











 nueva página del texto (beta)
nueva página del texto (beta) Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
Permalink

Introducción
Estudios recientes en México indican que, el factor de mayor influencia en la degradación de los ecosistemas y la biodiversidad, es el cambio del uso de la tierra (Sarukhán y col., 2015). De igual forma, la destrucción del hábitat y la fragmentación han tenido un gran impacto en los ecosistemas terrestres y acuáticos (Williams-Linera, 2002; Santos y Tallería, 2006; Rodríguez-Estrella y col., 2016). Por ejemplo, en 1976, la cobertura original de la vegetación primaria terrestre en el país se redujo en un 54 %, mientras que para 2011, la extensión original de la vegetación cubierta, en su condición primaria y secundaria, se menguó a 72 % según información del Instituto Nacional de Estadística y Geografía (INEGI, 2013). Sin embargo, recientemente, las tasas de deforestación han disminuido en muchas regiones del país, en parte debido a que las áreas restantes son inaccesibles, consideradas de bajo valor comercial o bien se encuentran bajo algún régimen de protección (Rosete-Vergés y col., 2014; FAO, 2016). Más allá de esto, actualmente, una parte significativa de la vegetación restante se compone de pequeños fragmentos aislados o está representada por el crecimiento secundario, es decir, vegetación en proceso de regeneración, después de ser sometida a perturbación de acuerdo a la Comisión Nacional para el Conocimiento y Uso de la Biodiversidad (CONABIO, 2009).
Un factor importante en el cambio del uso del suelo es la actividad pecuaria (Arriaga-Cabrera, 2009). De las 1 960 189 ha del territorio continental, a nivel nacional, aproximadamente 81 758 631 ha se identifican con uso de agostadero, enmontado o de otro tipo (INEGI, 2017). Esta situación se refleja en el norte de México, con el constante incremento de las actividades pecuarias y agrícolas (Návar-Chaidez, 2008; Cantú-Silva y col., 2017). Dichas prácticas han ocasionado cambios en la estructura del ecosistema y en su diversidad florística, con la consecuente pérdida de especies y material genético, invasión de especies generalistas y la alteración de las funciones, como la polinización y dispersión de semillas, entre otros (Jiménez-Pérez y col., 2012; Moreno-Medina y col., 2017).
Actualmente se busca integrar el uso y la conservación de los recursos naturales con las necesidades productivas de la población (Maass y col., 2005; Jose, 2012; SAGARPA, 2013). Desde hace 30 años, en el sector agropecuario, han surgido prácticas, en las que se integra y combina el manejo de árboles o arbustos con ganado (Musálem, 2002; Sánchez y col., 2003). Constituyen actividades agroforestales, que se han empezado a utilizar por ganaderos, con el fin de incrementar la productividad de sus tierras y promover la conservación de los recursos naturales (CONAFOR, 2010; SAGARPA, 2013).
Los sistemas agroforestales brindan beneficios múltiples, como la dispersión de especies vegetales y la conservación de animales silvestres y el paisaje conjunto (Mahecha y Zoot, 2002; Barrance y col., 2009). En términos de productividad, se ha descrito que los árboles presentes en ellos, además de proveer madera para cercas y combustible, generan un microclima mediante la sombra, que el ganado aprovecha (Mitlohner y col., 2002). Estas prácticas han sido tema de estudio, principalmente en los trópicos y ecosistemas templados del continente y del país, enfocándose primordialmente a la productividad de los mismos (Sánchez y col., 2003; Ramírez y col., 2012; Rosales-Adame y col., 2014). Aunque, se ha preconcebido la importancia y el potencial ecológico que sostienen las especies arbóreas que los integran, la evidencia científica que comprueba estas afirmaciones es aún escasa, en particular en las comunidades vegetales áridas y semiáridas.
El matorral espinoso tamaulipeco (MET) es una comunidad vegetal semiárida, caracterizada por la presencia de una alta diversidad de plantas, con elevados valores de densidad en su estrato arbóreo-arbustivo (Cram y col., 2006; Alanís-Rodríguez y col., 2008; González-Rodríguez y col., 2010; Alanís-Rodríguez y col., 2015; Pequeño-Ledezma y col., 2017). Sin embargo, es una de las comunidades en donde la ganadería extensiva ha sido practicada durante los últimos 350 años (Foroughbakhch y col., 2009), siendo esta una de las causas principales de su deforestación (Arriaga-Cabrera, 2009; Alanís-Rodríguez y col., 2015), por lo que resulta importante encontrar y aplicar nuevas prácticas de aprovechamiento que conserven las características mencionadas. En el MET se han reportado aproximadamente 80 especies maderables, de las cuales, 9 han sido destacadas con potencial agroforestal (Pando-Moreno y Villalón-Mendoza, 2001).
El objetivo de esta investigación fue evaluar algunos parámetros dasométricos y de diversidad de los elementos leñosos presentes en un matorral espinoso tamaulipeco (MET), sometido a la actividad pecuaria en el estado de Nuevo León, ubicado en la región noreste de México, para determinar si las modificaciones realizadas para este uso, contribuyen o no, a la conservación de la diversidad vegetal, con base en estudios recientes del MET.
Materiales y métodos
Área de estudio
El área se ubica en el ejido Emiliano Zapata, municipio de Linares, Nuevo León, en el noreste de México (Figura 1). Las coordenadas registradas están dentro del rango de los 24°54’ N y los 99°35’ W, en el suroeste de la región fisiográfica Llanura Costera del Golfo Norte (INEGI, 1986). El clima dominante en esta región es semicálido subhúmedo, con precipitación mayor a los 800 mm. El sistema de topoformas pertenece a lomerío suave con bajada, con predominancia de suelos regosoles. Las posibilidades de uso pecuario son de pastoreo extensivo sobre vegetación natural distinta del pastizal (INEGI, 1986).
La vegetación principal es el MET, en el cual predominan las asociaciones de estratos arbustivos altos o medianos y arbóreos altos, caracterizados en su mayoría por especies espinosas, con hojas compuestas (García-Hernández y Jurado, 2008; Alanís-Rodríguez y col., 2013). Las especies más representativas son Castela texana (Torr. & A. Gray) y Rose, Celtis pallida Torr., Cordia boissieri A. DC., Ebenopsis ebano (Berland.) Barneby & J.W. Grimes, Parkinsonia texana var. macra (I.M. Johnst.) Isely, Prosopis laevigata (Humb. & Bonpl. ex Wild.) M.C. Johnst., Randia obcordata S. Watson, Sideroxylon celastrinum (Kunth) T.D. Penn. y Vachellia rigidula (Benth.) Seigler & Ebinger, entre otras (Alanís-Flores, 1996; Jiménez-Pérez y col., 2013).
Análisis de la vegetación
En una superficie de 19 249 m2 se realizaron aclareos por lo bajo en los años 2011 y 2012, eliminando la mayoría de los individuos con d < 5 cm. La intención de este manejo fue modificar la estructura de la comunidad vegetal existente, creando espacios abiertos para el establecimiento de pastos y permitir el desplazamiento del ganado. Sin embargo, se conservan los árboles y arbustos de diámetros mayores. Las copas de los árboles brindan sombra que favorece el resguardo del ganado en temporadas de calor y proporcionan una fuente de alimento para el ganado y las aves, en variadas épocas del año.
En junio del año 2013, se realizó la evaluación de la comunidad vegetal, mediante el establecimiento de 4 sitios de muestreo de 1 600 m2 cada uno (33 % del área de estudio), distribuidos al azar. En los sitios de muestreo se evaluaron todas las especies arbóreas y arbustivas. Se registró la nomenclatura taxonómica de cada individuo y se realizaron mediciones de altura total (h), diámetro basal (d0.5) y cobertura de copa, en sentido nortesur y este-oeste.
El d0.5 se midió a partir de las bifurcaciones que presentaron los individuos a mayor altura, ya que resulta más práctico y económico realizar una sola medición (Alanís-Rodríguez y col., 2008).
Análisis de la información
Para evaluar el significado ecológico de las especies arbóreas se estimaron los indicadores relativos de densidad (ARi), dominancia (DRi), frecuencia (FRi ) e índice de valor de importancia (IVI) (Müeller-Dombois y Ellenberg, 1974; Magurran, 2004), calculados a partir de las siguientes ecuaciones. La densidad relativa se obtuvo por medio de la densidad absoluta, y el cálculo de su valor relativo. Se utilizaron las siguientes ecuaciones:
Donde: Ai es la densidad absoluta de la especie Ai, Ni es el número de individuos de la especie i, y S la superficie muestral en ha.
Donde: ARi es la densidad relativa de la especie i, Ai con respecto a la densidad total ∑ Ai.
Para estimar la dominancia relativa también se empleó el uso de dos ecuaciones, la dominancia absoluta y el cálculo de su valor relativo. Se utilizaron las siguientes ecuaciones:
Donde: Di es la dominancia absoluta de la especie i, Abi el área basal de especie i, y S la superficie muestral (ha).
Donde: DRi es la dominancia relativa de la especie i, Di con respecto a la dominancia total ∑ Di.
La frecuencia relativa se obtuvo calculando la frecuencia absoluta y posteriormente su valor relativo. Se utilizaron las siguientes ecuaciones:
Donde: Fi es la frecuencia absoluta de la especie i, Pi es la frecuencia de la especie i en los sitios de muestreo, y NS el número total de sitios de muestreo.
Donde: FRi es la frecuencia relativa de la especie i, con respecto a la frecuencia total ∑Fi.
El índice de valor de importancia (IVI) se define como:
Donde: ARi es la densidad relativa de la especie i, DRi es la dominancia relativa de la especie i, y FRi es la frecuencia relativa de la especie i.
Para estimar la diversidad alfa y la riqueza de especies se utilizaron, el índice de ShannonWiener (H´) (Shannon y Weaver, 1949) y el índice de Margalef (Clifford y Stephenson, 1975), respectivamente. El índice de Shannon-Wiener se estimó mediante la siguiente ecuación:
Donde: S es el número de especies presentes, ln es logaritmo natural y pi es la proporción de las especies, la cual se calcula con la siguiente ecuación:
Donde: Ni es el número de individuos de la especie i y N es el número total de individuos.
Con el mismo significado de las variables en común, el índice de diversidad de Margalef (DMg) se estimó con la ecuación:
La distribución de las especies en tres diferentes zonas de altura se calculó mediante el índice de distribución vertical de Pretzsch (Pretzsch, 1996), el cual toma tres zonas de altura: zona I: 80 a 100 % de la altura máxima de la población; zona II: 50 % a 80 %; zona III: 0 % a 50 %.
Posteriormente, se calculó el índice de distribución vertical de especies (A), el cual cuantifica la diversidad de especies y su ocupación en el espacio vertical en la población (Estrada-Castillón y col., 2005). El índice de distribución vertical se calculó de acuerdo a la siguiente fórmula:
Donde: S es el número de especies presentes; Z el número de zonas de altura y pij la proporción de especies en cada zona de altura, la cual se calcula con la siguiente ecuación:
Donde: nij es el número de individuos de la misma especie (i) en la zona (j) y N el número total de individuos.
Del índice A se derivan el Amax, que corresponde al valor máximo de A, dado por el número de especies (S) y zonas de altura (Z); y el Arel, que es la estandarización en porcentaje del índice A. Estos son indicativos de la distribución de las especies en los estratos de altura, de manera que, entre más cercano sea el valor de Amax al índice de A, más se acerca el valor de Arel al 100 %, lo cual se presenta cuando el número total de individuos se encuentra equitativamente distribuido entre estratos y es pecies (Rubio-Camacho y col., 2014).
Donde: ln es el logaritmo natural, S el número de especies y Z zonas de altura.
Para describir la estructura de la comunidad en términos de la densidad de cada especie, se utilizó un modelo que describe la relación gráfica entre el valor de la densidad (en una escala logarítmica) y las especies, en función de un arreglo secuencial de la más a la menos abundante (Martella y col., 2012).
Resultados
Se registraron 17 especies arbóreas y arbustivas, distribuidas en 16 géneros y 8 familias de plantas vasculares (Tabla 1). La familia con mayor riqueza específica fue Fabaceae, con 8 especies.
Tabla 1 Nombre científico y común, familia y forma biológica de las especies arbustivas y arbóreas registradas en un área de matorral espinoso tamaulipeco con actividad pecuaria en el noreste de México.
Table 1. Common and scientific name, family and life form of the tree and shrub species recorded in a Tamaulipas thornscrub area with livestock activity in Northeastern, Mexico.
| Nombre científico | Nombre común | Familia | Forma biológica* |
|---|---|---|---|
| Celtis pallida Torr. | Granjeno | Ulmaceae | Arbustiva |
| Condalia hookeri M. C. Jhonst. | Brasil | Rahmnaceae | Arbórea |
| Cordia boissieri A. DC. | Anacahuita | Boraginaceae | Arbustiva |
| Diospyros texana Scheele | Chapote | Ebenaceae | Arbórea |
| Ebenopsis ebano (Berland.) Barneby & J. W. Grimes | Ébano | Fabaceae | Arbórea |
| Ehretia anacua (Berland.) Barneby & J. W. Grimes | Anacua | Boraginaceae | Arbórea |
| Eysenhardtia polystachya (Ortega) Sarg. | Vara dulce | Fabaceae | Arbórea |
| Havardia pallens (Benth.) Britton & Rose | Tenaza | Fabaceae | Arbustiva |
| Helietta parvifolia Benth. | Barreta | Rutaceae | Arbórea |
| Parkinsonia texana var. macra (I. M. Johnst.) Isely | Palo verde | Fabaceae | Arbustiva |
| Porlieria angustifolia A. Gray | Guayacán | Zygophyllaceae | Arbustiva |
| Prosopis laevigata (Humb. & Bonpl. ex Wild.) M. C. Johnst. | Mezquite | Fabaceae | Arbórea |
| Senegalia greggii Britton. & Rose | Uña de gato | Fabaceae | Arbustiva |
| Sideroxylon celastrinum (Kunth) T.D. Penn. | Coma | Sapotaceae | Arbórea |
| Vachellia farnesiana (L.) Wight & Arn. | Huizache | Fabaceae | Arbustiva |
| Vachellia rigidula (Benth.) Seigler & Ebinger | Gavia | Fabaceae | Arbustiva |
| Zanthoxylum fagara (L.) Sarg. | Colima | Rutaceae | Arbustiva |
*Especies con un solo fuste y diámetro promedio mayor a 10 cm se consideraron arbóreas.
Parámetros ecológicos
Densidad
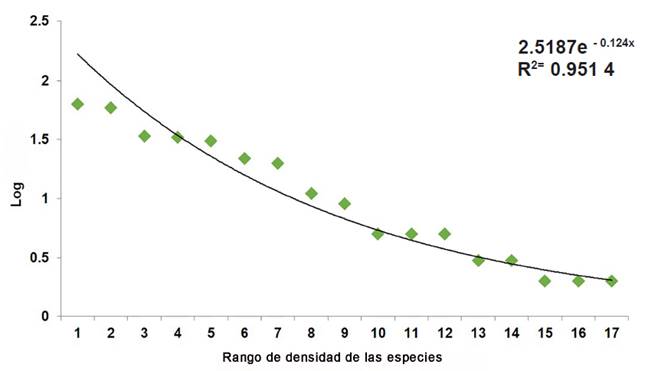
La densidad absoluta, para el total del área de estudio, fue de 309 individuos/ha. Las 3 especies más abundantes fueron Prosopis leavigata (Humb. & Bonpl. ex Wild.) M. C. Johnst., Havardia pallens (Benth.) Britton & Rose y Celtis pallida Torr. (Tabla 2). La forma de la curva de la densidad de las especies (en una escala logarítmica) fue una Log-normal típica, con una línea de tendencia exponencial, la cual es muy útil cuando los valores de los datos aumentan o disminuyen a intervalos cada vez mayores (Figura 2). Se observa que el valor R2 es 0.951 4, cercano a 1, lo cual indica un buen ajuste de la curva a los datos.
Tabla 2 Densidad, dominancia, frecuencia e índice de valor de importancia (IVI) de las especies arbustivas y arbóreas registradas en un área de matorral espinoso tamaulipeco con actividad pecuaria en el noreste de México (ordenadas de acuerdo al IVI).
Table 2. Density, dominance, frequency and importance value index (IVI) of the tree and shrub species recorded in a Tamaulipan thornscrub area with livestock activity in Northeastern Mexico (ranked by IVI).
| Nombre cientifico | Densidad absoluta (Ni/ha) | Densidad relativa (%) | Densidad absouluta(m2 /ha) | Densidad relativa (%) | Frecuencia relativa(%) | IVI(%) |
|---|---|---|---|---|---|---|
| Prosopis laevigata | 63 | 20.39 | 5 341.21 | 56.09 | 9.30 | 28.59 |
| Havardia pallens | 59 | 19.09 | 1 128.20 | 11.85 | 9.30 | 13.41 |
| Cordia boissieri | 33 | 10.68 | 639.22 | 6.71 | 9.30 | 8.89 |
| Celtis pallida | 34 | 11.00 | 382.72 | 4.02 | 9.30 | 8.11 |
| Condalia hookeri | 31 | 10.03 | 439.95 | 4.62 | 6.98 | 7.21 |
| Vachellia rigidula | 22 | 7.12 | 442.90 | 4.65 | 6.98 | 6.25 |
| Ehretia anacua | 9 | 2.91 | 142.58 | 1.50 | 9.30 | 4.57 |
| Sideroxylon celastrinum | 20 | 6.47 | 195.20 | 2.05 | 4.64 | 4.39 |
| Senegalia greggii | 11 | 3.56 | 246.91 | 2.59 | 6.98 | 4.38 |
| Vachellia farnesiana | 5 | 1.62 | 168.24 | 1.77 | 4.65 | 2.68 |
| Ebenopsis ebano | 3 | 0.97 | 154.73 | 1.63 | 4.65 | 2.42 |
| Porlieria angustifolia | 5 | 1.62 | 34.90 | 0.37 | 4.65 | 2.21 |
| Zanthoxylum fagara | 3 | 0.97 | 41.23 | 0.43 | 4.65 | 2.02 |
| Parkinsonia texana var. macra | 5 | 1.62 | 118.22 | 1.24 | 2.33 | 1.73 |
| Eysenhardtia polystachya | 2 | 0.65 | 27.62 | 0.29 | 2.33 | 1.09 |
| Diospyros texana | 2 | 0.65 | 14.98 | 0.16 | 2.33 | 1.05 |
| Helietta parvifolia | 2 | 0.65 | 3.02 | 0.03 | 2.33 | 1.00 |
| Total | 309 | 100.00 | 9 521.83 | 100.00 | 100.00 | 100.00 |
Dominancia
La cobertura de copa en el área fue de 9 521.83 m2/ha, lo que indica que, en general, existe un 95 % de la superficie cubierta por el dosel, y únicamente el 5 % se encuentra desprovista (asumiendo mínimo traslape de copas). Prosopis leavigata fue la especie dominante, con 56.09 % de la cobertura de copa de todas las especies en el área de estudio. La segunda especie en dominancia fue Havardia pallens, seguida de Cordia boissieri.
Frecuencia
Se registró la presencia de 5 especies en todos los sitios de muestreo, Celtis pallida, Cordia boissieri, Ehretia anacua (Berland.) Barneby & J. W. Grimes, Harvardia pallens y Prosopis leavigata, cada una con 9.3 % de frecuencia relativa (Tabla 2). Le siguen Condalia hookeri M. C. Jhonst., Vachellia rigidula y Senegalia greggii Britton. & Rose, con presencia en cuatro de los 5 sitios de muestreo.
Índice de valor de importancia (IVI)
La especie con mayor peso ecológico, según el índice de valor de importancia fue Prosopis leavigata (28.59 %), le siguen Havardia pallens y Cordia boissieri. Las especies con menor peso ecológico fueron Helietta parvifolia Benth. y Diospyros texana Scheele (Tabla 2).
Distribución vertical de las especies
Las especies del estrato I registraron una densidad de 16 individuos/ha, del estrato II de 208 individuos/ha y del estrato III de 85 individuos/ha; lo que indicó que la comunidad vegetal era cohortal, es decir, que predominaba un solo estrato (II) (Figura 3). De acuerdo al índice de distribución vertical de especies (A), el estrato I (alto, de 9.85 m a 12.3 m) se encontraba conformado por las especies Prosopis leavigata, Havardia pallens y Vachellia farnesiana (L.) Wight & Arn. En el estrato II (medio, de 6.16 m a 9.84 m), que fue el mejor representado, más del 50 % de las especies fueron Havardia pallens, Prosopis laevigata, Condalia hookeri y Vachellia rigidula, entre otras, con menor presencia. Havardia pallens fue la especie predominante en este estrato. Por otro lado, el estrato III (bajo, menor a 6.16 m) se encontraba conformado casi en un 60 % por Cordia boissieri, Celtis pallida y Sideroxylon celastrinum, entre otras, con menor presencia; Sideroxylon celastrinum alcanzó la altura máxima de 6.14 m.
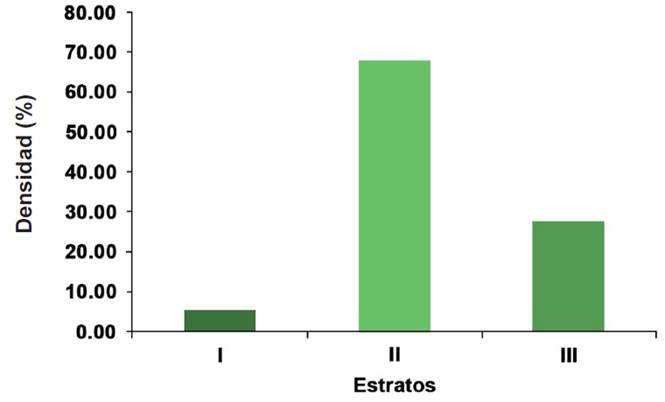
El índice de distribución vertical (A) fue de 2.88, con un Amax de 3.93 y un Arel de 73 %, lo que indica uniformidad alta, en cuanto a la diversidad de altura.
Densidad de los individuos por clase diamétrica
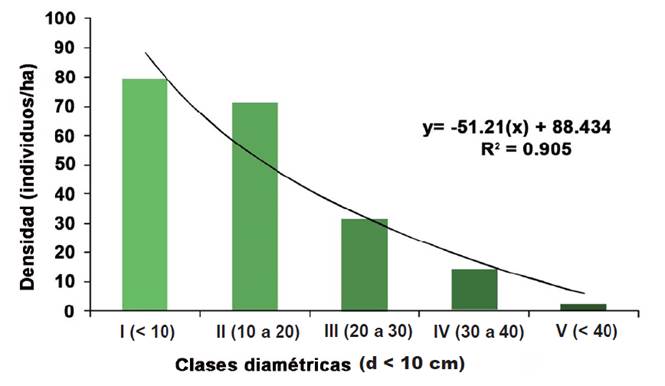
Al analizar la densidad de individuos por ha, de acuerdo a las clases diamétricas, mediante un gráfico de barras, se observó una línea logarítmica, con tendencia negativa en la densidad de individuos, conforme aumenta el diámetro de los mismos (Figura 4). Las densidades mayores de individuos ( ≥ 60 individuos/ha), se concentraron en las clases diamétricas I y II.
Discusión
La familia con mayor número de especies fue Fabaceae, lo cual coincide con estudios recientes realizados en el MET, bajo diversas condiciones (Estrada-Castillón y col., 2005; González-Rodríguez y col., 2010; Jiménez-Pérez y col., 2012; Molina y col., 2013; Mora-Donjuán y col., 2013a; 2013b; Pequeño-Ledezma y col., 2017). Comparando con otras evaluaciones del MET, en donde se aplicó la misma metodología (igual número y dimensiones de los sitios de muestreo), la riqueza específica registrada fue mayor, que en áreas con regeneración pasiva e historial de pastoreo intensivo (Pequeño-Ledezma y col., 2012; Yerena-Yamallel y col., 2014), pero menor que en áreas sin disturbio reciente (Alanís-Rodríguez y col., 2011; Mora-Donjuán y col., 2013a; Yerena-Yamallel y col., 2014).
Parámetros ecológicos
Densidad
Havardia pallens, la segunda especie de mayor densidad relativa, en este estudio, ha sido reportada como una de las tres especies con mayor densidad relativa en áreas de MET sin disturbio (García-Hernández y Jurado, 2008; Yerena-Yamallel y col., 2014) y en terrenos con historial de ganadería extensiva (Pequeño-Ledezma y col., 2012). Por el contrario, Prosopis leavigata y Celtis pallida, las cuales fueron también dos de las especies más abundantes en este trabajo, presentaron menor densidad en los estudios referidos, así como en el trabajo de Jiménez-Pérez y col. (2013), quienes analizaron áreas del MET con diferentes tipos de alteraciones.
La forma de la curva de densidad estuvo determinada por la densidad por especie, la cual sigue una distribución normal (Figura 2), con muchas especies con valores intermedios y pocas con valores mínimos o máximos de densidad. Este tipo de distribución se encuentra en la mayoría de las comunidades vegetales (Martella y col., 2012), como el MET en el presente estudio, donde las especies Prosopis leavigata y Havardia pallens fueron las que dominaron con valores máximos de densidad, mientras que Eysenhardtia polystachya (Ortega) Sarg., Diospyros texana y Helietta parvifolia registraron los valores mínimos (Tabla 2).
Dominancia
Frecuentemente, las especies reportadas como más dominantes en el MET, con historial pecuario y agrícola, son Vachellia farnesiana y Vachellia rigidula (Alanís-Rodríguez y col., 2008; Pequeño-Ledezma y col., 2012; JiménezPérez y col., 2013; Mora-Donjuán y col., 2013b); Diferente a lo que reportaron Mora-Donjuán y col. (2013a; 2013b) en un área sin disturbio reciente, donde las especies dominantes fueron Vachellia amentacea, Diospyros texana y Cordia boissieri, con esta última presente en el sistema pastoreado. La suma total de la dominancia absoluta de las especies en el área de estudio (Tabla 2) es mayor a los registros de dominancia absoluta documentadas por Mora-Donjuán y col. (2013b), en sus tres áreas de estudio del MET (dos con historial pecuario y una sin disturbio).
Frecuencia
Los resultados del presente estudio, para este parámetro, difieren de los reportados por Alanís-Rodríguez y col. (2008); Jiménez-Pérez y col. (2012) y Pequeño-Ledezma y col. (2012), quienes evaluaron áreas del MET con algún grado de disturbio, ya que registraron no más de tres especies presentes en todos los sitios de muestreo. Sin embargo, los resultados de este trabajo, concuerdan con los de Mora-Donjuán y col. (2013a) , estudio de un área sin alteración reciente, donde se registraron que siete especies se encontraban en todos los sitios de muestreo. Lo anterior, prueba que, conforme más especies se encuentren en todos los sitios de muestreo, menos disturbio muestra el área evaluada, ya que, al no haber afectaciones graves en la comunidad vegetal, más especies tienen posibilidad de establecerse en diversos sitios, evitando que predominen un número reducido de especies en toda el área.
Índice de valor de importancia (IVI)
En el presente estudio, Prosopis leavigata fue la especie con mayor IVI (28.59 %), con una diferencia porcentual de 15.18 %, respecto a la siguiente especie Havardia pallens. Este estudio es el segundo que registra a Prosopis leavigata como la especie con el mayor IVI en el MET con uso ganadero (Alanís-Rodríguez y col., 2008; Jiménez-Pérez y col., 2012; Pequeño-Ledezma y col., 2012; Mora-Donjuán y col., 2013a; 2013b).
Prosopis leavigata presentó una densidad relativa (20.39 %) ligeramente superior a la de Havardia pallens (19.09 %). Sin embargo, su dominancia relativa (56.09) fue mucho mayor, con una diferencia de 44.24 % (Tabla 2). Esto indica una alta preferencia del manejador para conservar dicha especie, ya que, si se compara con otros estudios, que se han llevado a cabo para el MET, se observa que son otras las especies con mayor densidad, entre ellas, Diospyros texana y las del género Vachellia (Alanís-Rodríguez y col., 2008; Pequeño-Ledezma y col., 2012; Jiménez-Pérez y col., 2013; Mora-Donjuán y col., 2013a). Esta preferencia se adjudica al conocimiento sobre los múltiples usos del Prosopis leavigata (mejor conocido como mezquite) en la región, tales como leña, fuente de carbón, uso artesanal y de forraje (Estrada-Castillón y col., 2005), siendo el último de especial interés para la producción ganadera (Ríos-Saucedo y col., 2012).
Distribución vertical de las especies
Aunado a los resultados y siendo que el diámetro menor registrado fue de 4 cm (datos no mostrados), se tiene una comunidad vegetal que no se encontraba en regeneración, como otros matorrales (Alanís-Rodríguez y col., 2011; Jiménez-Pérez y col., 2012; MoraDonjuán y col., 2013b), pero que tampoco tiene las condiciones óptimas de un matorral primario (Mora-Donjuán y col., 2013b; Yerena-Yamallel y col., 2014); sino que sus componentes conforman una masa irregular joven e incoetánea, en la cual, los árboles crecen principalmente en diámetro (Araujo e Iturre, 2006).
Diversidad
El valor de DMg, registrado en este estudio, fue superior al evaluado por Alanís-Rodríguez y col. (2008), en un área con historial de uso de ganadería intensiva y extensiva. También el índice de diversidad de Shannon-Wiener (H´), reportado en este trabajo, fue superior a los obtenidos en áreas con diferente historial de uso productivo (Alanís-Rodríguez y col., 2008; Jiménez-Pérez y col., 2012; Pequeño-Ledezma y col., 2012; Mora-Donjuán y col., 2013b; Molina-Guerra y col., 2013) e incluso comparado con áreas de MET sin alteración reciente (Mora-Donjuán y col., 2013a; 2013b). Una explicación de que los valores reportados en el estudio fueron superiores, con respecto a los reportados en las investigaciones mencionadas, sería que la comunidad evaluada se encontraba en un estado joven, donde la constante selección de especies crea nuevos espacios, que permiten que se establezcan otras especies o nuevos individuos.
Conclusiones
La composición de especies arbóreas y arbustivas en el área de estudio conserva atributos cercanos a las comunidades de matorral espinoso tamaulipeco (MET) sin alteraciones, como la riqueza de especies, diversidad, densidad y área de copa. Sin embargo, en el manejo de selección de las especies destaca la preferencia por conservar individuos de altura distribuidas en el estrato II (6.16 m a 9.84 m), con diámetros de 10-20 cm, características de una comunidad vegetal irregular e incoetánea. La especie de preferencia fue Prosopis laevigata, deseable por ser multipropósito (fuente de leña y carbón, uso artesanal y forrajera). Se considera importante aumentar la investigación de los aprovechamientos mixtos en el noreste de México.

