











 nueva página del texto (beta)
nueva página del texto (beta) Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
Permalink
Hechtioideae Givnish currently contain three genera, based on morphological, molecular, and biogeographical evidence (Ramírez-Morillo et al. 2018a, b), all previously included in the genus Hechtia Klotzsch (Givnish et al. 2007). The first one, Mesoamerantha I. Ramírez & K. Romero (Ramírez-Morillo et al. 2018b), with three species confined to the geographical area south of the Motagua River in Guatemala to the northern portion of Nicaragua (Romero-Soler et al. 2022a), in the drier regions north of the Lago Cocibolca, with a disjunct population of M. guatemalensis (Mez) I. Ramírez & K. Romero in Belize (Holst et al. 2017). The second genus is Bakerantha L.B. Sm., with five species endemics to the central eastern region of Mexico (Romero-Soler et al. 2022b), with plants that vegetatively resemble those of the distantly related species of Pitcairnia L’Hér. by their long, hanging leaves, especially on species such as B. lundelliorum (L.B. Sm.) I. Ramírez & K. Romero and B. tillandsioides (André) L.B. Sm. The third genus is Hechtia, groups several different lineages (see Ramírez-Morillo et al. 2018b), that includes from cespitose to rhizomatous rosettes, with central to lateral inflorescences, variable inflorescence branching pattern, floral color, fruit texture, and seed (Ramírez-Morillo et al. 2018b; Espejo-Serna et al. 2020). Hechtia is the genus with the widest geographical distribution and larger number of species (with more than 92 species for Hechtioideae sensuGouda & Butcher 2022; our estimate is 100 taxa for the subfamily, five for Bakerantha, three for Mesoamerantha (Ramírez-Morillo et al. 2018b) and 92, including the species here described, for the genus Hechtia), spreading from the south of the USA throughout Mexico to at least Honduras, within an area called Megamexico III by Rzedowski (1991), the distributional range for Hechtioideae.
The relationships within Hechtia are not clear yet but some lineages show high support (Ramírez-Morillo et al. 2018a, b, Rivera-Martínez et al. 2022). One of them is the Hechtia glomerata complex (Clade D in Ramírez-Morillo et al. 2018a, b), basically characterized by the presence of lateral, glomerulate, floccose infructescences, bearing fragrant flowers with a superior or almost superior ovary and mostly white petals. A previous phylogenetic hypothesis (Ramírez-Morillo et al. 2018a) indicates that all these characters have evolved independently more than once in the family but are not linked; we call it the “Hechtia glomerata syndrome”. Despite the monophyletic nature of the Hechtia glomerata complex, all species with lateral inflorescence challenge taxonomists because the wide variation in the size of the plants and inflorescences (Ramírez-Morillo et al. 2018a, Espejo-Serna et al. 2020), mostly on features related with the age of the polycarpic rosette, i.e., an individual rosette blooms many times, sometimes more than once in one year, the larger and older the rosette gets, the larger and more branched the inflorescence/infructescence become. These age-related changes are linked to the poor understanding of species limits in this lineage, with other species remaining undescribed, or few proposed as described twice (sensuBurt-Utley et al. 2011), when treated H. mexicana L.B. Sm. as a synonym of H. glomerata Zucc., and H. elliptica L.B. Sm. and H. zacatecae L.B. Sm. as synonyms of H. texensis L.B. Sm. Although field work allows the observation and understanding of population variation, cultivation of plants is an excellent choice when long-term field observations cannot be conducted. Daily observations allow detailed recording of changes on vegetative and floral characters, usually overseen in the field and definitively impossible to see on herbarium material.
About 15 years ago, Dr. Adolfo Espejo-Serna showed us a picture of a new species also from the Mexican State of San Luis Potosí (López-Ferrari et al. 3059, CICY, SLPM, UAMIZ); in 2018, CTHL visited the botanical garden El Izotal from the Universidad Autónoma de San Luis Potosí and saw the same species in bloom, mentioning the most remarkable feature of the species: lateral inflorescences and pink petals in female and male flowers. In 2020 IMRM visited the same botanical garden and photographed a staminate individual in bloom. In 2021, Ing. Francisco Sánchez Barra, who is in charge of the garden, managed to get pistillate plants in bloom. Based on these material and herbarium specimens reviewed in several herbaria, the new species is described and illustrated herein.
Materials and methods
This work is based on the study of specimens deposited in the following herbaria: CICY, ENCB, HGOM, IEB, MEXU, SLPM, TEX-LL, and UAMIZ (acronyms according to Thiers 2022). Also, live specimens were studied and photographed from cultivated plants at the Botanical Garden El Izotal from the Universidad Autónoma de San Luis Potosí. Micromorphological characters of pollen and seed were obtained from herbarium and fresh material and analyzed using images captured using a scanning electron microscope (SEM) at Centro de Investigación Científica de Yucatán, A.C. A geographic distribution map was created using QGIS 3.12 (qgis.org). In some cases, geographic coordinates were estimated from label data on herbarium specimens using Google Earth Pro v. 7.3.2.5776. These estimates of coordinates are provided within brackets in the specimen citations. For morphological terminology we used Smith & Downs (1974) and Hornung-Leoni & Gaviria (1999), for growth patterns Ramírez-Morillo et al. (2014). Limits of the biogeographical provinces were defined following Morrone et al. (2017). An assessment of the conservation status of the species was carried out using the IUCN methodology (www.iucnredlist.org). We relied on the set of B criteria, which allows an evaluation based on data on the distribution of the species, obtained from herbarium specimens. The subcriteria B1 (extent of occurrence, EOO) and B2 (area of occupancy, AOO), were calculated through the GeoCAT software (Bachman et al. 2011). These assessments were further documented with our own field experience and published data.
Results
Hechtia sanchezii I. Ramírez & C.T. Hornung, sp. nov. (Figures 1-3, 5).
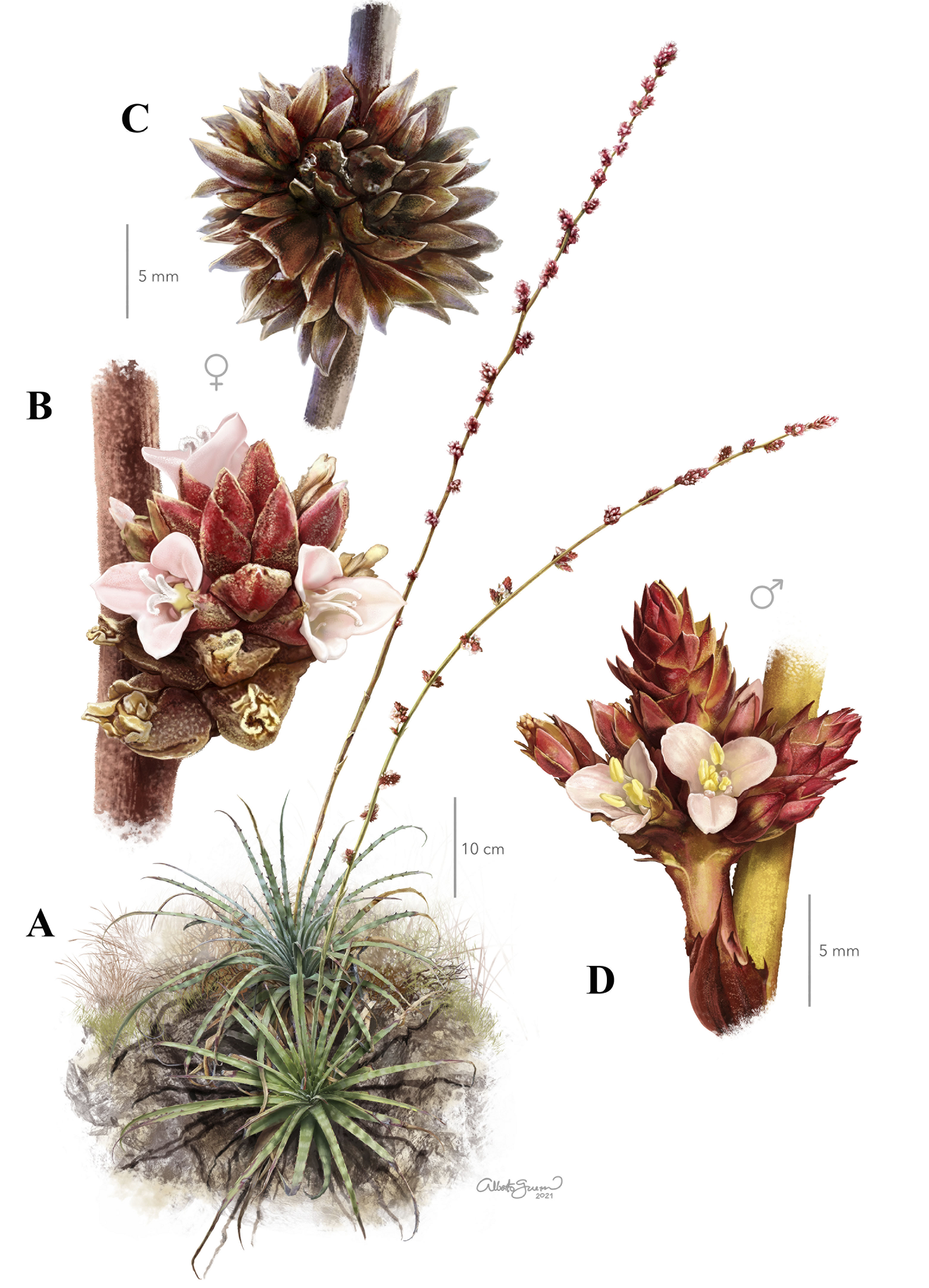
Figure 1 Hechtia sanchezii. A) Plants in bloom, showing the pistillate (right) and staminate (left) inflorescences, B) female flowers, C) open fruits, D) male flowers. Illustrated by Alberto Guerra based on photographs by (A) Ivón M. Ramírez-Morillo, (B) Francisco Sánchez Barra, (C) Claudia T. Hornung-Leoni, (D) Germán Carnevali.
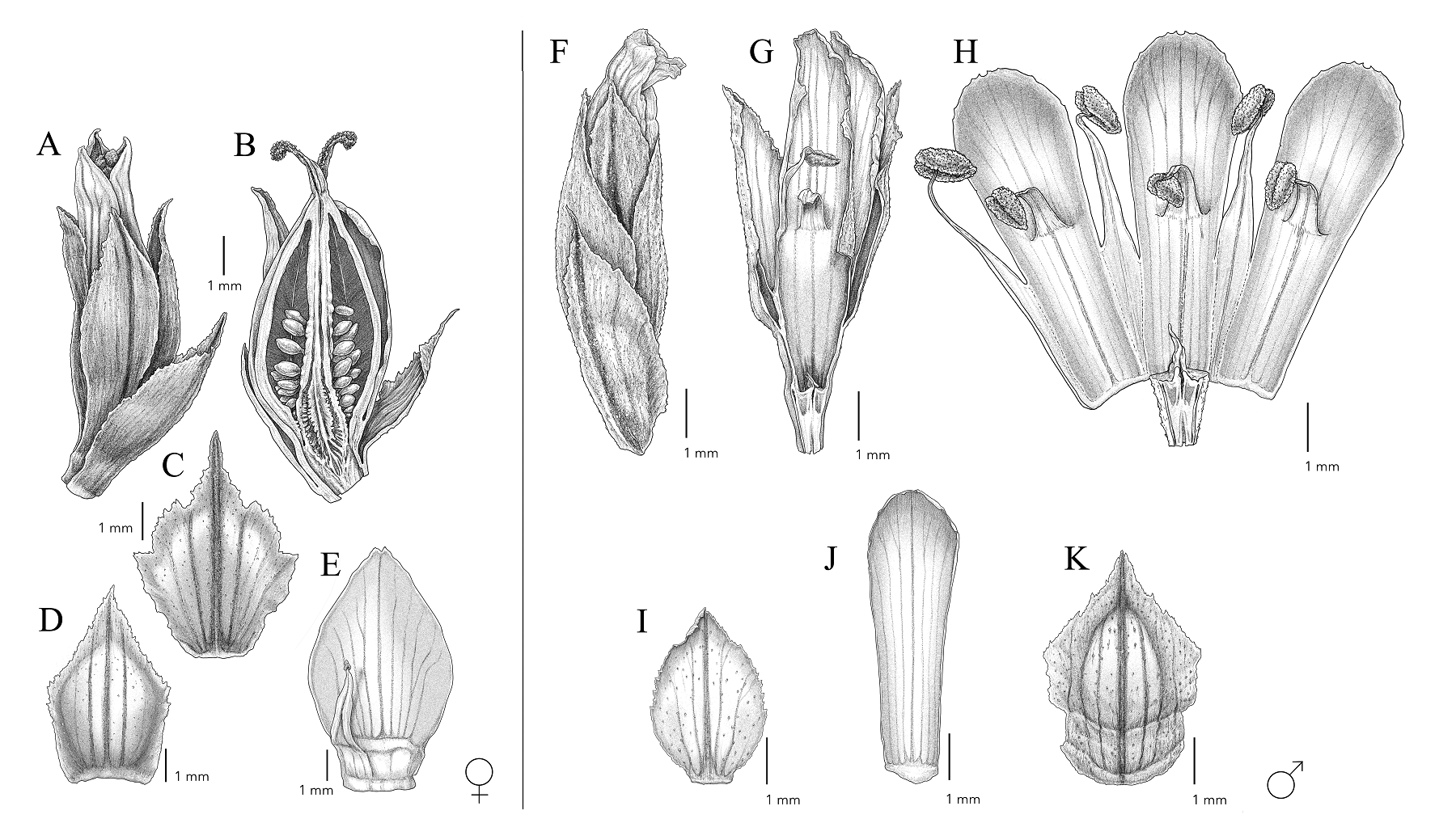
Figure 2 Hechtia sanchezii (A-E) Female flower, A) flower closed, B) longitudinal section showing ovary and ovules, C) floral bract, D) sepal, E) petal and staminode. (F-K) Male flower, F) flower closed, G) longitudinal section of the flower, H) details of stamens, I) sepal, J) petal, K) floral bract. Illustrated by Alberto Guerra based on (A-E) F. Sánchez 2079 (SLPM), (F-K) I. Ramírez et al. 2441a (CICY).

Figure 3 A) Type specimen of Hechtia sanchezii (S. Zamudio 13454 ♂, isotype CICY), B) details of the rosette, C) lateral inflorescence development in H. sanchezii, D) details of foliar indumenta (adaxial, upper image, abaxial surface, bottom image of a young leaf, E) male branches, F) pollen, G) seed. Photographs by Ivón Ramírez-Morillo (A-E), Lilia Can (F-G).
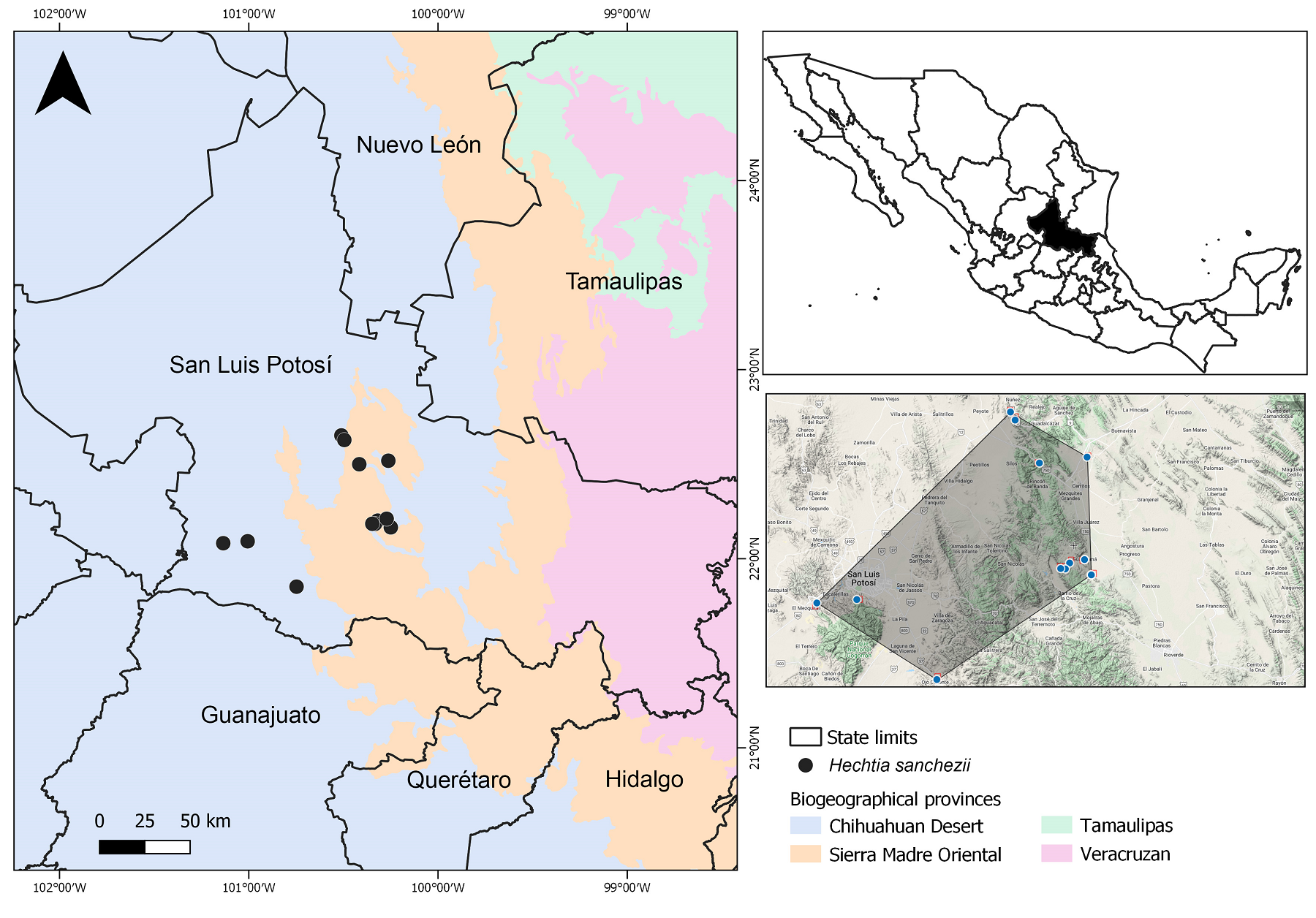
Figure 4 Distribution of Hechtia sanchezii based on specimens collected in central San Luis Potosí, the polygon generated (GeoCAT) with their information.

Figure 5 Comparison of most relevant morphological features between Hechtia glomerata (A, C, E, G) and Hechtia sanchezii (B, D, F, H). A, B) rosettes, note foliar surface; C, D) male inflorescences, note the shape of the branches; E, F) pistillate flowers, note floral colors; G, H) staminate flowers, note indument and floral colors. Photographs by (A, C, D) Ivón Ramírez-Morillo, (B, E, G) Claudia T. Hornung-Leoni, (F) Francisco Sánchez Barra, (H) Germán Carnevali.
Type. Mexico. San Luis Potosí, municipio Villa Juárez, +/- 2 km al SW de Buenavista, 22° 12’ 07’’ N; 100° 19’ 09’’ W, 1,281 m, 5 julio 2006, S. Zamudio 13454 ♂ (Holotype: IEB!; isotypes: CICY!, UAMIZ!).
Diagnosis. Hechtia sanchezii differs from H. glomerata in having red to ferruginous indument on the bracts and sepals (vs. white), floral bracts and sepals red (vs. greenish or brownish), pink (vs. white) petals, scentless (vs. fragrant) flowers, stamens with filaments of two lengths, three epipetalous, with the short filaments attached at mid length of the petal, and three alternating with petals with filaments basally connate to the base of them (vs. two filaments of different lengths but attached to each petal).
Description. Plants lithophytic, rosettes cespitose, globose in general shape, 25-30 cm tall, 35-40 cm diameter, generally forming dense, large colonies of 20-30 rosettes, the new ones are produced in the periphery of the clump where they become asymmetrical by the clumping of the rosettes. Rhizomes short to inconspicuous. Leaves 30-40 in number, rigid, reflexed; sheaths broadly ovate, 4-6 × 4.5-6.5 cm, light brown, a round-shaped, dark brown spot in the middle of it, margins erose, thin and soft when dry, densely white lepidote abaxially, lustrous and glabrous adaxially; blades narrowly triangular, attenuate, 25-30 ×1.8(-4) cm, succulent, channeled in cross section then margins erect when live, light green, densely white lepidote when young and becoming glabrous with age, with small trichome spots, minutely verrucose on both surfaces when dry, densely white lepidote below, margin armed; spines antrorse and erect due to the transversely channeled foliar blade, triangular, 3-4 mm long, 1.5-2 cm apart, opposite to alternate, reddish, pungent, with a tuff of white trichomes at the axile of the basal ones, shorter than the spine. Inflorescence lateral, erect to arcuate, emerging from a mature rosette (type PMP growth pattern). Staminate inflorescences a 2-divided panicle, narrow cylindrical in shape, erect to arcuate, 40-100 cm long; peduncle terete, (15-)30-40 cm long, (3-)4-5 mm diameter at the base, surpassing the rosette, internodes (2-)4.5-5 cm long, purple when young becoming brown as flowering progresses, glabrous; peduncle bracts surpassing (basal ones) or not (apical ones) internodes, with triangular sheaths, almost enclosing the peduncle, 2.5-3 × 0.4-0.5 cm, light green, drying light brown, margins entire to erose, thin, the blades foliaceous, triangular, long attenuate, multinerved, 3-4 × 0.2-0.4 cm, basally spinose, densely white lepidote abaxially, glabrous adaxially; main axis 20-50 cm long, 4-5 mm diameter, terete, reddish, drying brown, internodes 2.5-5 cm long; primary bracts triangular, attenuate, 1.5-2 × 0.4-0.5 cm, shorter than branches except those at the base where they are as long as the branch (usually one branch), brown when live and dry, sparsely white-lepidote on both surfaces, margin spinose-dentate; primary branches 7-9 in number, forming an angle of 45° or less with the main axis, secondary branches 1-3 in number, 1.5-3 cm long, less than 1 cm diameter, each with 40-70 flowers; stipe ca. 0.5 mm long, 2-3 mm diameter, flattened, drying brown, glabrous; floral bracts wide-triangular to oblong, acute, 5.5-6 × 3-5 mm, membranous, white basally, then red, and apically brown, margin erose, punctuated adaxially, 5-nerved, erect and adnate to the sepals. Flowers sessile, polystichous, actinomorphic; sepals wide-elliptical, 3.5-4 × 2 mm, red, minutely serrulate, scarcely punctulate, 5-nerved, shorter than petals, erect; petals oblong to obovate, rounded, 5-6(-7.5) × 1.5-2.5 mm, pink, 7-nerved; filaments in two lengths, the shortest one attached to the petal mid length, ca. 2-3 mm long, white, the longest between the margins of petals, 4-4.2 mm long, with triangular filaments, flattened, basally connate; anthers oblong, ca. 1.5 × 0.6 mm long, dorsifixed, pale yellow, pollen yellow, ellipsoid, monosulcate, 37-40 µm polar, 17 µm equatorial, reticulate; pistillode reduced, ca. 2 mm long, greenish-yellowish, stigmatic lobes reduced, erect, ca. 0.5 mm long. Pistillate inflorescences a 1-divided panicle to rarely 2-divided panicle, cylindrical in shape, erect, 30-40(-100) cm long; peduncle terete, 20-30 cm long, 5-6 mm diameter at the base, reddish, internodes 2.4-5.6 cm long, glabrous; peduncle bracts much shorter (1/2 to 1/3 of the length) than internodes, with oblong sheaths, 1-8 × 5-6 mm, green, drying light brown, a central spot dark brown, margins entire to erose, thin, the blades foliaceous, narrowly triangular, long attenuate, slightly pungent, multinerved, 1-2 × 0.5-0.6 cm, sparsely spinose, green with purple spots or reddish, drying light brown, densely white lepidote abaxially, glabrous adaxially; main axis 25-75 cm long, 2-3 mm diameter, terete, green to red, drying dark brown, internodes (1-)3-4.5 cm long; primary bracts with no distinction between sheath and blade, mostly present on the most basal branches, wide triangular, long and abruptly attenuate, 0.9-1.7 × 0.6-0.8 cm, longer than branches (the most basal ones), to shorter than branches to absent on apical branches, red becoming brown, sparsely white-lepidote, margin sparsely spinose to serrulate, thin, hyaline; branches 7-20 in number; secondary branches sessile, 1-4 in number, fasciculate, ellipsoid to wide-oblong in shape, (1-)2-3 cm long, 1-1.5 cm diameter, with 10-40 flowers each; floral bracts elliptical to ovate-elliptical, abruptly acute, 5-7 × 4-5 mm, red to purple, sparsely punctulate, margin sparsely serrulate, irregular, 5-nerved, erect. Flowers almost sessile, polystichous, actinomorphic; sepals ovate to elliptical, acute-attenuate, (4-)6-7 × (3-)4-6 mm, red, minutely serruate, pubescent at margin, 5-nerved; petals ovate to elliptical, 5-7 × 3-4 mm, pink, entire, 9-nerved; staminodes six, hyaline, triangular, laminar, ca. 4 mm long, white; ovary superior, ellipsoid, 5-7 mm long, 2-3 mm diameter, white, slightly lepidote, stigmatic lobes recurved, 2-3 mm long, white. Fruits ellipsoid, sessile, 9-12 mm long, 6-9 mm diameter, glabrous, erect, brown when mature, septicidal and loculicidal dehiscence; seeds fusiform, brown to reddish, 3-5.6 mm long, 1-1.4 mm diameter, with two apical wings, these hyalines.
Distribution and ecology. Plants of Hechtia sanchezii have been found in submontane shrublands, with species such as Gochnatia hypoleuca (DC.) A. Gray (Asteraceae), Dasylirion longissimum Lem. (Asparagaceae), on gypsum soils, along with species of Selaginella P. Beauv. (Selaginellaceae) among other species. This new species is presently reported from San Luis Potosí, but it may be found in neighboring states due to similarities in the associated vegetation types. The actual distribution of Hechtia sanchezii occurs within the biogeographical areas of the Chihuahuan Desert and Sierra Madre Oriental according to Morrone et al. (2017), specifically in the region of the Zacatecano-Potosino Plateau (sensu CONABIO www.gob.mx/conabio), characterized by arid to semiarid climates, on elevations from 1,274 to 2,280 m, inhabiting xerophytic shrubland on gypsum substrate, as several other species in the genus.
Preliminary conservation status. The EOO is 4,438 km2, within the threshold for “Endangered” status under subcriterion B1, while the AOO is estimated as 48 km2, which is below the threshold for “Endangered” status under subcriterion B2. Hechtia sanchezii is only known from 20 collections, which represents 12 occurrences, and about 10 subpopulations, collected between 1955 to 2021 from San Luis Potosí. Most of these collections have been made along roads or near urban areas, which may have a negative impact on the habitat of this species, causing its fragmentation and degradation in the short to medium term. Therefore, these 12 occurrences represent nine locations (www.iucnredlist.org) with regard to the most serious plausible threats (road impacts and urbanization). These threats predict a present and future decline of the quality of its habitat. Hechtia sanchezii is therefore assigned a preliminary status of “Vulnerable” [VU B1ab(iii)+2ab(iii)].
Phenology. Specimens have been collected with flowers during June, July, and September, with fruits in September.
Etymology . The specific Latin epithet honors Francisco Sánchez Barra, the garden manager of Botanical Garden El Izotal, located in Tangamanga Park, owned and managed by Universidad Autónoma de San Luis Potosí, in Mexico. Francisco Sánchez Barra has collected and cultivated this species and many more from San Luis Potosí, he provided photographs and material that allow the description of the new species.
Additional specimens examined (paratypes). Mexico. San Luis Potosí, Mun. De Cerritos, cauce del arroyo El Sauz, ca. 7 km después de Pozo de Acuña, rumbo a Cerritos, 22° 31’ 07’’ N, 100° 15’ 47’’ W, 1,225 m, 4 September 2003, A.R. López-Ferrari, A. Espejo & J. Ceja 3047 fruits (UAMIZ); 1 km south of Guadalcázar, path to base of hills of town, 22° 30’ N; 100° 25’ W, 9 June 1989, D. & J. Bogler 621 ♂ (TEX-LL). Mun. De Ciudad Fernández, 3 km después de Puerto de Santa Gertrudis, ca. 12 km después de El Paraíso, en brecha a Saucillo a Puerta del Río, 22° 11’ 07’’ N, 100° 20’ 52’’ W, 1,302 m, 13 September 2015, I. Ramírez, A. Espejo, A. López-Ferrari, J. Ceja & A. Mendoza 1370 fruits (CICY). Mun. De Guadalcázar, ca. 3 km al noreste de Charco Blanco, 22° 39’ 14” N; 100° 30’ 37” W, 5 June 1964, F. Medellín Leal 1556 ♀ (SLPM), 1555 ♂ (SLPM); Ca. 10 km al W de Guadalcázar, 22° 37’ 42’’ N, 100° 29’ 44’’ W, 1,590 m, 4 September 2003, A.R. López-Ferrari, A. Espejo & J. Ceja 3042 fruits (UAMIZ). Mun. de San Luis Potosí, 3 km al W de Pozuelos, ca. 17.5 km al SW de San Luis Potosí, carretera San Luis Potosí-Aguascalientes, 22° 04’ 57’’ N; 101° 08’ 7’’ W, 2,280 m, 16 July, 1994, J. Ceja, A. Espejo & A.R. López-Ferrari 13 fruits (CICY, UAMIZ). Mun. Santa María del Río, Ojo Caliente, [21° 51 ‘10” N; 100° 44’ 53” W], 1,800 m, 5 June 1959, F. Takaki 133 ♂ (SLPM). Mun. De Villa de Arriaga, Sierra de San Miguelito, cañada de San Antonio, [22° 05’ 34” N; 101°00’ 23” W], 2,250 m, 5 enero 1955, J. Rzedowski 5679 fruits (SLPM). Mun. de Villa Juárez, ca. 1 km al oeste de Agua del Medio, creciendo sobre bancos de yeso, 22° 09’ 59.2’’ N, 100° 15’ 0.34’’ W, matorral xerófilo, 1,274 m, 28 October 2021, F. Sánchez 2079 ♀ (SLPM); same previous locality; 28 October 2019, I. Ramírez & N. Raigoza 2441a ♂ (CICY), 2441 fruits (CICY, SLPM); Cerro de Guascamá [Guaxcamá], [22° 12’ 42” N; 100° 16’ 19” W], 1,250 m, 26 March 1962, F. Takaki 1250 ♀ (ENCB); 8.5 km después de Guaxcamá, rumbo al Puerto de Santa Gertrudis, 22° 11’ 05’’ N, 100° 20’ 00’’ W, 1,418 m, 4 September 2003, A.R. López Ferrari, A. Espejo & J. Ceja 3059 ♂ (2xCICY, UAMIZ); Cerro de Guascamá [Guaxcamá], [22° 12’ 42” N; 100° 16’ 19” W], 26 March 1962, F. Takaki 1715 ♀ (MEXU, SLPM).
Discussion
Hechtia sanchezii is an interesting species due to the presence of a combination of morphological characters previously unknown in the genus. It is morphologically similar to members of the Hechtia glomerata complex by its cespitose growth, rosette with narrow triangular leaves, serrate margins, and lateral female inflorescences with glomerule-like branches. Nevertheless, staminate inflorescences of the new species have oblongoid branches (vs. pyramidal) and longer in H. glomerata, with floral bracts membranaceous (vs. stramineous). Live plants of the new species show unique characters, main inflorescence axis, primary and floral bracts, as well as sepals are red, and flowers have pink petals (Figures 1-3, 5). Furthermore, a detail examination of the staminate flowers revealed the presence of stamens with filaments of two lengths, three epipetalous, with the short filaments attached at mid length of the petal, and three alternating with petals with filaments connate to the base of them (vs. two filaments of different lengths but attached to each petal; see Figure 2). These structures were cited already by Smith & Till (1998) for some Hechtia species. An additional feature of this new species, when compared with H. glomerata, is the absence of floral fragrance compared with the sweet diurnal fragrance on flowers of the latter. Floral fragrance is a common feature on Hechtioideae: a sweet diurnal smell perceptible by the human nose, but imperceptible just in few species, especially those with red sepals and/or petals, such as H. rosea E. Morren ex Baker, H. meziana L.B. Sm., H. iltisii Burt-Utley & Utley, and the new taxon here described. Preliminary studies (Ramírez-Morillo et al. 2022) characterizing nectar composition on species with different pollination syndromes (melittophily and ornithophily) in Hechtia (H. jaliscana L.B. Sm. and H. rosea, respectively) show differences supporting the hypothesis of two different syndromes in the genus.
One of the most conspicuous features of Hechtia sanchezii, is that male inflorescences continuously produce flower buds as the basal flowers open, which causes a long flowering period and the continuous elongation of the branches, something uncommon in the genus, where branches reach their maximum length, produce all the buds and then, just after that, blooming begins. This phenomenon has also been observed in species in the Hechtia glomerata complex native to Tamaulipas, which have not yet been described.

