











 text new page (beta)
text new page (beta) Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO 
 Permalink
Permalink1. Introducción
El Orden Proboscidea actualmente incluye tres especies, el elefante africano de sabana Loxodonta africana, el elefante africano de bosque Loxodonta cyclotis y el elefante asiático Elephas maximus, todas incluidas en la familia Elephantidae (Shoshani, 2005). Este orden fue mucho más diverso en el pasado, ya que se han contabilizado 175 especies y subespecies, clasificadas en 42 géneros y 10 familias (Shoshani y Tassy, 2005). Proboscidea tuvo su origen en el Paleoceno tardío, a lo largo de la costa del Mar Tethys en el norte de África y sur de Asia, se extendió a Eurasia durante el Mioceno temprano y, posteriormente, alcanzó América del Norte vía Beringia en el Mioceno tardío (Lambert y Shoshani, 1998).
Proboscidea está representado en el Pleistoceno de México por tres familias, Mammuthidae, con un sólo género: Mammut; Gomphotheriidae, con dos géneros: Cuvieronius y Stegomastodon y Elephantidae, con un género: Mammuthus (Arroyo-Cabrales et al., 2010). De estos, Mammuthus es el más comúnmente encontrado a lo largo y ancho del territorio mexicano. En la literatura se mencionan cinco especies para México: Mammuthus columbi, M. hayi, M. imperator, M. primigenius y M. sonoriensis (Arroyo-Cabrales et al., 2003b). Actualmente sólo se reconocen dos especies, el llamado complejo Mammuthus hayi/ meridionalis que se conoce de localidades del Pleistoceno temprano (Irvingtoniano) en el norte y centro del país y Mammuthus columbi, que se encuentra en sedimentos del Pleistoceno tardío (Rancholabreano) a lo largo y ancho de México, desde localidades norteñas como El Carrizal, en Baja California Sur (Rochín-Bañaga et al., 2017), hasta la más sureña ubicada en El Amate, en Chiapas (Gómez-Pérez y Carbot-Chanona, 2012), siendo Yucatán y Tabasco los únicos estados sin reportes hasta el momento (Arroyo-Cabrales et al., 2010). Mammuthus columbi (el mamut de Cristoforo Colombo, en referencia al Nuevo Mundo) fue descrito por Hugh Falconer (1857) bajo el nombre de Elephas columbi. Esta especie se distribuyó desde Canadá hasta Centroamérica (Kurtén y Anderson, 1980; Alvarado, 1986; Laurito, 1988; Lucas et al., 1997; Laurito y Aguilar, 2006) y se cree evolucionó en América del Norte a partir de un primitivo inmigrante euroasiático, similar a Mammuthus trogontherii (Lister y Sher, 2015).
1.1. REGISTROS DE MAMMUTHUS EN PUEBLA
Los primeros restos de Mammuthus inequívocamente identificados, provenían de las localidades de San Francisco Totimehuacan y San Baltazar Tétela, y estuvieron resguardados en el gabinete del antiguo Colegio del estado, hoy Benemérita Universidad Autónoma de Puebla (Armenta-Camacho, 1978). Posteriormente, se adicionaron registros de Mammuthus gracias a los trabajos de Félix y Lenk (1891), Osborn (1905, 1942), Freudenberg (1922), Armenta-Camacho (1957, 1978), Irwin-Williams (1967), Silva-Bárcenas (1969), Lagunas-Rodríguez y Suárez-Cruz (1997), Suárez-Cruz y Lagunas-Rodríguez (2008), Lagunas-Rodríguez et al. (2013), entre otros.
En la última década, en Puebla se han reportado hallazgos de mamuts cuyos restos son expuestos como consecuencia de la construcción de caminos y edificaciones, siendo la mayoría atendidos por personal del Instituto Nacional de Antropología e Historia, delegación Puebla (Lagunas-Rodríguez y Suárez-Cruz, 1997; Lagunas-Rodríguez et al., 2013). En junio de 2007 y marzo de 2013, se atendieron las denuncias sobre el hallazgo de restos fósiles en barranca El Morillo, al norte de Ciudad Serdán, municipio de Chalchicomula de Sesma y en la colonia Las Cruces, municipio de Amozoc. En ambos casos, se rescataron restos óseos de Mammuthus que son el motivo del presente manuscrito.
A pesar de que los registros de mamuts son frecuentes en México y los restos abundantes, poco de ese material ha sido descrito formalmente, por lo que se desconocen las variaciones intraespecíficas entre poblaciones, preferencia de hábitat o aspectos generales sobre su paleobiología, como la edad y el sexo. Por tal motivo, el objetivo de este trabajo es describir formalmente el material de Mammuthus encontrado en el centro-este de Puebla, así como aportar datos sobre algunos aspectos biológicos de los individuos, como edad, sexo, dieta y preferencia de hábitat, información que sin duda contribuirá a entender mejor la paleobiología de los mamuts en México.
Abreviaturas institucionales: CRINAH, Colección Regional, Instituto Nacional de Antropología e Historia, Centro INAH-Puebla, Puebla de Zaragoza, Puebla.
2. Área de estudio y estratigrafía local
El material descrito en este trabajo proviene de dos localidades en la región centro-este del estado de Puebla. La primera localidad se denomina barranca El Morillo y se localiza a 2 km al norte de Ciudad Serdán, municipio de Chalchicomula de Sesma (Figura 1). En este sitio aflora una secuencia de sedimentos fluviolacustres de 4.5 m de espesor, donde se observan tres secciones litológicas diferentes. La inferior con un espesor de 2.9 m, está conformada por arcilla con una fracción de arena fina que intemperiza a color café claro y en donde, además, hay presencia de pequeños lentes de ceniza volcánica de entre 15 a 20 cm de espesor; este estrato es donde se encontró el material de Mammuthus correspondiente al ejemplar CRINAH-2275. A su vez este estrato de arcilla es sobreyacido por un estrato de arena gruesa de color gris oscuro de aproximadamente 1 m de espesor, donde están presentes litoclastos subredondeados de cuarzo, sienita, basalto, andesita y granito de tamaños que van desde 5 cm hasta 20 cm de diámetro. El estrato superior tiene 60 cm de espesor y está conformado por arena fina con alto contenido de materia vegetal. En esta sección hay presencia de paleocanales, los cuales están rellenos por arena gruesa que contienen además pequeños litoclastos rodados de cuarzo, calcita, basalto, granito y andesita, que van de 0.50 a 2 cm de diámetro (Figura 2A).
La segunda localidad se denomina Sierra de Amozoc y se localiza a 3.5 km al sur de Amozoc de Mota, en la ladera oriente de una loma a 40 metros del antiguo camino a Cuautinchan, en las inmediaciones de la colonia Las Cruces (Figura 1). La secuencia estratigráfica expuesta mide 2.20 m de espesor y se observan tres niveles sedimentarios. El estrato inferior tiene 1.40 m de espesor y está conformado por arena de grano medio y toba andesítica de color ocre muy consolidada; en esta sección se encontraron los restos de Mammuthus que corresponden al ejemplar CRINAH-2274. La sección inmediata superior está conformada por un estrato de aproximadamente 45 cm de arena gruesa a mediana con pequeños litoclastos subredondeados de rocas volcánicas. Finalmente, la secuencia culmina con un estrato de 50 - 55 cm de arena fina y limo que presenta un alto contenido de materia vegetal (Figura 2B).
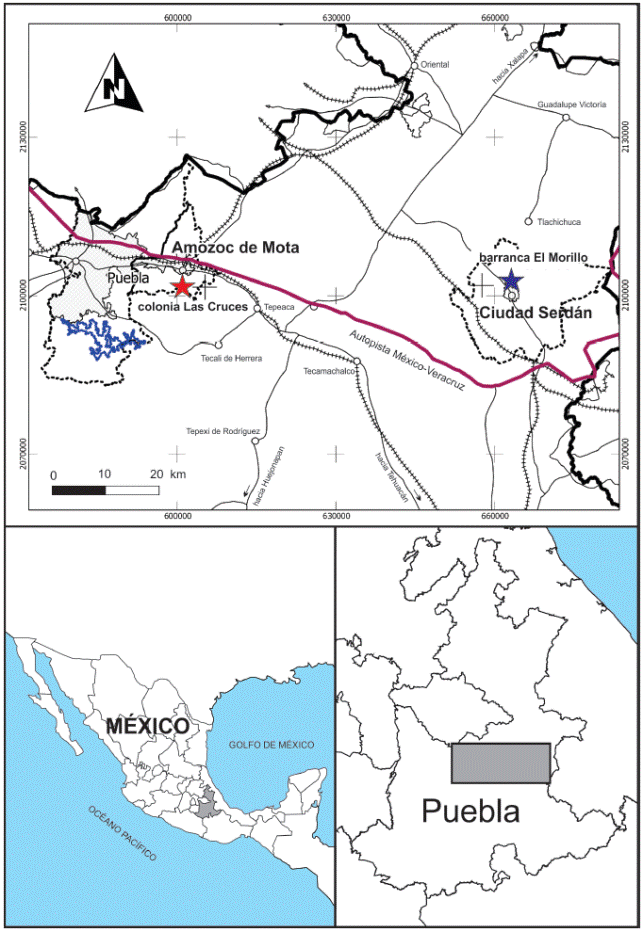
Figura 1 Ubicación de las localidades con restos de Mammuthus columbi descritos en este trabajo. La estrella azul marca la ubicación de la localidad barranca El Morillo, cerca de Ciudad Serdán; la estrella roja marca la ubicación de la localidad Sierra de Amozoc, en las inmediaciones de la colonia Las Cruces, municipio de Amozoc de Mota.

3. Material y método
3.1. MATERIAL
Se describen dos ejemplares; el primer ejemplar fue recolectado en Sierra de Amozoc y está catalogado como CRINAH-2274 y comprende un cráneo parcialmente deformado que conservó en su mayoría la parte derecha; el M2 derecho, el M2 y M3 izquierdos y parte de la defensa izquierda en mal estado de conservación. El segundo ejemplar fue rescatado en la barranca El Morillo y está catalogado como CRINAH-2275, e incluye el M2 y M3 derechos, M2 y M3 izquierdos; fémur derecho al cual le falta el extremo proximal y distal; tibia izquierda parcialmente conservada, vértebras incompletas y varios fragmentos de costillas. Ambos ejemplares están resguardados en el laboratorio de Paleontología del Centro INAH-Puebla, del Instituto Nacional de Antropología e Historia.
3.2. OBTENCIÓN DE MEDIDAS Y FOTOGRAFÍAS
Las medidas craneales se obtuvieron siguiendo a Gillette y Madsen (1993), mientras que las medidas dentales y postcraneales se sacaron siguiendo las recomendaciones de Agenbroad (1994). Los huesos de gran talla fueron medidos con una cinta métrica. Las medidas dentales se obtuvieron con un calibrador digital con una precisión de 0.1 mm. En las Figuras 3 y 4 se muestra cómo se tomaron las medidas del cráneo y molares.

Figura 3 Esquema que muestra cómo se tomaron las medidas en el cráneo: 1) longitud total: proyectado sobre la línea media (acrocraniónprostión); 2) longitud basal (basión-prostión; 3) longitud de la caja craneal (ectorbital-opisthion); 4) longitud facial lateral (ectorbitalprostión); 5) longitud basión-staphilion; 6) longitud del paladar (staphilion-prostión; 7) longitud del diastema: desde el alveolo del M3 hasta el alveolo del incisivo (medido desde el punto medio del alveolo); 8) diámetro transverso interno del alveolo del incisivo; 9) ancho máximo de la curvatura del maxilar en el punto medio del alveolo del M3; 10) ancho máximo posterior del paladar: medido desde el extremo posterior de los alvéolos del M3; y 11) altura del cráneo: desde el paladar hasta el acrocranión. Abreviaturas: A, acrocranión; B, basión; Ec, ectorbital; O, opisthion; P, prostión; S, staphilion. Modificado de Gillette y Madsen (1993).

Figura 4 Esquema que muestra cómo se tomaron las medidas en los molares. Abreviaturas: H, altura máxima del diente paralela a la orientación de las láminas; Ld, longitud máxima en plano diagonal; Lo, longitud máxima en el plano oclusal. Modificado de Agenbroad (1994).
Se obtuvo la frecuencia lamelar de cada molar; esta se refiere al número promedio de placas en 100 mm tomados a lo largo del eje anteroposterior en la superficie oclusal del molar (Maglio, 1973). Las fotografías se obtuvieron con una cámara digital Canon PowerShot SX40 HS de 12.1 megapixeles y fueron procesadas en CorelDraw X8.
3.3. NOMENCLATURA CRANEAL Y DENTAL
En elefantes los primeros tres dientes son premolares deciduos y aunque las láminas dentales de los premolares permanentes están presentes en el epitelio oral, estos no se desarrollan (Kozawa et al., 2001). Por consiguiente, aunque existen varios sistemas nomenclaturales para los molariformes de Elephantidae, nosotros preferimos el propuesto por Saunders (1970), por lo que nombramos la serie como dP2, dP3, dP4, M1, M2, y M3, ya que consideramos se apega más a la nomenclatura tradicional de mamíferos; contrariamente al propuesto por Laws (1966), que nombra a los molariformes como M1, M2, M3, M4, M5 y M6, en la cual el dP2, dP3 y dP4 son nombrados como M1, M2 y M3 respectivamente.
La nomenclatura morfológica usada en la descripción del cráneo está basada en Van der Merwe et al. (1995).
3.4. ESTIMACIÓN DE LA EDAD
La edad de los ejemplares se calculó siguiendo a Haynes (1991) y se expresa en Años de Elefante Africano (AEA), según la escala de Laws (1966).
3.5. INFERENCIA DE LA DIETA
Para evaluar la dieta de los ejemplares se usó la metodología de ángulos de mesodesgaste para elefántidos propuesta por Saarinen et al. (2015), la cual consiste en medir el ángulo formado por las paredes de esmalte al interior del valle de tres láminas en uso; con estas tres medidas se obtiene la media. Los ángulos por arriba de 130° indican una dieta estrictamente pacedora (> 90 % de plantas C4); de 130° a 124° la dieta es dominada por pastos, pero hay ingesta de plantas arbustivas (> 70 % de plantas C4); de 124° a 113° la dieta es mixta (ingesta casi igual de plantas C3 y C4); de 113° a 106° la dieta es dominantemente ramoneadora, pero hay ingesta de pastos (> 70 % de plantas C3); y los ángulos por debajo de los 106° indican dieta estrictamente ramoneadora (> 90 % de plantas C3).
Los ángulos se obtuvieron con un medidor de contornos, el cual se posicionó sobre la superficie oclusal de cada molar para obtener el relieve de los valles de las láminas. Posteriormente, se midió el ángulo del contorno de tres láminas en uso y se obtuvo la media (Figura 5).

4. Resultados
4.1. PALEONTOLOGÍA SISTEMÁTICA
Clase Mammalia Linnaeus, 1758 Superorden Paenungulata Simpson, 1945 Orden Proboscidea Illiger, 1811 Familia Elephantidae Gray, 1821 Género MammuthusBrookes, 1828Mammuthus columbi (Falconer, 1857)
Holotipo. BM(NH)40769, M3 incompleto con 10 láminas conservadas, colectado en el Canal de Brunswick, cerca de Darien Georgia, en el año de 1838 durante la excavación del canal y actualmente alojado en el British Museum of Natural History (Maglio, 1973; Patterson et al., 2012). Sinonimias. Archidiskodon imperator (Leidy, 1858), Elephas (Euelephas) imperator (Leidy, 1858), Euelephas imperator (Leidy, 1858), Mammuthus imperator (Leidy, 1858), Mammuthus (Archidiskodon) imperator (Leidy, 1858), Elephas jeffersonii (Osborn, 1922), Parelephas roosevelti (Hay, 1922), Elephas washingtonii (Osborn, 1923), Parelephas progressus (Osborn, 1924), Elephas eellsi (Hay, 1926), Archidiskodon sonoriensis (Osborn, 1929), Elephas floridanus (Osborn, 1929), Parelephas columbi (Osborn, 1929), Parelephas floridanus (Osborn, 1929), Parelephas jeffersonii (Osborn, 1930).
4.2.1. EJEMPLAR CRINAH-2274
El cráneo es más alto que ancho, con el frontal y el parietal extendiéndose posterodistalmente (Figura 6A). La parte posterior del parietal y supraoccipital están perdidos y permiten visualizar la neumatización interior del cráneo. Sólo está presente el meatus acusticus externus derecho y este se observa como una abertura de forma circular que se ubica por detrás del arco zigomático. El arco zigomático derecho está completo y se extiende desde la parte anterior del M2 hasta la parte posterior del M3. La facies orbitalis es profunda y se encuentra bordeada en su parte superior por el margo supraorbitalis. Únicamente está conservada la narina externa derecha, la cual se aprecia como una abertura de forma semilunar que se ubica en la parte frontal del cráneo. En la parte posterior se observa el foramen magnum, el cual es amplío y de forma circular (Figura 6B). El processus alveolaris derecho está conservado parcialmente y se orienta anterolateralmente, lo que indica que ambos incisivos son divergentes. El alveolo del incisivo derecho no está completo, pero la parte conservada permite estimar su diámetro en 210 mm (Tabla 1). Ventralmente se observan ambos palatinos pareados, estos son una lámina delgada orientada anteroposteriormente. Ambos palatinos se fusionan lateralmente con los procesos palatinos del maxilar. En la parte posterior de cada palatino se observa el foramen palatinum major. El vómer es una lámina delgada que divide el piso de la cavidad nasal y el techo del pasaje nasofaríngeo. Ambos canalis infraorbitalis están presentes y se localizan en la parte anterior de los alvéolos de los M2.

Figura 6 Ejemplar CRINAH-2274, de Sierra de Amozoc, Puebla. Cráneo en vista A) lateral (lado derecho), B) posterior y C) ventral. Barra de escala igual a 200 mm. Abreviaturas: ci, canalis infraorbitalis; co, condylus occipitalis; fmg, foramen magnun; for, facies orbitalis; fpm, foramen palatinum major; mae, meatus acusticus externus; p, parietal; pa, processus alveolaris; pl, palatino; ppl, processus palatinus; pz, processus zygomaticus; vo, vómer.
Tabla 1 Medidas del cráneo CRINAH-2274.
| Dimensiones | mm |
| 1. Longitud total: proyectado sobre la línea media (acrocranión-prostión). | 1250 |
| 2. Longitud basal (bastión-prostión). | 850 |
| 3. Longitud de la caja craneal (ectorbital-opisthion). | 410 |
| 4. Longitud facial lateral (ectorbital-prostión). | 700 |
| 5. Longitud basión- staphilion | 290 |
| 6. Longitud del paladar (staphilion-prostión). | 680 |
| 7. Longitud del diastema: desde el alveolo del M3 hasta el alveolo del incisivo (medido desde el punto medio del alveolo). | 260 |
| 8. Diámetro trasverso interno del alveolo del incisivo. | 220 |
| 9. Ancho máximo de la curvatura del maxilar en el punto medio del alveolo del M3. | 100 |
| 10. Ancho máximo posterior del paladar: medido desde el extremo posterior de los alveolos del M3. | 120 |
| 11. Altura del cráneo: desde el paladar hasta el acrocranión. | 810 |
Están conservados el M2 izquierdo y el M3 derecho e izquierdo. El M2 es cuadrangular, sus medidas son 90 mm de largo por 90 mm de ancho; solamente cinco láminas están presentes. Los M3 poseen 22 láminas (Figura 6C). La superficie oclusal en uso de los M3 presenta nueve láminas y tienen entre 140 - 150 mm de largo y 90 - 100 mm de ancho. La frecuencia lamelar es de 6 a 7. Tanto en el M2 como los M3 las láminas son delgadas. El esmalte es fuertemente crenulado, con un grosor de 1.5 a 4mm.
El fragmento de defensa está muy mal conservado, pero se aprecia que corresponde a la porción proximal y tiene una longitud de 1050 mm.
4.2.2. EJEMPLAR CRINAH-2275
El M2 derecho está fuertemente desgastado y sólo conserva tres láminas; su forma es cuadrangular y mide 80 mm de longitud por 89 mm de ancho. En el M3 derecho la superficie en uso mide 126 mm de largo por 91 mm de ancho; posee un total de 20 láminas, de las cuales ocho están en uso. Por su parte, la superficie oclusal del M2 izquierdo tiene una longitud de 115 mm y 84 mm de ancho; conserva únicamente tres láminas (Tabla 2). El M3 izquierdo posee 21 láminas de las cuales ocho están en uso. En todos los molares las láminas son delgadas; el esmalte es crenulado, con un grosor de 2.0 a 3.5 mm (Figura 7). Frecuencia lamelar de seis láminas en 100 mm.
Tabla 2 Medidas (en mm) de los M3 del ejemplar CRINAH 2275.
| Molar | Ld | Lo | H |
| M3 derecho | 260 | 208 | 199 |
| M3 izquierdo | 220 | 170 | 180 |

Figura 7 Ejemplar CRINAH-2275, de Barranca El Morillo, Puebla. M2 y M3 derechos en vista A) labial, B), lingual y C) oclusal; D) M2 y M3 izquierdos en vista oclusal. Barra de escala igual a 100 mm.
La tibia únicamente conserva la diáfisis, ya que los extremos distal y proximal están perdidos. La diáfisis es de forma suboval en corte transversal, pero hacia los extremos se ensancha abruptamente en sentido lateral-medial; el extremo proximal es más ancho que el distal.
Sólo se conservó la parte proximal de la diáfisis del fémur, la cual es de forma tubular. La forma y proporciones indican que se trata de un hueso largo, pero delgado.
4.3. EDAD
La edad de Mammuthus puede ser estimada usando el tamaño y grado de desgaste de los molares (Haynes, 1991), o el grado de fusión entre la epífisis y diáfisis en huesos apendiculares (Lister, 1999). Debido a que sólo un individuo posee huesos apendiculares y estos no están completos, la edad se estimó usando las variables dentales antes mencionadas.
Los M2 de M. columbi tienen un rango de 12 a 15 láminas y los M3 de 17 a 22. El ejemplar CRINAH-2274, conserva un M2 izquierdo con cinco láminas lo que indica que ha perdido entre un 60 - 67 % del total del molar. El M3 izquierdo tiene en
uso ocho láminas y el M3 derecho nueve láminas de un total de 22, que muestra que los molares tienen un desgaste del 41 %. Según Haynes (1991), estas características ubican al ejemplar en la clase XX de Laws (1966), que sugiere tiene aproximadamente 34 AEA.
El ejemplar CRINAH-2275 conserva ambos M2 y M3. En los M2 sólo están presentes tres láminas de un promedio de 12 - 15, que indican la pérdida del 73 % de los molares. Los M3 poseen 11 láminas en uso de las 20 totales, que muestran un desgaste en los molares del 55 %. Siguiendo a Haynes (1991), el ejemplar CRINAH-2275 se ubica entre las clases XXI y XXII de Laws (1966), con una edad aproximada de 37-41 AEA.
4.4. SEXO DE LOS INDIVIDUOS
En Mammuthus el sexo de un individuo puede ser determinado con base en tres criterios: tamaño y robustez del esqueleto, morfología de la pelvis y dimensiones de las defensas y sus alvéolos (Lister y Agenbroad, 1994). Desafortunadamente, debido a lo incompleto de los ejemplares, sólo pudo identificarse el sexo del ejemplar CRINAH-2274, quien puede ser referido como un macho, basado en el diámetro del alveolo (210 mm).
4.5. DIETA
El valor promedio de los ángulos obtenidos en el valle interior de tres láminas en los M3 del ejemplar CRINAH-2274 fue de 136° y para el M3 del ejemplar CRINAH-2275 fue de 134.5°. Esto indica que ambos especímenes eran pacedores estrictos con una dieta conformada principalmente por plantas C (Saarinen et al., 2015).
5. Discusión
5.1. IDENTIDAD TAXONÓMICA
En México llegaron a describirse varias especies de mamuts, pero actualmente sólo se reconocen dos, Mammuthus hayi que se conoce únicamente para localidades del Pleistoceno temprano (Irvingtoniano) en Arizpe, Sonora; El Mezquital, Baja California; y Culhuacán, Ciudad de México (Arroyo-Cabrales et al., 2007). M. columbi se ha encontrado en sedimentos del Pleistoceno tardío (Rancholabreano), en casi todos los estados, a excepción de la Península de Yucatán y Tabasco (Arroyo-Cabrales et al., 2010). No obstante, Arroyo-Cabrales et al. (2007), mencionan que la identificación del material de M. hayi en México es cuestionable. Por su parte Lister y Sher (2015) argumentan que los M3 de M. hayi de El Mezquital Baja, California y Culhuacán, Ciudad de México, caen dentro del rango de medidas y frecuencia lamelar de M. columbi. Lo anterior cuestiona la presencia de M. hayi en México.
Sin embargo, nosotros preferimos reconocer conservadoramente la presencia de ambas especies a raíz de la falta de evidencia concreta que señale la inexistencia de M. hayi en el territorio mexicano. Los molares son una fuente altamente diagnóstica para identificar las especies dentro del género Mammuthus (Maglio, 1973), por ende, nuestra asignación se basa en la morfología y medidas de los M3 de ambos individuos. Mammuthus hayi tiene un M3 con un número bajo de láminas, 12 en total, por lo que la frecuencia lamelar es de 4; esta es la principal diferencia con el M3 de M. columbi que tiene una frecuencia lamelar de 5 a 7 (Arroyo-Cabrales et al., 2003a). Adicionalmente, el M3 de Mammuthus columbi tienen una longitud total de 250 - 320 mm, con un ancho de 75 a 108.5 mm; esmalte fuertemente crenulado con un grosor de 1.5 a 3.8 mm y poseen de 17 a 22 láminas (Arroyo-Cabrales et al., 2003a; McDaniel y Jefferson, 2006). Las características morfológicas y morfométricas de los M3 de los ejemplares CRINAH-2274 y CRINAH-2275 caen dentro del rango de los caracteres diagnósticos para M. columbi, por lo que ambos individuos son asignados a esta especie.
5.2.1. EDAD Y SEXO
La determinación sexual y de la edad en mamíferos es importante para entender aspectos de su biología y ecología. En mamíferos extintos la determinación de estos aspectos biológicos es posible sólo en algunos casos. Por ejemplo, en Mammuthus se ha podido asignar edad y sexo a individuos equiparándolo con las especies de elefantes vivientes (Haynes, 1991).
En las especies de elefantes actuales hay diferencia de tamaño entre machos y hembras. En Loxodonta africana, los machos llegan a pesar de 4700 a 6048 kg y alcanzan una altura hasta los hombros de 3.20 a 4.01 m, mientras que las hembras tienen rangos de 2160 a 3232 kg y altura hasta los hombros de 2.20 a 2.60 m (Laursen y Bekoff, 1978). En Elephas maximus, los machos llegan a pesar hasta 5400 kg y alcanzan tallas de 3.20 m hasta lo hombros, mientras que las hembras llegan a pesar en promedio 2720 kg y medir 2.24 m hasta los hombros (Shoshani y Eisenberg, 1982). En Mammuthus columbi estas diferencias de talla también están presentes, siendo los machos más grandes que las hembras, tal como lo han demostrado estudios que incluyen restos de varios ejemplares encontrados en un mismo yacimiento (e.g. Lister y Agenbroad, 1994; McDaniel y Jefferson, 2006). En este sentido, las dimensiones de las defensas y sus alvéolos son de gran valor para la determinación del sexo (Lister y Agenbroad, 1994), ya que los machos tienen defensas más robustas, por lo que es posible separar machos de hembras basados en el diámetro de éstas (Vereshchagin y Tikhonov, 1986).
En el caso de los ejemplares descritos en este trabajo, sólo el ejemplar CRINAH-2274 pudo ser referido como un macho con base al diámetro del alveolo derecho. La dimensión del alveolo de CRINAH-2274 es de 210 mm. Esta medida es similar al del ejemplar apodado “Zed” colectado en Rancho La Brea, el cual tiene un diámetro de 240 mm, pero es ligeramente mayor al del individuo con número de entrada 88.18 de la Utah Division of State History, el cual es de 190 mm; ambos ejemplares referidos como machos (Gillette y Madsen, 1993; El Adli et al., 2015).
Los machos de Elephas y Loxodonta alcanzan la madurez sexual alrededor de los 15 años, aunque generalmente llegan a aparearse hasta los 30 años (Flower, 1943; Sukumar, 2003). La edad estimada para el ejemplar CRINAH-2274 es de 34 AEA y por lo tanto es un macho sexualmente maduro. En elefantes africanos actuales, los machos sexualmente maduros son excluidos de los grupos familiares dominados por hembras, por lo que viven solos o formando pequeños grupos con otros machos, excepto en las temporadas de apareamiento (Haynes, 1991; Moss y Poole, 1983). Esto podría explicar por qué cuando se encuentran restos de ejemplares solitarios de Mammuthus generalmente corresponden a machos adultos (e.g.Gillette y Madsen, 1993), como es el caso de CRINAH-2274.
Por su parte, el ejemplar CRINAH-2275 tiene una edad de entre 37 a 41 AEA, pero no presenta caracteres que permitan determinar su sexo. No obstante, como el ejemplar fue encontrado solo, podría también tratarse de un macho sexualmente maduro.
5.2.2. DIETA
Saarinen et al. (2015) encontraron que en las especies actuales de proboscídeos existe una estrecha relación entre los valores isotópicos de δ13C (que depende de la dieta) y el ángulo formado entre los valles de las láminas en uso de los molares. Basados en sus resultados, estos autores determinaron ángulos de mesodesgaste, método con el que es posible conocer la dieta de elefántidos extintos (ver apartado Material y Método). Los ejemplares CRINAH-2274 y CRINAH-2275 tienen un ángulo promedio de 135°, que indican que la dieta de ambos individuos estuvo basada principalmente en pastos (> 90 % de plantas C4). Esta dieta es similar al encontrado en los individuos de Rancho La Brea, California, los cuales tienen un promedio de 135.5° y se consideran estrictamente pacedores (Saarinen et al., 2015). Una dieta pacedora, basada en juncia (Carex spp.) y pastos, al parecer es característica en individuos de M. columbi procedentes de localidades rancholabreanas en Estados Unidos de América, tal como lo demuestran análisis isotópicos de los ejemplares de Ichetucknee, Florida; Ingleside, Texas; y Gila Pueblo, Arizona (Cerling et al., 1999); y análisis de heces procedentes de Salt Lake Meridian y Bechan Cave, Utah (Mead et al., 1986; Gillette y Madsen, 1993).
En México, la dieta de individuos M. columbi, provenientes de diversas localidades del centro y sur del país, ha sido determinada usando isótopos estables de δ13C y δ18O extraídos del esmalte dental, así como con unos pocos análisis de microdesgaste. En este trabajo, la determinación de la dieta de mamuts usando el método de ángulos de mesodesgaste se aplica por primera vez en México.
Los resultados obtenidos por los métodos arriba mencionados demuestran que la mayoría de los ejemplares tenían una dieta mixta basada en plantas C3 y C4, mientras que un menor número de individuos eran estrictamente pacedores con una ingesta exclusiva de plantas C4. Lo anterior se ha documentado en las localidades de El Cedral y Laguna de Las Cruces, en San Luis Potosí (Pérez-Crespo et al., 2009, 2013); Tocuila, Estado de México (Pérez-Crespo et al., 2010); Valsequillo, Puebla (Pérez-Crespo et al., 2014); La Cinta-Portalitos y La Piedad-Santa Ana, en el límite Michoacán-Guanajuato (Gutiérrez-Bedolla et al., 2016); El Amate, en la Depresión Central, Chiapas (Pérez-Crespo et al., 2015); y ahora en barranca El Morillo y Sierra de Amozoc, Puebla.
5.2.3. PREFERENCIA DE HÁBITAT
La inferencia de la dieta de mamíferos herbívoros extintos ha sido frecuentemente usada como un indicador de la vegetación presente en un área y tiempo determinados, ya que la ingesta de alimento está ligada estrechamente a las plantas que se encuentran disponibles en el entorno. Esto a su vez da una aproximación de las condiciones paleoambientales existentes del área geográfica en un determinado intervalo de tiempo (Rivals y Athanassiou, 2008), debido a que las plantas poseen vías metabólicas variadas y, por consiguiente, tienen requerimientos abióticos específicos para su desarrollo (Koch, 1998). La inferencia de la dieta de individuos de M. columbi de las localidades estadounidenses y mexicanas ha permitido tener una idea de las condiciones paleoambientales en los que estos vivían. La información disponible indica que M. columbi en Estados Unidos de América y México habitaba zonas abiertas con abundancia de plantas herbáceas, en los que había presencia de árboles bajos, donde además predominaban climas secos y fríos. En consecuencia, con base en la determinación de la dieta de los ejemplares CRINAH-2274 y CRINAH-2275, es posible inferir que durante el Pleistoceno en las localidades Barranca el Morillo y Sierra de Amozoc, existieron zonas abiertas con pastos. Este paleoambiente podría ser equiparado al existente durante el Pleistoceno en la Cuenca de Valsequillo, Puebla, zona que se ubica a tan sólo 78 km de barranca El Morillo y a 22 km de Sierra de Amozoc. La reconstrucción del paleoambiente de la Cuenca de Valsequillo indica predominio de vegetación abierta, pero con presencia de coníferas (Pinus, Quercus, Alnus), donde predominaba un clima de baja humedad, que alternaba episodios secos y fríos (Stevens et al., 2012; Pérez-Crespo et al., 2014; Metcalfe et al. ̧ 2016). Por consiguiente, es posible que este tipo de hábitat fuera común en el centro del estado de Puebla durante el Pleistoceno, favoreciendo el desarrollo de las poblaciones de M. columbi.
6. Conclusiones
La descripción del material de proboscídeos catalogados como CRINAH-2274 y CRINAH-2275, procedente de las localidades barranca El Morillo y Sierra de Amozoc, permitió la identificación del mamut Mammuthus columbi, adicionando así nuevos registros de esta especie en México. Asimismo, pudo establecerse que ambos ejemplares eran adultos sexualmente maduros. Únicamente se determinó el sexo para el individuo CRINAH-2274, basado en el diámetro del alveolo del incisivo, cuyas dimensiones son similares a las de ejemplares de Estados Unidos de América, que permitió referirlo como un macho.
La determinación de la edad y sexo en M. columbi se determinó por primera vez para ejemplares procedentes del estado de Puebla. Con esto se contribuye al conocimiento de algunos aspectos paleobiológicos de esta especie en México, que sin duda servirán para entender mejor la dinámica poblacional de M. columbi en América del Norte. Por último, se aplicó por primera vez en México el análisis de ángulos de mesodesgaste. Con este método se determinó que la dieta de los ejemplares CRINAH-2274 y CRINAH-2275, fue estrictamente pacedora. Lo anterior, a su vez, permite inferir que el hábitat de ambos individuos fue un área abierta con abundancia de pastos. Esto permite tener un esbozo general sobre las condiciones paleoambientales ocurridas en la región centro-este del estado de Puebla durante el Pleistoceno.

