It is produced mainly in the Sierra Nevada, in municipalities such as San Matias Tlalancaleca, San Lorenzo Chiahutzingo, San Rafael Tlanalapa, Moyozingo, Huejotzingo and San Lucas (Huerta et al., 2007). In 2007, the state of Puebla presented an area cultivated with Poblano chili of 600 ha and a production of 4 800 t (SDR Puebla, 2007), in this respect Rodríguez et al. (2007) mention that the production of poblano chili has decreased from 25 t ha-1 to less than 10 t ha-1 in the last 10 years.
The decrease in the production of poblano chili is associated to several factors, and highlight the "drying" and "drowning" caused by Phytophthora capsici, Fusarium spp., Rhizoctonia solani, Pythium spp., Alternaria spp. the wilt caused by the nematode Nacobbus aberrans, causing losses of up to 100% of the crop, either in the greenhouse, in the field or in storage (Rodriguez et al., 2007, Bautista-Calles et al., 2010). In addition, factors such as viral diseases and physical damage to crops caused by insects (aphids, mites and thrips) are added (Huerta et al., 2007), as well as the availability of nutrients and water (Mena-Violante et al., 2006). According to Huerta et al. (2007) the poblano chili of the Sierra Nevada is better suited to soils with sandy-silty texture than in clay soils, so excess moisture should be avoided.
According to the FAO classification, the soils that predominate in the study area are regosols, cambisols, feozems and fluvisols (INEGI, 2017), unlike the first two, the soils of the feozem and fluvisol types have high and moderate levels of organic matter (FAO, 2006, Galicia et al., 2015). High levels of organic matter induce greater soil fertility, due to high nutrient contents and organic carbon; The latter, as an energy source, allows a greater diversity of bacteria, fungi, nematodes, earthworms, insects and arthropods that interact synergistically in the ecosystem (Mader et al., 2002; Oehl et al., 2002). But when organic matter is low, there is low organic carbon, low fertility and greater attack by phytopathogens when using crops as their source of food, according to Ghorbani et al. (2010), nitrogen fertilization in soils with a low amount of organic matter, favors the development of pathogens such as Rhizoctonia sp., Fusarium sp., Sclerotium sp., etc.
The soil is the physical substrate for the life of terrestrial animals (including humans) and a means for the growth of plants, dampens water flows, decomposes and releases nutrients and allows the regulation of emissions of greenhouse gases (Stott and Taylor, 2016). The microorganisms that live in the soil are intimately associated with all these functions (Aislabie and Deslippe, 2013). The microbiotic are found in higher density near the roots of plants, where there is greater availability of exudate organic compounds (Massenssini et al., 2014), the quality of these exudates allows the recruitment of certain microorganisms that will have a positive, neutral effect or negative on the plant, potentially determining the composition of plant communities (Wolfe and Klironomos, 2005).
The most studied groups of organisms are plant growth promoting rhizobacteria (RPCV) and arbuscular mycorrhizal fungi (HMA), due to the benefits they provide to plants. The RPCV provide nutrients to plants by fixing and solubilizing elements such as atmospheric nitrogen, phosphorus and potassium; inhibit the development of phytopathogens and synthesize growth regulators such as auxins, gibberellins and cytokinins (Hariprasad and Niranjana 2009, Sandhya et al., 2010). The HMA favor the absorption of water and nutrients, and increase the tolerance to stress caused by biotic and abiotic factors (Perner et al., 2007, Sawers et al., 2008). The objective of the research was to quantify the populations of RPCV and HMA in plantations of poblano chili cultivated in the open field and their possible effects on crop yield.
Materials and methods
In September 2012, rhizospheric chili poblano soil was collected in nine sites of the Sierra Nevada region in the state of Puebla, Mexico. The sampling included poblano chili fields from three municipalities: San Matias Tlalancaleca with three sites, whose altitudes were from 2 414 to 2 467 meters above sea level, San Lorenzo Chiautzingo with two sites at altitudes of 2 404 and 2 425 meters above sea level, and Huejotzingo with four sites of altitudes ranging from 2 284 to 2 313 meters above sea level (Table 1 and Figure 1).
Table 1 Soil characteristics and location of study sites in the Sierra Nevada region of Puebla, Mexico.
| Sites | Localization | Altitude (m | pH | MO | NT | P | K | Textural |
| San Matías Tlalancaleca | ||||||||
| 1 | 19°22.313’ N y 98°32.032’ W | 2467 | 5.5 | 0.27 | 0.01 | 184.7 | 412 | FA |
| 2 | 19°22.257’ N y 98°31.924’ W | 2453 | 5.8 | 0.54 | 0.03 | 224 | 324 | FAA |
| 3 | 19°20.665’ N y 98°29.209’ W | 2414 | 6.8 | 1.48 | 0.07 | 314.5 | 854 | FA |
| San Lorenzo Chiautzingo | ||||||||
| 4 | 19°13.440’ N y 98°28.903’ W | 2425 | 6.9 | 0.94 | 0.05 | 153.7 | 546 | FA |
| 5 | 19°13.052’ N y 98°28.261’ W | 2404 | 4.9 | 0.27 | 0.01 | 187.1 | 242 | FA |
| Huejotzingo | ||||||||
| 6 | 19°08.504’ N y 98°24.700’ W | 2313 | 5.8 | 0.4 | 0.02 | 41.5 | 266 | AF |
| 7 | 19°08.968’ N y 98°24.430’ W | 2292 | 6.7 | 0.94 | 0.05 | 428.8 | 508 | AF |
| 8 | 19°11.985’ N y 98°25.711’ W | 2286 | 7.3 | 1.48 | 0.07 | 50.1 | 430 | AF |
| 9 | 19°12.295’ N y 98°25.510’ W | 2284 | 7.6 | 0.67 | 0.03 | 44.3 | 410 | AF |
MO= organic matter (%); NT= total nitrogen (%); P= phosphorus (mg kg-1); K= potassium (mg kg-1); FA= sandy loam; FAA= loam clay sandy; AF= loamy sand.
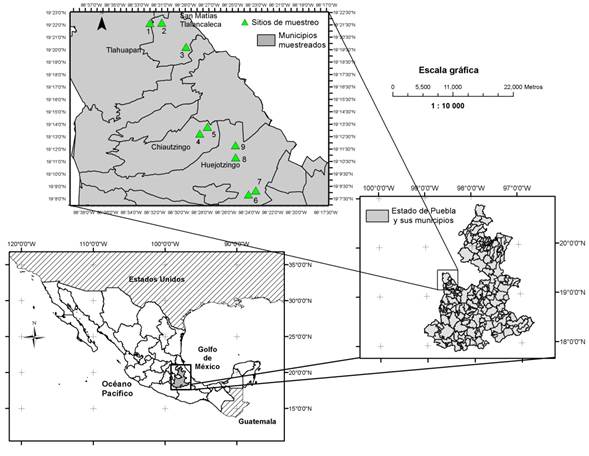
Figure 1 Geographic location of the nine sites where rhizospheric soil samples of poblano chili were collected in the Sierra Nevada, Puebla State, Mexico.
Four samples were collected per site, which included soil and plants of poblano chili selected visually for their best appearance (health and vigor), they were kept in a cooler for their transfer to the microbiology laboratory of the Montecillo Campus of the Postgraduate School, Montecillo, State from Mexico. In the sampling sites the pH values (soil, water 1:2) fluctuated between 4.9 to 7.6. The organic matter (MO) values were from 0.27 to 1.48%, the total nitrogen (NT) from 0.01 to .07%, the phosphorus (P) from 41.1 to 428.8 mg kg-1 and of potassium (K) from 242 to 854 mg kg-1. Soil texture was classified as sandy loam at sites one, three, four and five, sandy loam for site two and loamy sand for sites six, seven, eight and nine (Table 1).
The collected samples were processed in a laminar flow hood under aseptic conditions. The 10 g of sample was weighed and placed in bottles with 90 mL of sterile distilled water (first dilution) and from it serial decimals dilutions were made (up to 10-4). In Petri dishes, 0.1 mL of each dilution was seeded and the aliquot was distributed on the surface of the solid medium with the help of an L-shaped glass rod. Specific culture media were used: nutritive agar for total bacteria (BT), Pikovskaya, to detect phosphate solubilizing bacteria (BSP), Rennie, for nitrogen fixers (BFN), Luria-Bertani (LB), for auxin-producing bacteria (BPA) and PDA medium (potato dextrose agar) for total fungi. The colonies were quantified after three days of incubation at 28 °C.
The number of fruits was quantified in poblano chili plants at the time of collection in the field, the roots of the plants collected from poblano chili were evaluated for colonization by HMA following the method of thinning and staining with trypan blue (Phillips and Hayman, 1970), total colonization (PCT) was estimated in roots of 1 cm in length expressed in percentages (Biermann and Linderman, 1981). A part of the rhizospheric soil was used for the extraction and quantification of HMA spores by means of the wet sieving and decanting technique (Gerdemann and Nicolson, 1963), expressed in 100 g of dry soil (100 g). The data obtained were analyzed with the SAS statistical package for Windows (SAS Institute Inc. 2002), performing an analysis of variance and means comparison test (Tukey, α= 0.05).
Results and discussion
The populations of microorganisms found at each site were statistically different (Tukey α= 0.05) (Table 2). Site seven, whose sample was collected in the municipality of Huejotzingo, presented the highest populations of BT (53 x 105 UFC g-1 soil) and BPA (92 x 104 UFC g-1 soil), while the site one of San Matias Tlalancaleca had the highest populations of total fungi (82 x 102 UFC g-1 soil), BSP (46 x 104 UFC g-1 soil) and BFN (36 x 104 UFC g-1 soil). The lowest populations were found in sites one for BT (10 x 105 UFC g-1 soil), nine for HT (3 x 102 UFC g-1 soil), three for BSP (1 x 104 UFC g-1 soil), five and eight for BPA (4 x 104 UFC g-1 soil) and five and six for BFN (10 x 104 UFC g-1 soil) (Table 2).
Table 2 Colony forming units (UFC) quantified in rhizospheric soil of poblano chili collected in nine sites of the Sierra Nevada de Puebla, Mexico.
| Sites | BT | BFN | BSP | BPA | HT | ||
| 105 UFC g-1 suelo | 104 UFC g-1 soil | 102 UFC g-1 soil | |||||
| San Matías Tlalancaleca | |||||||
| 1 | 10.3 ±2.2a | 36 ±13.2a | 46 ±13.7a | 7 ±1.7bc | 82 ±33.2a | ||
| 2 | 11.2 ±3.8a | 18 ±6.2ab | 36 ±22.5ab | 6 ±1.1bc | 56 ±12.8ab | ||
| 3 | 16 ±5.3a | 11 ±1.5b | 1 ±0.05c | 22 ±2.8b | 26 ±2.3bc | ||
| San Lorenzo Chiautzingo | |||||||
| 4 | 26.1 ±1.6a | 12 ±2ab | 8 ±1.5ab | 10 ±2.2bc | 7 ±2.5c | ||
| 5 | 20.4 ±3.7a | 10 ±1.3b | 20 ±9.5ab | 4 ±0.8c | 10 ±2.5c | ||
| Huejotzingo | |||||||
| 6 | 27 ±6.7a | 10 ±1.3b | 26 ±8.6ab | 5 ±1.8c | 7 ±2.7c | ||
| 7 | 53.2 ±27.4a | 15 ±2.7ab | 6 ±2.4b | 92 ±42a | 19 ±4.9bc | ||
| 8 | 14.5 ±1.2a | 11 ±1.2b | 9 ±1.3ab | 4 ±0.8c | 6 ±0.9c | ||
| 9 | 16.4 ±2a | 14 ±1ab | 9 ±0.7ab | 11 ±1.9bc | 3 ±0.7c | ||
BT= total bacteria; HT= total fungi; BSP= phosphate solubilizing bacteria; BPA= auxin-producing bacteria and BFN= nitrogen-fixing bacteria. Different letters within the same column present significant statistical differences (Tukey, α= 0.05, a> b). Means n= 8, ± standard error.
The densities of HT, BFN and BSP showed positive and statistically significant correlation (α= 0.05) with the altitude of the different sites studied (Table 3), which indicates that the densities of these rhizobacteria were increased in the higher altitude sites. In addition, altitude was negatively correlated with pH, organic matter and nitrogen content, and positively with the phosphorus content in soil, factors that determined the high densities of microorganisms at sites seven and one. The characteristics of the soil influenced the microbial populations evaluated, due to the fact that a negative and significant correlation (α= 0.05) was observed between the HT population and the pH of the soil; negative and significant correlation (α= 0.01) between BSP and pH, MO, NT and K and positive and significant correlation (α= 0.01) with BPA and P content (Table 3).
Table 3 Correlations between the number of microorganisms and soil characteristics in poblano chili collected in nine sites of the Sierra Nevada de Puebla, Mexico.
| Altitude | pH | MO | NT | P | K | Fruits | |
| BT | -0.191 | 0.047 | 0.025 | 0.059 | 0.231 | 0.028 | 0.016 |
| HT | 0.47** | -0.289* | -0.212 | -0.206 | 0.22 | 0.007 | 0.108 |
| BFN | 0.248* | -0.134 | -0.196 | -0.202 | 0.067 | -0.031 | 0.109 |
| BSP | 0.238* | -0.337** | -0.36** | -0.356** | -0.093 | -0.298* | -0.086 |
| BPA | -0.2 | 0.134 | 0.131 | 0.168 | 0.457** | 0.181 | 0.102 |
| CT | -0.693** | 0.538** | 0.066 | -0.074 | -0.246* | -0.039 | 0.248* |
| V | -0.157 | -0.163 | -0.186 | -0.149 | 0.004 | -0.196 | -0.136 |
| E | -0.325** | -0.077 | -0.18 | -0.116 | 0.364** | -0.266* | -0.455** |
| Altitude | 1 | -0.55** | -0.274* | -0.258* | 0.255* | 0.126 | -0.142 |
| Fruits | -0.142 | 0.65** | 0.56** | 0.561** | -0.11 | 0.46** | 1 |
BT= total bacteria; HT= total fungi; BFN= nitrogen-fixing bacteria; BSP= phosphate solubilizing bacteria; BPA= auxin-producing bacteria; CT= total colonization; V= vesicles; E= spores; MO= organic matter (%); NT= total nitrogen (%); P= phosphorus (mg kg-1); K= potassium (mg kg-1). *Significant correlation α= 0.05 and **α = 0.01.
There are few studies on altitude and their influence on the distribution of soil microorganisms are contradictory, and have been related to factors of climate, soil, vegetation and biotic factors (Hofmann et al., 2016). For example, the diversity of Phylum acidobacteria decreases at higher altitudes and is attributed mainly to soil pH (Bryant et al., 2008). The bacterial abundance found by Hofmann et al. (2016) decreased with altitude, and attributed to the influence that altitude has on the temperature and soil organic matter. Weyens et al. (2009), mention that soil microorganisms are influenced by characteristics such as humidity, texture, pH, organic matter, nutritional content, temperature and salinity; they can also be influenced by factors such as the plant species, age and nutritional status of the plant (Adeboye et al., 2006).
Soil ecosystems are highly complex due to the great diversity of microbial species that they harbor, which can be harmful or beneficial for plants. The beneficial microorganisms, such as those evaluated in this study, are classified as RPCV due to the functions they perform, for example the biological fixation of N2 (BFN) and the solubilization of phosphorus (BSP) to make them available to plants in the form of ammonium and inorganic phosphates (Herridge et al., 2008, Restrepo-Franco et al., 2015); In addition, they promote the production of auxins (BPA) that directly influence the development of plants through cell elongation and division, tissue differentiation and apical dominance (Duca et al., 2014). Additionally, RPCV allow the biological control of pathogens (Sandhya et al., 2010), the degradation of organic matter, the adaptation of plants to contaminated soils, drought conditions and extreme pH values (Saraf et al., 2011) and the synthesis of growth regulators such as ethylene, auxins and gibberellins (Kim et al., 2010).
The roots of the chili plants form symbiotic associations with the arbuscular mycorrhizal fungi (Davies Jr et al., 1992), which is confirmed in this study, since the poblano chili was colonized by the HMA between 5 and 68%, existing significant statistical differences between the nine sites sampled (Tukey α= 0.05). The highest colonization (68%) was located in site nine, which corresponds to the collection of soil made in the municipality of Huejotzingo, the lowest value was found in site five, municipality of San Lorenzo Chiautzingo (Table 4). The high percentage of colonization of site nine, possibly related to the low content of P (44.3 mg kg-1) found at the site, as opposed to site five which was much higher (187.1 mg kg-1). According to Javaid (2009), HMA efficiently colonize plants when soils have low phosphorus content.
Table 4 Colonization by HMA in poblano chili plants collected in nine sites of the Sierra Nevada de Puebla, Mexico.
| Sites | Total colonization (%) | Vesicles (%) | Spores 100 g ss | Fruits plant-1 |
| San Matías Tlalancaleca | ||||
| 1 | 8 ±3cd | 1 ±0.5b | 195 ±12d | 6 ±0.3ab |
| 2 | 12 ±4cd | 1.5 ±0.9ab | 477 ±55bc | 5 ±0.6ab |
| 3 | 16 ±7cd | 0.3 ±0.3b | 235 ±60d | 6 ±0.9ab |
| San Lorenzo Chiautzingo | ||||
| 4 | 23 ±12bcd | 0.7 ±0.7b | 486 ±60bc | 5 ±0.4ab |
| 5 | 5 ±2d | 0.3 ±0.3b | 654 ±107b | 0 ±0c |
| Huejotzingo | ||||
| 6 | 44 ±2b | 5.2 ±3.1a | 447 ±34c | 4 ±0.8b |
| 7 | 43 ±5b | 2.8 ±1.5ab | 896 ±98a | 4 ±0.8b |
| 8 | 27 ±7bc | 0.4 ±0.4b | 355 ±24cd | 7 ±0.9a |
| 9 | 68 ±10a | 0 ±0c | 483 ±71bc | 5 ±0.8ab |
Different letters within the same column, present significant statistical differences. Means n= 4 ± standard error (Tukey, α= 0.05, a> b).
The presence of vesicles was relatively low, being the site six with the highest presence of these mycorrhizal structures with 5.2%, while site nine did not present vesicles. The highest number of spores in soil was found in site seven with 896 spores in 100 g, while in site one the smallest amount was found with 195 spores. The CT showed positive and significant correlation (α= 0.01) with the pH and the number of spores with the P of the soil (Table 3), while the altitude influenced the CT and the spores present of HMA when presenting negative and significant correlation (α= 0.01), indicating that at higher altitude there is less mycorrhizal colonization. The P presented negative correlation with the CT and positive with the number of spores, which indicates that the lower the P content, the higher the CT and as the P increases, the number of spores will increase.
Factors such as the altitudinal gradient, physical (texture, structure, porosity) and chemical characteristics of the soil (pH, organic matter, nutrients) (Coutinho et al., 2015) and human activities (Dumbrell et al., 2010) influence the diversity, colonization and number of spores present of HMA. The management of crops with fertilizers or herbicides affects the mycorrhizal symbiosis (Pasaribu et al., 2011), causing a decrease in the diversity and abundance of spores (Oehl et al., 2004). In the region where poblano chili is grown in the state of Puebla, no herbicides are used, so the variation in microbial populations is due to other factors.
The HMA-plant symbiosis is estimated to occur between 70-90% of terrestrial plant species, in agricultural ecosystems, as in this study, or in natural ecosystems (Smith and Reed, 2008; Zhu et al., 2010). The HMA are obligate symbionts and acquire carbon from their host plants to complete their life cycle (Bago et al., 2000); in return, the fungus provides several benefits for the plant, for example, increased absorption of nutrients mainly P, N and K (Perner et al., 2007), Zn, Cu, Fe, S, Ca (Allen, 2009) , Mg and B (Subramanian et al., 2006; Altomare and Tringovska, 2011) and tolerance to stress caused by biotic and abiotic factors (Sawers et al., 2008).
The quantified amount of fruits per plant presented significant statistical differences (Tukey α= 0.05), being the best site eight with seven fruits produced by plants on average, followed by sites one and three with six fruits, respectively. Site five did not present healthy fruits due to the presence of fungal diseases in the crop, which did not allow the phenological stage of fruiting and fruit development to be reached. The CT by HMA presented positive and significant correlation (α= 0.05) with the production of fruits per plant, indicative that the higher the CT, the higher the fruit yield; however, the soil characteristics also influenced this variable, since the plants with the highest fruit produced were found in those sites with better characteristics of pH, MO, NT and K, obtaining a significant correlation (α= 0.01) of these variables with the fruit produced by the plant (Table 3).
The fruit yield, found in each of the sites, was affected by soil characteristics (pH, MO, NT and K), which in turn were affected by the altitude of the sites. The number of fruits obtained was not very high because the sampling was done before the plant completed its reproductive cycle, however, in all the sites the phenological stage in which the poblano chili crops were found was very similar, which allowed to evaluate to a large extent the production of fruits that was had per plant and to compare between sites.
Conclusions
The present study highlights the importance of poblano chili culture and its association with soil microorganisms, mainly those that are beneficial for the soil-plant ecosystem. The altitudinal gradient of the sites was a determining factor in the chemical properties of the soil and in the distribution of the bacterial communities evaluated in this study.
The problems faced by poblano chili in the Sierra Nevada can be neutralized by decreasing the use of chemical products, which would imply less soil erosion and the maintenance of bacterial colonies, which could favor crop yield. However, additional studies are required to select these beneficial microorganisms that, when inoculated as biofertilizers, promote the growth of the plant and its adaptation to the field, when these are transplanted.
Literatura citada
Adeboye, M. K. A.; Iwuafor, E. N. O. and Agbenin, J. O. 2006. The effects of crop rotation and nitrogen fertilization on soil chemical and microbial properties in a Guinea Savanna Alfisol of Nigeria. Plant and Soil. 281(1-2): 97-107. [ Links ]
Aislabie, J. and Deslippe, J. R. 2013. Soil microbes and their contribution to soil services. In: Dymond, J. R. (Ed.). Ecosystem services in New Zealand - conditions and trends. Lincoln, New Zealand, Manaaki Whenua Press. 112-161 pp. [ Links ]
Allen, M. F. 2009. Water Relations in the Mycorrhizosphere. In: progress in botany. Lüttge U.; Beyschlag, W.; Büdel, B. and Francis, D. (Eds). California USA: Springer Berlin Heidelberg. Progress Bot. 70(1):257-276. [ Links ]
Altomare, C. and Tringovska, I. 2011. Beneficial soil microorganisms, an ecological alternative for soil fertility management. In: genetics, biofuels and local farming systems. Lichtfouse, E. (Ed.). Sustainable Agriculture Reviews, Springer Netherlands. 7(1):161-214. [ Links ]
Bago, B.; Pfeffer, P. E. and Shachar-Hill, Y. 2000. Carbon metabolism and transport in arbuscular mycorrhizas. Plant Physiol. 124(3):949-958. [ Links ]
Bautista-Calles, J.; García-Espinosa, R.; Montes-Belmont, R.; Zavaleta-Mejía, E.; Pérez-Moreno, J.; Ferrera-Cerrato, R.; García, C. R. y Huerta-Lara, M. 2010. Disminución de la marchitez del chile por introducción de antagonistas en cultivos de rotación. Interciencia. 35(9):673-679. [ Links ]
Bierman, B. and Linderman, R. G. 1981. Quantifying vesicular-arbuscular mycorrhizae: A proposed method towards standardization. New Phytologist. 87(1):423-432. [ Links ]
Bryant, J. A.; Lamanna, C.; Morlon, H.; Kerkhoff, A. J.; Enquist, B. J. and Green, J. L. 2008. Microbes on mountainsides: contrasting elevational patterns of bacterial and plant diversity. Proc. Natl. Acad. Sci. USA 105:05-11. [ Links ]
Coutinho, E. S.; Fernandes, G. W.; Berbara, R. L. L.; Valério, H. M. and Goto, B. T. 2015. Variation of arbuscular mycorrhizal fungal communities along an altitudinal gradient in rupestrian grasslands in Brazil. Mycorrhiza. 25(8):627-638. [ Links ]
Cyphers, A.; Escalante, G. P.; García-Bárcena, J.; García, M. B.; López, L. L.; Matos, M. E.; Nalda, E.; Noriega, A. M. N.; Pacheco, J. E. y Uriarte, C. M. T. 2009. Los chiles de México catálogo visual. Arqueología Mexicana. Edición especial núm. 32. 90 p. [ Links ]
Davies, Jr , F. T.; Potter, J. R. and Linderman, R. G. 1992. Mycorrhiza and repeated drought exposure affect drought resistance and extraradical hyphae development of pepper plants independent of plant size and nutrient content. J. Plant Physiol . 139(3):289-294. [ Links ]
Duca, D.; Lorv, J.; Patten, C. L.; Rose, D. and Glick, B. R. 2014. Indole-3-acetic acid in plant-microbe interactions. Antonie van Leeuwenhoek. J. Microbiol. 106(1):85-125. [ Links ]
Dumbrell, A. J.; Nelson, M.; Helgason, T.; Dytham, C. and Fitter, A. H. 2010. Idiosyncrasy and overdominance in the structure of natural communities of arbuscular mycorrhizal fungi: role of stochastic processes. J. Ecol. 98(2):419-428. [ Links ]
FAO. 2006. Organización de las Naciones Unidas para la Agricultura y la Alimentación. Base referencial mundial del recurso suelo. Informes sobre Recursos Mundiales de Suelos No. 103. IUSS, ISRIC, FAO, Roma, Italia. 117 p. [ Links ]
Galicia, L.; Gamboa, C. A. M.; Cram, S.; Chávez, V. B.; Peña, R. V.; Saynes, V. y Siebe, C. 2016. Almacén y dinámica del carbono orgánico del suelo en bosques templados de México. Terra Latinoam. 34(1):1-29. [ Links ]
Gerdemann, J. W. and Nicolson, T. H. 1963. Spores of mycorrhizal endogone species extracted from soil by wet sieving and decanting. Transactions of the British Mycological Society. 46(2):235-244. [ Links ]
Ghorbani, R.; Koocheki, A.; Brandt, K.; Wilcockson, S. and Leifert, C. 2010. Organic agriculture and food production: ecological, environmental, food safety and nutritional quality issues. in: sociology, organic farming, climate change and soil science. Chapter 4. Sustainable Agriculture Reviews. 3(1):77-107. [ Links ]
Hariprasad, P. and Niranjana, S. R. 2009. Isolation and characterization of phosphate solubilizing rhizobacteria to improve plant health of tomato. Plant Soil. 316(1-2):13-24. [ Links ]
Herridge, D. F.; Peoples, M. B. and Boddey, R. M. 2008. Global inputs of biological nitrogen fixation in agricultural systems. Plant Soil . 311(1-2):1-18. [ Links ]
Huerta, P. A.; Fernández, R. S. y Ocampo, F. I. 2007. Manual de chile poblano, importancia económica y sociocultural. Colegio de Postgraduados Campus Puebla. Fundación Produce Puebla AC Altres costa- Amic. Primera edición. México. 80 p. [ Links ]
Hofmann, K.; Lamprecht, A.; Pauli, H. and Illmer, P. 2016. Distribution of prokaryotic abundance and microbial nutrient cycling across a high-alpine altitudinal gradient in the Austrian Central Alps is affected by vegetation, temperature, and soil nutrients. Microb. Ecol. 72(3):704-716. [ Links ]
INEGI. 2017. Instituto Nacional de Estadística y Geografía. Datos. México en cifras. Puebla. http://www.beta.inegi.org.mx/app/areasgeograficas/?ag=21. [ Links ]
Javaid, A. 2009. Arbuscular mycorrhizal mediated nutrition in plants. J. Plant Nutr. 32(10):1595-1618. [ Links ]
Kim, K.; Yim, W.; Trivedi, P.; Madhaiyan, M.; Boruah, H. P. D.; Rashedul, M. I.; Lee, G. and Sa, T. 2010. Synergistic effects of inoculating arbuscular mycorrhizal fungi and Methylobacterium oryzae strains on growth and nutrient uptake of red pepper (Capsicum annuum L.). Plant Soil . 327(1):429-440. [ Links ]
Mader, P.; Fliebbach, A.; Dubois, D.; Gunst, L.; Fried, P. and Niggli, U. 2002. Soil fertility and biodiversity in organic farming. Science. 296(5573):1694-1697. [ Links ]
Massenssini, A. M.; Bonduki, V. H. A.; Melo, C. A. D.; Tótola, M. R.; Ferreira, F. A. and Costa, M. D. 2014. Soil microorganisms and their role in the interactions between weeds and crops. Planta Daninha, Viçosa-MG. 32(4):873-884. [ Links ]
Mena, V. H.; Ocampo, J. O.; Dendooven, L.; Martínez, S. G.; González, C. J.; Davies, F. T. and Olalde-Portugal, V. 2006. Arbuscular mycorrhizal fungi enhance fruit growth and quality of chile ancho (Capsicum annuum L. cv San Luis) plants exposed to drought. Mycorrhiza. 16(4):261-267. [ Links ]
Oehl, F.; Sieverding, E.; Mäder, P.; Dubois, D.; Ineichen, K.; Boller, T. and Wiemken, A. 2004. Impact of long-term conventional and organic farming on the diversity of arbuscular mycorrhizal fungi. Oecologia. 138(4):574-583. [ Links ]
Oehl, F.; Oberson, A.; Tagmann, H. U.; Besson, J. M.; Dubois, D.; Mader, P.; Roth, H. R. and Frossard, E. 2002. Phosphorus budget and phosphorus availability in soils under organic and conventional farming. Nutr Cycl. Agroecosyst. 62(1):25-35. [ Links ]
Pasaribu, A.; Mohamad, R. B.; Awang, Y.; Othman, R. and Puteh, A. 2011. Growth and development of symbiotic arbuscular mycorrhizal fungi, Glomus mosseae (Nicol. and Gerd.), in alachlor and glyphosate treated soils. African Journal of Biotechnology. 10(55):11520-11526. [ Links ]
Perner, H.; Schwarz, D.; Bruns, C.; Mäder, P. and George, E. 2007. Effect of arbuscular mycorrhizal colonization and two levels of compost supply on nutrient uptake and flowering of pelargonium plants. Mycorrhiza . 17(5):469-474. [ Links ]
Phillips, J. M. and Hayman, D. S. 1970. Improved procedures for clearing roots and staining parasitic and vesicular-arbuscular mycorrhizal fungi for rapid assessment of infection. Transactions of the British Mycological Society . 55(1):158-161. [ Links ]
Restrepo, F. G. M.; Marulanda, M. S.; de la Fe, P. Y.; Díaz-de la Osa, A.; Lucia, B. V. y Hernández, R. A. 2015. Bacterias solubilizadoras de fosfato y sus potencialidades de uso en la promoción del crecimiento de cultivos de importancia económica. CENIC Ciencias Biológicas. 46(1):63-76. [ Links ]
Rodríguez, J.; Peña, O. B. V.; Gil, M. A.; Martínez, C. B.; Manzo, F. y Salazar, L. L. 2007. Rescate in situ del chile poblano en Puebla, México. Rev. Fitotec. Mex. 30(1):25-32. [ Links ]
Sandhya, V.; Ali, S. Z.; Venkateswarlu, B.; Reddy, G.; Grover, M. 2010. Effect of osmotic stress on plant growth promoting Pseudomonas spp. Archives of Microbiology. 192(10):867-876. [ Links ]
Saraf, M.; Kumar, J. C. and Patel, D. 2011. The role of ACC deaminase producing PGPR in sustainable agriculture. In: plant growth and health promoting bacteria (D. K. Maheshwari. (Ed.). Springer Berlin Heidelberg. Microbiol. Monographs. 18(1):365-385. [ Links ]
SAS. 2002. Statistical Analysis System. Software: the SAS System for Windows version 9.0. SAS Institute Inc., Cary, NC 25513, USA. [ Links ]
Sawers, R. J. H.; Gutjahr, C. and Paszkowski, U. 2008. Cereal mycorrhiza: an ancient symbiosis in modern agriculture. Trends Plant Sci. 13(2):93-97. [ Links ]
SDR. 2007. Secretaría de Desarrollo Rural de Puebla. Cadenas productivas agropecuarias y acuícolas del estado de Puebla. Primera edición. Gobierno del estado de Puebla. México. 97 p. [ Links ]
Smith, S. E. and Read, D. J. 2008. Mycorrhiza l symbiosis. Academic Press. 3th . (Ed.). New York, USA. 800 p. [ Links ]
Stott, M. B. and Taylor, M. W. 2016. Microbial ecology research in New Zealand. New Zealand J. Ecol. 40(1):12-28. [ Links ]
Subramanian, K. S.; Santhanakrishnan, P. and Balasubramanian, P. 2006. Responses of field grown tomato plants to arbuscular mycorrhizal fungal colonization under varying intensities of drought stress. Sci. Hortic. 107(3):245-253. [ Links ]
Wolfe, B. E. and Klironomos, J. N. 2005. Breaking new ground: soil communities and exotic plant invasion. BioSci. 55(6):477-487. [ Links ]
Weyens, N.; van der Lelie, D.; Taghavi, S. and Vangronsveld, J. 2009. Phytoremediation: plant-endophyte partnerships take the challenge. Curr Opin Biotechnol. 20(2):248-254. [ Links ]
Zhu, C. X.; Song, B. F. and Xu, W. H. 2010. Arbuscular mycorrhizae improve low temperature stress in maize via alterations in host water status and photosynthesis. Plant Soil . 331(1-2):129-137. [ Links ]
Received: January 2018; Accepted: March 2018
 Este es un artículo publicado en acceso abierto bajo una licencia Creative Commons
Este es un artículo publicado en acceso abierto bajo una licencia Creative Commons
