











 text new page (beta)
text new page (beta) Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO 
 Permalink
PermalinkINTRODUCCIÓN
La Reserva de la Biósfera Los Petenes, en el estado mexicano de Campeche, es un área natural protegida, que integra tanto ambientes continentales, como marinos de gran relevancia ecológica. En la porción continental, los petenes son ecosistemas particulares que solo se encuentran en las penínsulas de Yucatán, México, la Florida, Estados Unidos de Norteamérica y en la isla de Cuba. Son islotes de vegetación asociada a cuerpos de agua formados por afloramientos de ríos subterráneos. En su porción marina, la RBLP cuenta con la más grande y mejor conservada superficie de pastos marinos del país, dominada por Thalassia testudinum, lo que la convierte en un hábitat propicio para fines de protección, alimentación, crianza y desove de diferentes especies entre de los que se encuentran recursos pesqueros de gran valor económico, como lisas, corvinas, pargos, huachinangos y cazón, entre otros (Villalobos-Zapata, 2004; CONANP, 2006; Muñoz-Rojas y col., 2013).
La distribución y abundancia de los recursos bióticos de la RBLP, particularmente de los peces, no es casual y responde a lo que en la literatura se ha descrito como programación espacial y temporal acoplada a la variabilidad ambiental (Lara-Domínguez y Yáñez-Arancibia, 1999). Este acoplamiento a las condiciones del hábitat para el desarrollo de distintas etapas del ciclo de vida de múltiples especies refleja altos niveles de exergía y se logra por el tipo de asociaciones que las especies establecen (Lugendo y col., 2007). De acuerdo con Arceo-Carranza y col. (2013), la variabilidad ambiental condiciona la formación de asociaciones de peces que tanto en escala espacial como temporal constituyen un mecanismo para disminuir la competencia inter e intraespecífica y aprovechar las oportunidades de alimentación que el hábitat les ofrece. Los estudios sobre las variaciones nictemerales de la estructura de las comunidades de peces en sistemas costeros tropicales son escasos y particularmente para la RBLP, por lo que analizar las asociaciones de peces dominantes en un mareo de variabilidad ambiental temporal (secas y lluvias) y nictemeral (día y noche), fue el objetivo del presente estudio y se considera una aportación de información de utilidad para acciones regulatorias y de conservación.
MATERIALES Y MÉTODOS
El área de estudio se localiza en la costa norte del estado de Campeche, en el sureste del Golfo de México (Figura 1). La Reserva de la Biósfera Los Petenes tiene una extensión de 282 857 ha y cuenta con la mayor superficie de protección en su zona marina. Su límite marino considera hasta las 12 millas náuticas (Villalobos-Zapata, 2004). El clima predominante en la zona centro-sur de la RBLP es Aw (cálido subhúmedo con lluvias en verano), mientras que en su extremo norte es del tipo BS'h'w (semiseco y seco-cálido). Se identifican dos épocas climáticas: época de secas que abarca de noviembre a abril y lluvias de mayo a octubre (CONANP, 2006).
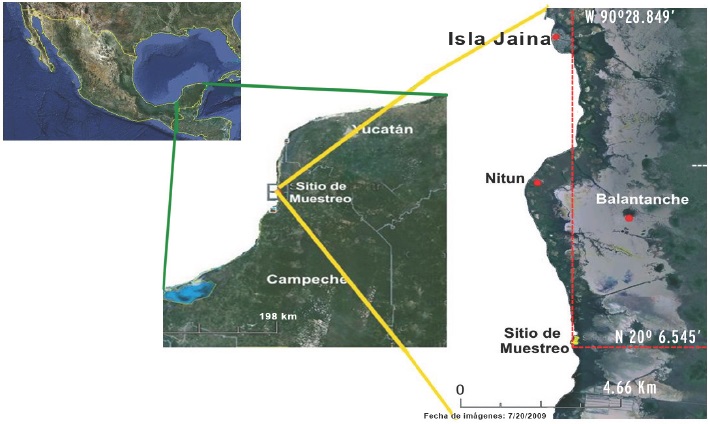
El sitio de muestro se ubica cerca de Isla Jaina (20°6.545' N y 90°28.849' W), en la región central de la porción marina de la RBLP. El sitio fue seleccionado a partir de muestreos previos, que destacaron la cobertura de pastos marinos y la gcomorfologia de la zona, que incluye la convergencia de al menos dos esteros que penetran al continente varios centenares de metros y que en la bajamar arrastran aportes continentales de materia orgánica.
Las campañas se realizaron bimestralmente entre diciembre 2010 a octubre 2011, recolectando muestras biológicas cada 2 h en un período de 24 h, utilizando una red de arrastre de 5 m de largo, 2.5 m de abertura de trabajo y 19 mm de luz de malla, operada por 12 min a bordo de una laneha con motor fuera de borda, a una velocidad de 2.5 nudos, para cubrir una área de arrastre aproximada de 1 800 m2. Se realizaron 72 arrastres durante el período de estudio. El ciclo nictemeral se dividió en horas de día y horas de noche, considerando las 7 horas como la primera hora de día y las 19 horas como la primera hora de noche. Los organismos capturados se almacenaron en bolsas de plástico etiquetadas y se conservaron en hielo.
En el sitio de muestreo se realizaron mediciones de: temperatura (°C), salinidad (UPS, unidades prácticas de salinidad), oxígeno disuelto (mg / L) y pH del agua; tanto a nivel superficial como de fondo, empleando una sonda multiparamétrica Hydrolab DS5 (Loveland, USA). La medición de las variables de superficie se realizó aproximadamente a 30 cm de profundidad y las de fondo a 10 cm del piso.
En el laboratorio los organismos fueron identificados utilizando literatura especializada (Fischer, 1978; Cervigón y col., 1992; Castro-Aguirre, 1999). El ordenamiento sistemático de las especies se realizó de acuerdo con Nelson (1984). Los valores de longitud total, longitud estándar (mm) y peso total (g) fueron registrados para cada espécimen, utilizando un ictiómetro convencional y una balanza digital de 2 160 g y 0.1 g de precisión.
El comportamiento temporal y nictemeral de las variables ambientales se analizó usando el programa estadístico Systat Software Ine. (2009). Los resultados se representan mediante gráficos de caja, que expresan los valores mínimo y máximo, así como la mediana, y los cuartiles primero y tercero. Los valores atípicos leves se indican con pequeños círculos y los valores atípicos extremos con asteriscos. Los valores atípicos se definen como:
Atípico leve:
Atípieo extremo:
Donde: Q1 y Q3 corresponden a los valores de los cuartiles primero y tercero respectivamente. IQR corresponde al intervalo interquartílieo Q3 - Q1.
Para analizar y discutir los resultados se realizaron pruebas estadísticas para comparar los distintos ciclos de muestreo. En primer lugar se realizaron pruebas de normalidad a datos agrupados por ciclo de muestreo, lo que permitió decidir si se utilizaban estadísticos de prueba paramétricos o no paramétricos. Considerando que la mayor parte de los datos agrupados por ciclo de muestreo presentaron diferencias significativas respecto de la normalidad (P < 0.05), se decidió utilizar la prueba no paramétrica de Kolmogorov-Smirnov para realizar dichas comparaciones. Este procedimiento se realizó con ayuda del software StatPlus (AnalystSoft Ine., 2009).
Para determinar la dominancia de las especies se consideró la abundancia numérica y en peso y la frecuencia de aparición, de acuerdo con el índice de importancia relativa (IIR) propuesto por Koranteng (2001), y que se define como:
Donde:
% W = porcentaje de la contribución en peso de la especie a la captura total; % N = porcentaje de la contribución en número de la especie a la captura total; % F = Porcentaje de número de estaciones en las que se presenta la especie en relación con el número total de estaciones muestreadas.
Las especies con valores IIR ≥ 20 se consideraron especies dominantes, valores entre 20 y 1 se consideraron de importancia media y valores menores de 1 se consideraron de baja importancia.
Las asociaciones entre las especies dominantes se determinaron mediante un análisis elúster (método Ward, distancia gamma), de las matrices de abundancia numérica por especie por ciclo de muestreo, utilizando el software Systat (Systat Software Ine. 2009). Las diferencias observadas entre secas y lluvias, así como entre día y noche, se analizan y diseuten considerando la variabilidad ambiental y las caraeterísticas biológicas y ecológicas de las especies.
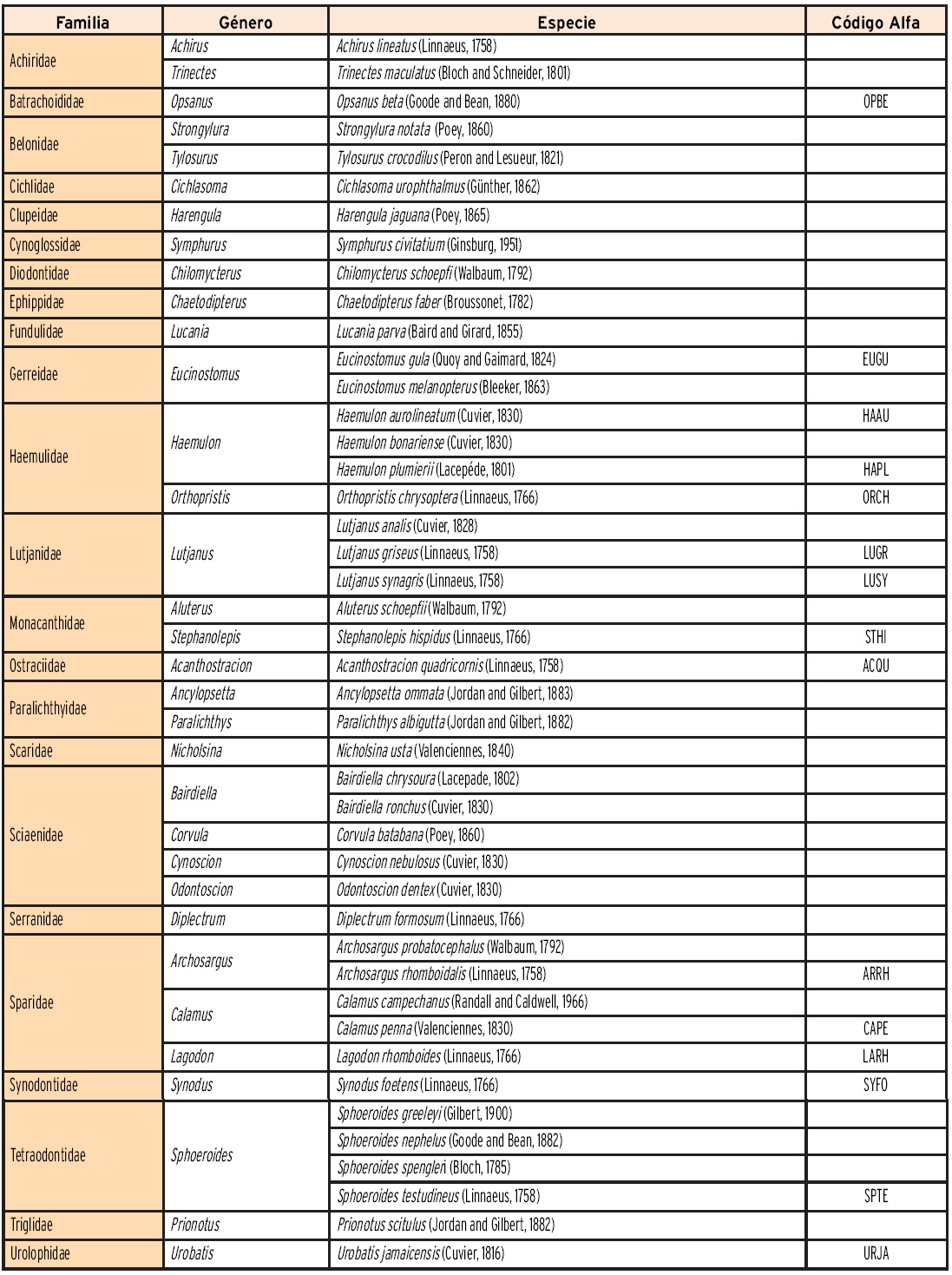
Las matrices de número de individuos por especie y valores de las variables ambientales organizadas por época climática y por horas de día y noche fueron sometidas a un Análisis Canónico de Correspondencias (ACC), con ayuda del programa computacional Multivariate Statistical Package (MVSP) (Kovach Computing Services, 2013), lo que permitió generar elementos de discusión sobre la influencia de las variables ambientales en la distribución y abundancia temporal y nictemeral de los peces en la zona de estudio. La representación gráfica de los clúster y de las correspondencias canónicas incluye el nombre de las especies utilizando el código alfa que integra las dos primeras letras del género y dos de la especie.
RESULTADOS
Los resultados obtenidos no mostraron diferencia significativa (P < 0.05), entre los valores registrados en superficie y fondo del sitio de muestreo para las variables de temperatura, salinidad, pH y oxígeno disuelto. Por lo cual la variabilidad ambiental temporal (Figura 2) y nictemeral (Figura 3), del sitio de estudio, se reporta en función de los valores promedio.
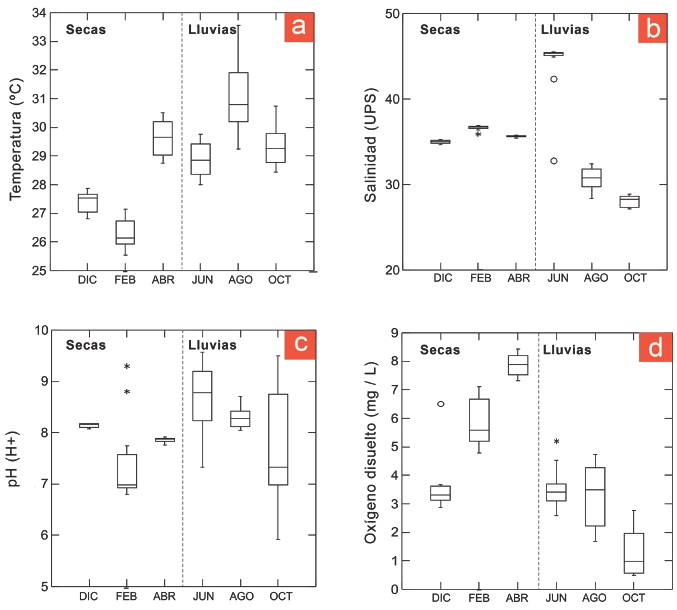
La variabilidad ambiental en la zona de estudio se representa mediante la Figura 2 que integra las mediciones de temperatura, salinidad, pH y oxígeno disuelto, agrupadas por mes y por época climática, y describen los valores de mediana, primero y tercer interquartil y valores mínimo y máximo. El valor mínimo de temperatura se registró en febrero (25.5 °C) y el máximo en agosto (33.5 °C), con el mayor intervalo de variación (Figura 2a). La salinidad mostró valores que oscilaron alrededor de los 35.75 UPS durante la época de secas, en tanto que durante la época de lluvias se registraron los valores mínimos en octubre (27.1 UPS) y máximos en junio (45.5 UPS), con dos valores atípicos extremos (Figura 2b). El pH del agua mostró la mayor variación durante la época de lluvias, con valores que oscilaron entre 5.9 y 9.57; los valores mínimos se observaron en febrero. Sin embargo, también en este mes se registraron dos valores atípicos leves (Figura 2e). Finalmente, el oxígeno disuelto mostró una tendencia de incremento durante la época de secas, en tanto que durante las lluvias la tendencia fue inversa. Los valores mínimos se presentaron en octubre (0.48 mg / L ) y los máximos en abril (8.43 mg / L) (Figura 2d).
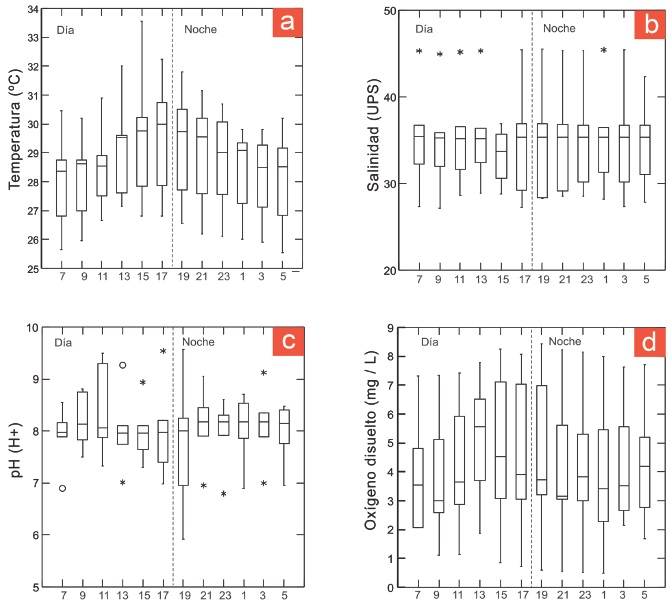
Durante los ciclos nictemerales, la temperatura alcanzó un promedio anual de 28.8 °C, con un intervalo de variación de 33.5 °C a las 15 horas y 25.5 °C a las 5 horas. Durante el día se distingue una tendencia de incremento y a partir de las 19 horas la temperatura disminuye paulatinamente (Figura 3a). La salinidad promedio para el período de estudio fue de 36.1 UPS con una variación entre 26.7 UPS y 45.5 UPS. Durante las horas de día se presentaron valores atípicos leves que correspondieron a las mediciones del mes de junio, cuando se registró la temperatura más elevada (Figura3b). El valor promedio del pH para el período de estudio fue de 8.03 con un intervalo de variación de 5.9 a 9.6. A las 13 horas se observaron valores atípicos leves y extremos, y a las 19 horas se presentó el intervalo de variación más amplio del período (Figura 3e). Finalmente, el oxígeno disuelto presentó un valor promedio de 4.51 mg / L durante todo el período de muestreo, con un intervalo de variación de 0.48 mg / L a 8.43 mg / L. Los intervalos de variación de los valores agrupados por horas de muestreo son amplios, ya que integran los registros de todo el período de muestreo (Figura 3d).
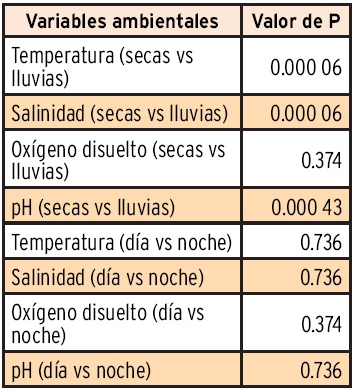
Los resultados de la prueba de normalidad W de Shapiro-Wilk, para las variables ambientales agrupadas por época climática y por día y noche, se muestran en la Tabla 1. La salinidad de lluvias y el oxígeno disuelto y pH de secas presentaron una distribución normal (P < 0.05), pero el resto tuvieron una diferencia significativa respecto de la normalidad (P < 0.05). En el caso de la agrupación por horas de día y de noche sólo la temperatura y pH de día y el oxígeno disuelto de noche rechazan la normalidad (P < 0.05). Con base en estos resultados se decidió dar un tratamiento no paramétrico a todos los datos agrupados para hacer las comparaciones entre ciclos nictemerales y por épocas climáticas.
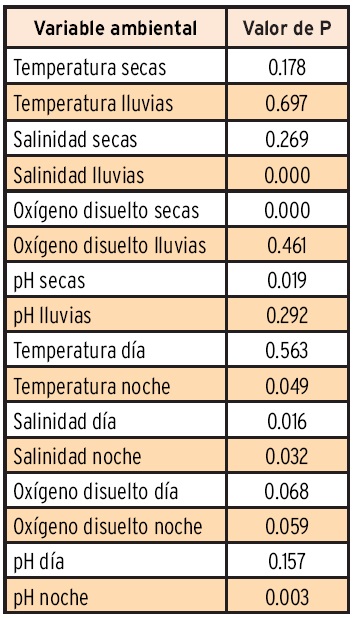
La comparación entre las épocas climáticas de secas y lluvias, mediante la prueba de diferencia máxima de Kolmogorov-Smirnov permitió encontrar diferencias significativas (P < 0.000 05) para la temperatura, salinidad y pH, sin embargo el oxígeno disuelto no presentó diferencias significativas (P < 0.37). En el mismo sentido, al comparar las horas de día con respecto a las horas de noche de los ciclos nictemerales no se encontraron diferencias significativas (P < 0.37). En la Tabla 2 se muestran los resultados de significancia de dichas comparaciones.
Se capturaron en total 10 602 peces con un peso global de 163.6 kg. El análisis de estos especímenes permitió identificar 44 especies, agrupadas en 33 géneros, pertenecientes a 23 familias. En la Tabla 3 se presenta el listado sistemático de las especies capturadas, se incluye el código alfa que identifica a las especies dominantes y se destaca la diversidad de las familias Sciaenidae y Sparidae con cinco especies cada una, Tetraodontidae con cuatro y Haemulidae y Lutjanidae con tres.
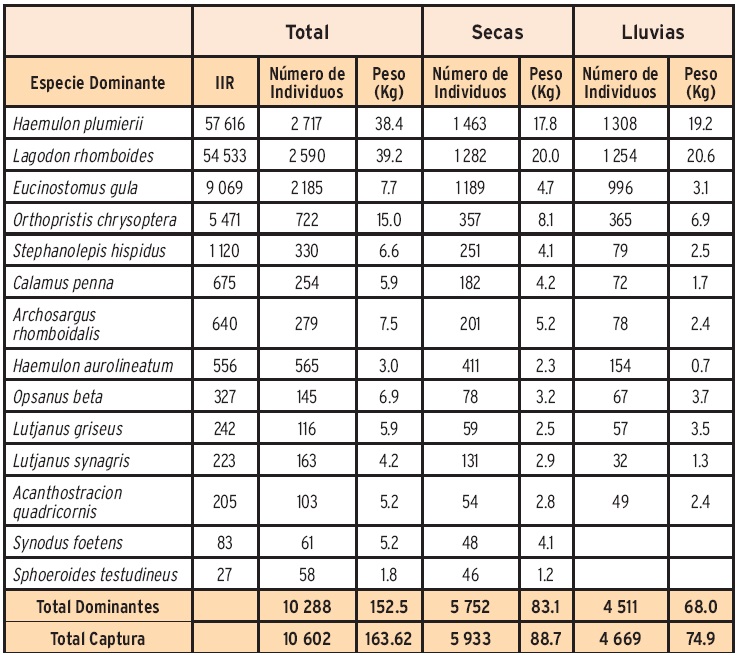
En conjunto, se identificaron 14 especies con dominio ecológico, las cuales representaron el 97.04 % en número de especímenes y el 93.32% en peso de la captura total (Tabla 4). La especie más importante por su abundancia y peso fue H plumierii, localmente conocido como Chac-chi (vocablo maya que significa boca roja). En la época de secas se capturó un total de 5 933 especímenes, con un peso conjunto de 88.7 kg, y se identificaron 14 especies dominantes, las cuales representaron el 96.9 % en número de especímenes y el 93.6 % en peso de la captura total. Las especies más importantes por su abundancia y peso fueron H. plumierii y L. rhomboides. En contraste, en la época de lluvias se capturaron 4 669 individuos con un peso total de 74.9 kg con 12 especies dominantes, las cuales representaron el 96.6% en número de individuos y el 90.7 % en peso de la captura total, manteniéndose la mayor abundancia proporcional de L rhomboides y H. plumierii.
En la Figura 4 se muestra el dendrograma con las asociaciones para el ciclo nictemeral (día y noche), así como la descripción de los códigos alfa asociados a sus nombres científicos, estableciéndose la presencia de dos grupos principales. Se identificaron 15 especies dominantes para horas de día con L. rhomboides como la especie más abundante, en tanto que para horas de noche se identificaron 13 especies dominantes, todas ellas presentes en el grupo de especies dominantes del día, destacando para horas de la noche los valores de abundancia de H plumierii.
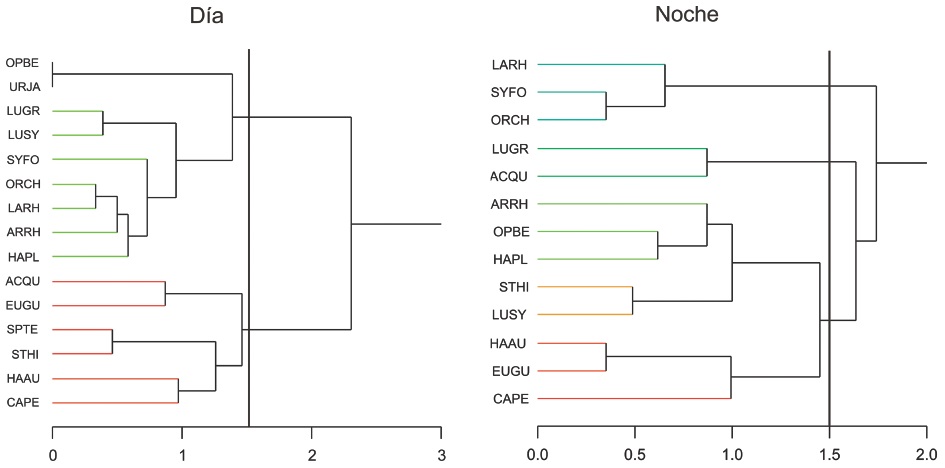
Figura 4. Dendrogramas del análisis de la abundancia de las especies dominantes en la Reserva de la Biósfera Los Petenes por ciclo nictemeral. Los nombres de las especies se representan mediante los códigos alfa descritos en la Tabla 3.
Figure 4. Dendrograms of abundance analysis of the dominant species in los Petenes Biosphere Reserve by nyctohemeral cycle. The specie's names are represented by alpha codes described in Table 3.
En la Figura 5 se muestran los dendrogramas que representan las asociaciones de las especies dominantes para cada época climática (secas y lluvias). En la época de secas se identificaron tres asociaciones principales con 14 especies dominantes. En tanto que en la época de lluvias se identifican solo dos grupos con 12 especies dominantes y todas ellas fueron también especies dominantes en la época de secas.
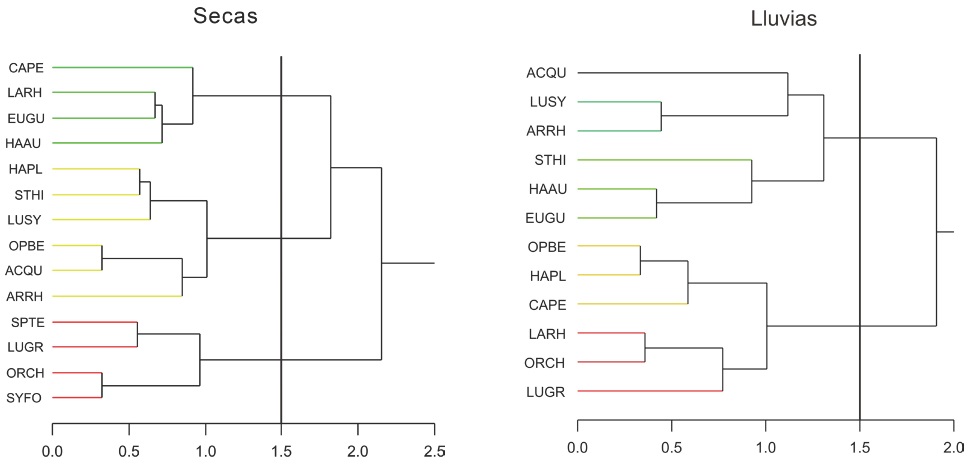
Figura 5. Dendrogramas del análisis de la abundancia de las especies dominantes en la Reserva de la Biósfera Los Petenes por épocas climáticas (Secas y Lluvias, respectivamente). Los nombres de las especies se representan mediante los códigos alfa descritos en la Tabla 3.
Figure 5. Dendrograms of abundance analysis of the dominant species in Los Petenes Biosphere Reserve by climatic periods (Dry and Rainy seasons, respectively). The specie's names are represented by alpha codes described in Table 3.
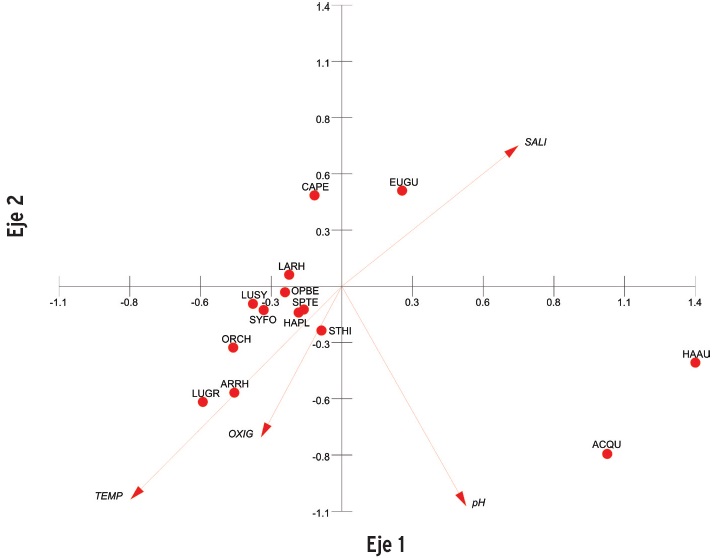
El ACC que relaciona la composición y abundancia de los peces dominantes con la variabilidad ambiental, se realizó con los datos agrupados por época climáticas, ya que no se encontró una diferencia de interés en la composición de las especies afectadas por el cielo nictemeral. En la época de secas, el análisis expresado en los dos ejes principales, alcanzó los eigenvalores de 0.199 y 0.094 para el eje uno y dos respectivamente, con una varianza de 26.037 y 12.271 respectivamente (Figura 6). La correlación entre la abundancia y la variable ambiental en el eje uno fue de 0.904, y para el eje dos de 0.856. Se destaca la relación existente entre salinidad y abundancia de E. gula en el cuadrante uno, entre el pH y A quadrieornis, y H. aurolincatum en cuadrante dos. El resto de las especies dominantes se vincularon con la temperatura y el oxígeno disuelto, aunque en una relación muy débil.
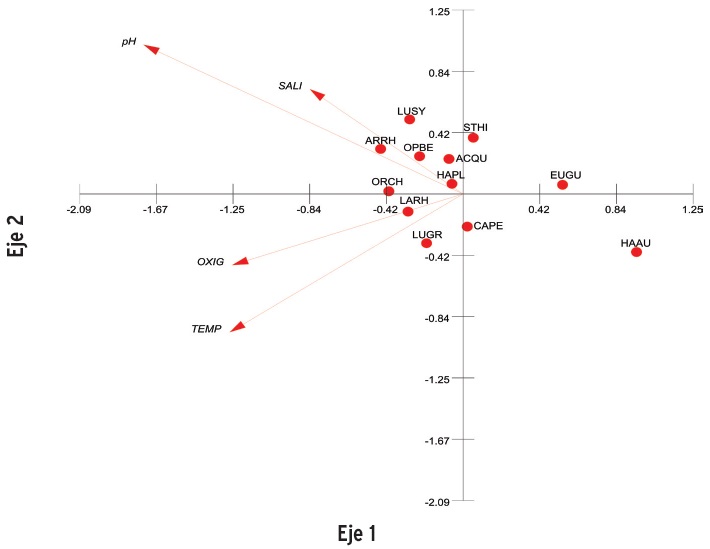
En el caso de la época de lluvias, el ACC presentó eigenvalores de 0.142 para el eje 1, y de 0.022 para el eje 2, con una varianza de 31.56 y 4.83 respectivamente (Figura 7). La correlación entre la abundancia y las variables ambientales en el eje uno fue de 0.806, y para el eje dos de 0.64. En los cuadrantes uno y dos solo se refleja la abundancia de S. hispidus, E gula, C. penna y H. aurolincatum, y los efectos de las variables ambientales se orientaron hacia los cuadrantes tres y cuatro, lo que se interpreta como una correlación no significativa con la abundancia. Finalmente, se destaca la asociación encontrada entre L. rhomboidalis y el oxígeno disuelto en el cuadrante tres.
DISCUSIÓN
Las variables ambientales de la porción litoral de la RBLP están influenciadas principalmente por la temporalidad climática (época de lluvias y de secas), el efecto de las mareas, las condiciones de baja profundidad y alta transparencia, la amplia cobertura de pastos marinos y los afloramientos de ríos subterráneos que transportan agua continental.
En el sitio de muestreo, la profundidad osciló entre 2 m y 2.5 m, a pesar de ubicarse a una distancia de cerca de 2 km de la línea de costa. El cambio en la profundidad por efecto de la marea fue mínimo durante el ciclo nictemeral y ocasionalmente el viento provocó mayor movimiento de agua que se registró en la medición de profundidad. Las características de gran amplitud (cerca de 200km) y muy baja inclinación de la plataforma continental en esta región, permite el establecimiento de abundantes parches de vegetación sumergida que contribuyen a incrementar los niveles de transparencia del agua. En este sentido, las diferencias entre las variables ambientales de día y noche no fueron significativas (P-≤ = 0.05). De acuerdo con Castillo-Rivera y col. (2005), las mareas constituyen una de las fuentes de energía principales en sistemas acuáticos costeros condicionando la variabilidad ambiental nictemeral; sin embargo, esto no fue observado para la asociación de peces dominantes.
La temperatura del agua mostró un intervalo de variación cercano a los 10 °C en un ciclo anual, lo cual es un reflejo de la condición tropical de la zona de estudio. En este sentido, Ayala-Pérez y col. (2012), describen la variación de la temperatura ambiente de tres estaciones meteorológicas de Campeche, aledañas a la RBLP, las cuales son similares a lo que se reporta en el presente estudio.
La salinidad del mar en la RBLP varió por efecto del aporte de agua continental, tendiendo a disminuir durante la época de lluvias y a aumentar al final de la época de secas por el incremento en la evaporación asociada a los altos valores de temperatura. Este comportamiento ha sido reportado previamente por Muñoz-Rojas y col. (2013), para la misma región.
Los valores de pH registrados en este estudio son comparables con los reportados por Vega-Cendejas (2004), para la Reserva de la Biósfera Celestún, que es un sistema colindante y en ambos estudios se coincide en señalar, que la materia orgánica aportada desde el continente, así como los niveles de productividad primaria, se suman a las condiciones de temperatura y salinidad para definir los valores de pH. La oxidación de la materia orgánica y el consumo de bióxido de carbono por la fotosíntesis son procesos que deben ser considerados para explicar los valores de pH, tal como lo menciona Hernández-Ayón y col. (2003).
Finalmente, el oxígeno disuelto es una variable que cambia rápidamente dependiendo de las condiciones de temperatura y salinidad, pero también por el desarrollo de procesos biológicos como la oxidación de la materia orgánica y producción fotosintética, sin dejar de considerar la presencia e intensidad del viento. Durante la época de lluvias, los valores de oxígeno disuelto son más bajos dado el incremento en la temperatura del agua, pero además, por la aportación de materia orgánica alóctona, transportada por las descargas de agua continental. Este comportamiento también ha sido discutido por Ayala-Pérez y col. (2012), para la región sur de la costa de Campeche, donde las aportaciones alóctonas son de mayor magnitud por la descarga de toda la red hidrológica de la cuenca del Grijalva-Usumacinta.
Estadísticamente existen diferencias significativas entre las épocas de secas y lluvias, considerando las variables de temperatura, salinidad y pH, sin embargo, para el caso del oxígeno disuelto, la diferencia no fue significativa (P = 0.37), lo que permite argumentar la continua e intensa actividad biológica tanto de procesos fotosintéticos fitoplanctónicos, como de los pastos marinos, así como las interacciones entre funciones de fuerza, como la temperatura, el viento y la aportación de agua continental.
Los sistemas costeros tropicales se caracterizan por una compleja estructura ecológica y una amplia variedad de hábitats con elevada productividad primaria, lo cual es aprovechado para el desarrollo de todo o parte de los ciclos de vida de los peces (Ayala-Pérez y Martínez-Romero, 2011). El caso de la RBLP no es la excepción, ya que de las 44 especies de peces encontradas, se identificaron componentes de origen marino, principalmente especies estuarinas e incluso especies típicamente dulceacuícolas, como es el caso de Ciehlasoma urophthalmus, la cual también fue reportada por Torres-Castro y col. (2008), en una comunidad integrada por 44 especies de peces que habitan en ojos de agua permanentes y temporales de la porción continental de la RBLP. Su presencia en la porción marina permite argumentar el vínculo ecológico con ecosistemas dulceacuícolas. Es preciso también mencionar que Caso-Chávez y col. (1986), describieron una población de Curophthalmus, asociada a los pastos marinos del litoral interno de la Laguna de Términos, en condiciones de alta salinidad, lo cual constituye un indicador de la gran capacidad adaptativa de la especie, tal y como lo describe Sehmitter-Soto (2007).
Las familias Haemulidae, Sparidae y Lutjanidae se distinguieron por su diversidad y abundancia en la RBLP; en tanto que en la Reserva de la biósfera Ría Celestún, un ecosistema cercano, a menos de 250 km por mar, se destaca la presencia de las familias Sciaenidae, Sparidae y Carangidae (Vega-Cendejas, 2004), lo cual permite visualizar a grandes rasgos las asociaciones y el nivel de especialidad que el hábitat les permite a las especies. En este sentido, comparando la estructura de la comunidad de peces de la RBLP, con lo reportado para ecosistemas vecinos, se encontró que Ayala-Pérez y col. (2003), reportaron 107 especies de peces para la Laguna de Términos, de las cuales 30 (68 %), coinciden con los resultados de este estudio. Asimismo, Ayala-Pérez y col. (2012), reportaron 93 especies en la porción occidental de la costa de Campeche y la Laguna de Términos, de las cuales 23 (52 %) son coincidentes. Finalmente, Muñoz-Rojas y col. (2013), integraron un listado de 46 especies para toda la porción costero-litoral de la RBLP, de la cual el 72 % coincidieron con los resultados aquí reportados.
Las especies dominantes H. plumierii (Haemulidae) y L. rhomboides (Sparidae), se distinguieron por su abundancia relativa (50% del total), y contribución en peso (47%). Muñoz-Rojas y col. (2013) reportaron también a estas dos especies como dominantes para todo el litoral de la RBLP, aportando el 59 % en peso y 67 % en número de individuos del total de la captura. De igual manera Sosa-López y col. (2010), reportaron a estas dos especies como dominantes en el litoral Campechano, representando juntas, el 25 % de la captura total.
El resto de las especies dominantes encontradas a lo largo del estudio fueron: Eucinostomus gula, Orthopristis ehrysoptera, Calamus penna, Lutjanus griseus, Opsanus beta y Stephanolepis hispidus. Estas especies son principalmente marinas y desarrollan parte de su ciclo de vida en aguas estuarinas, y en etapas juveniles utilizan las zonas de pastos sumergidos para alimentarse, crecer y refugiarse (Castro-Aguirre y col., 1999).
Las asociaciones de especies tanto en la escala temporal como nictemeral son producto de su abundancia y frecuencia de aparición, sin embargo, permiten discutir la función ecológica que cada asociación tendría en el sistema. Durante la época de secas, 14 especies dominantes conformaron tres asociaciones. El primer grupo estuvo integrado por especies generalistas que se alimentan de pequeños peces, crustáceos, poliquetos, nematodos, plancton y algas que habitan sobre las praderas de pastos marinos (Morales-López y col., 2007). El segundo grupo lo conformaron seis especies, entre las cuales destaca Hplumierii, por ser la más abundante y estar estrechamente vinculada con los pastos marinos. L. synagrises una especie demersal, carnívora generalista y oportunista, que prefiere hábitats de arrecifes de coral y zonas de arena con vegetación, en aguas transparentes, pero que se asume aprovecha el hábitat para actividades de alimentación (Landínez-Gareía y col., 2009). S. hispidus y A. quadrieornis son especies asociadas a fondos rocosos y arenosos, con preferencias por vegetación sumergida. Los individuos capturados corresponden a juveniles, dadas sus tallas (Mexicano-Cíntora y col., 2007). Muñoz-Rojas y col. (2013), mencionan que A. rhomboidal y O. beta, son especies que se encuentran frecuentemente en fondos lodosos, entre raíces de manglar y sobre fondos arenosos cubiertos por vegetación sumergida. El tercer grupo se integra por cuatro especies, de las cuales: S. testudineus comúnmente se encuentra en bahías y aguas costeras protegidas, especialmente en lechos de algas marinas y aguas salobres, y tiene preferencia alimentaria por bivalvos, gasterópodos y macrófitas (Chi-Espínola y Vega-Cendejas, 2013). L. griseus, es una especie abundante en sistemas adyacentes como El Cayo, en Laguna de Términos, donde ha sido descrita como dominante, con interacción con fanerógamas marinas y manglar (Guevara y col., 2007); O. chrysoptera es una especie generalista, que habita sobre praderas de pastos marinos (Morales-López y col., 2007); mientras que S. foetens, es una especie vinculada con arrecifes y se le asocia constantemente con praderas de pastos marinos y fondos relativamente someros (Bedia y Franeo, 2008).
Para la época de lluvias, se identificaron 12 especies dominantes, que son exactamente las mismas encontradas para la época de secas, a excepción de S. testudineus y S foetens. Con el mismo nivel de similitud, en la época de lluvias solo se distinguen dos asociaciones que no tienen diferencia estadística con las asociaciones de la época de secas. Las especies dominantes mantienen el control de la exergía en el sistema durante todo el ciclo anual.
Para la escala nictemeral, durante las horas de día se identificaron 15 especies dominantes asociadas en dos grupos, mientras que en las horas de noche, 13jespecies fueron dominantes, las cuales formaron tres grupos al mismo nivel de similitud. Igual que en la escala temporal, no existen diferencias estadísticas entre las asociaciones de día y de noche. Las especies dominantes que son diferentes entre las agrupaciones de día y noche son S testudineus y U. jamaicensis.
La correspondencia canónica entre la abundancia de las especies dominantes y las variables ambientales durante la época de secas, destaca a la salinidad como la variable con mayor varianza, particularmente durante junio. Este comportamiento se asocia con la abundancia de E. gula, que es una especie marina con preferencia estuarina y por tanto, tiene gran capacidad de tolerancia a los cambios en la salinidad (Mexicano-Cíntora, 1999). En menor grado, se aprecia la relación entre el pH y H. aurolincatum y A. quadrieornis. La mayor agrupación de abundancia de las especies dominantes se presenta en el cuadrante tres donde se asocian con las variables de temperatura y oxígeno disuelto en forma negativa.
Para el caso de la época de lluvias, las correlaciones canónicas solo se representan en los cuadrantes tres y cuatro, donde también se concentran las especies dominantes. En los cuadrantes uno y dos se mantiene la presencia de Egula y Haurolincatum respectivamente. Diversos estudios han señalado la influencia de las variables ambientales sobre la abundancia y composición de las asociaciones de peces (Kupschus y Tremain, 2001; Torres-Castro y col., 2008; Petry y col., 2003), sin embargo, en la RBLP, la variabilidad ambiental, al menos para la porción costero-litoral, tiene características de alta homogeneidad debido a la amplia cobertura de vegetación sumergida. De acuerdo con Allen y col. (2006), las áreas con grandes praderas de pastos marinos son un buen criadero, centro de alimentación y protección, y dan estabilidad ambiental.
CONCLUSIONES
La RBLP es un hábitat crítico para muchas especies de peces que desarrollan actividades de alimentación, protección, crianza y refugio temporal con características particulares de abundancia, diversidad y distribución en espacio y tiempo. El análisis de la variabilidad ambiental entre épocas climáticas permitió encontrar diferencias significativas entre secas y lluvias, considerando la temperatura, salinidad y pH. Las diferencias entre los ciclos día y noche no fueron significativas dadas las condiciones de profundidad, del gradiente de inclinación de la plataforma continental y a la amplia cobertura de vegetación sumergida, los cuales en conjunto mitigan el efecto de la marea, que se observa como una de las más importantes funciones de fuerza del sistema. Las comunidades de peces en la Reserva de la biósfera Los Petenes aproveehan en espacio y tiempo, las condiciones de hábitat para desarrollar todo o parte de su ciclo de vida; su estructura refleja la interacción con ecosistemas vecinos, con representantes propios de arrecifes, como la familia Lujtanidae y de agua dulce como la familia Cichlidae. Los pastos marinos, además de constituir un área de refugio, ofrecen abundante alimento, pero además propician un ambiente homogéneo en cuanto a la variabilidad ambiental, de tal forma que las relaciones inter e intraespecíficas de las especies dominantes no cambian significativamente en escalas temporal y nictemeral. Estos resultados sugieren una eficiente utilización de la oferta energética que conceden las praderas de T. testudinum, por parte de la ietiofauna asociada.

