











 nova página do texto(beta)
nova página do texto(beta) Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO 
 Permalink
Permalink
Los bosques tropicales en México abarcaban aproximadamente el 27 % del territorio nacional, de los cuales alrededor del 50 % correspondían a bosques tropicales caducifolios (BTC) y 11 % a bosques tropicales subcaducifolios (BTSC; Rzedowski 1991). Actualmente, el BTC y BTSC sólo representan el 16 %, principalmente como bosques tropicales en sucesión o secundarios (Challenger & Soberón 2008). Los BTC y BTSC son comunidades vegetales formadas en su mayoría por especies tropicales que se restringen principalmente a cerros o laderas con suelos someros, aunque suelen presentarse especies de afinidad xérica en suelos más profundos (Rzedowski 1978, Lott & Atkinson 2010) y especies de afinidad mésica en cañadas y arroyos (Trejo 2010). Las comunidades en su mayoría se caracterizan por la pérdida de hojas durante la estación seca, aun cuando hay especies dentro de estos bosques que se mantienen siempre verdes (e.g., Pithecellobium dulce; Rzedowski 1978). Algunas especies pierden sus hojas por solo unas semanas y al final de la estación seca (e.g., Lonchocarpus sp; Arreola-Villa 2012) y otras especies que tienen fenología invertida, es decir, producen sus hojas en la estación seca (e.g., Coccoloba liebmannii;Arreola-Villa 2012). Los BTC y BTSC se distribuyen principalmente en suelos delgados y pedregosos con buen drenaje en laderas de colinas en elevaciones de 0 a casi 2,300 m (Rzedowski 1978, Rzedowski & Calderón de Rzedowski 2013). Se estima que este tipo de bosques cuenta con un poco más de 11,000 especies vegetales (Challenger & Soberón 2008) de las 23,500 especies que se encuentran en México (Villaseñor 2016). El 73 % de las especies en el BTC y BTSC son endémicas al país (Banda-R et al. 2016), con un 25 % a nivel de géneros y 45 % a nivel de especies (Rzedowski 1998, Challenger & Soberón 2008).
En los últimos 25 años, se ha generado información sobre los tres elementos básicos de la diversidad, es decir, la riqueza, la abundancia y la composición de los BTC y BTSC de México. A pesar de las variaciones en los diseños de muestreo y los análisis de diversidad utilizados, el número de especies ha sido la medida básica, intuitiva y directa de la diversidad en varios de los estudios (e.g.,Trejo & Dirzo 2002, Pineda-García et al. 2007, Méndez-Toribio et al. 2014 ), mientras que la distribución de la abundancia de las especies ha sido destacada en otros (e.g.,Trejo 1998, Pineda-García et al. 2007, Dzib-Castillo et al. 2014, Méndez-Toribio et al. 2014, Mesa-Sierra et al. 2020). Algunos estudios han estimado índices de entropía para explicar los patrones de diversidad alfa (e.g.,Dzib-Castillo et al. 2014, Méndez-Toribio et al. 2014). La diversidad alfa de una comunidad se define como el número total de especies y la distribución de sus abundancias a nivel local, es decir, dentro de un sitio determinado (Whittaker et al. 2001, Magurran 2004, Cultid-Medina & Escobar 2019). En general, los BTC y BTSC de México presentan una alta diversidad alfa, es decir, un elevado número de especies que destacan por representar una gran variedad de géneros y de especies endémicas, los cuales han sido explicados por procesos locales de diversificación, especialización fenológica y una amplia gama de condiciones ambientales derivadas de la altitud, la litología, la exposición, la pendiente y la topografía (Balvanera et al. 2002, Trejo & Dirzo 2002, Durán et al. 2006, Lott & Atkinson 2010). Recientemente, se ha hecho énfasis sobre el efecto negativo del disturbio sobre la diversidad alfa; por ejemplo, las actividades pastoriles, como el pastoreo libre del ganado vacuno, disminuye la presencia y abundancia de plántulas de las especies vegetales. El aprovechamiento de la madera reduce la presencia y abundancia de los individuos de mayores diámetros y alturas, mientras que las plantaciones o cultivos frutales (e.g., mango, sandia, pepino y maíz) han provocado la remoción de la vegetación natural con la consecuente pérdida de especies a nivel local (Lebrija-Trejos et al. 2010, Martínez-Cruz et al. 2013, Cervantes-Gutiérrez et al. 2017).
Actualmente, la diversidad alfa de una comunidad suele evaluarse con índices que consideran el número total de especies en combinación con la distribución de sus abundancias (Cultid-Medina & Escobar 2019). Esta diversidad suele evaluarse con índices basados en entropía, como el índice Shannon, índice de Margalef, entre otros; sin embargo, estos índices no ofrecen una descripción intuitiva de la diversidad de la comunidad debido a que sus unidades no pueden ser interpretadas de forma directa en términos biológicos o ecológicos (Cultid-Medina & Escobar 2019). Por tal motivo, se ha propuesto caracterizar y comparar la diversidad a a partir de la serie de Hill (Hill 1973) o los índices de diversidad de orden q (qD; Jost 2006), debido a que estos índices comparan la diversidad en número efectivo de especies, lo que resulta en medidas más intuitivas para evaluar las comunidades biológicas (Moreno et al. 2011). Además, estos índices mantienen propiedades matemáticas que permiten reconocer las diferencias de magnitud entre comunidades. Entre las propiedades más importantes se destaca la propiedad de duplicación (Hill 1973, Jost 2006, Moreno et al. 2011) o también llamada principio de replicación (Jost 2007, Moreno et al. 2011), la cual hace referencia a que la suma de dos comunidades con el mismo número de especies, igual abundancia y sin especies compartidas será exactamente el doble de cada comunidad inicial (Chao & Jost 2012, Cultid-Medina & Escobar 2019). Por lo tanto, estos índices detectan de forma explícita cualquier diferencia o cambio en la diversidad de las comunidades, mientras que los índices clásicos basados en entropía no lo hacen, debido a que no cumplen con esta propiedad (Moreno et al. 2011).
Por otra parte, la diversidad beta se ha estudiado en los BTC y BTSC de México usando distintos métodos e índices. La diversidad beta es entendida como el recambio direccional en la composición de especies a lo largo de un gradiente ambiental, espacial o temporal (Whittaker 1975, Anderson et al. 2011) o como la variación no direccional en la composición de especies dentro de un área geográfica de interés (Legendre et al. 2005, Anderson et al. 2011). El análisis de los datos provenientes de 20 localidades de BTC en México mostró que el recambio de especies a lo largo del territorio es muy alto, es decir, que de las 917 especies, cerca de 72 % sólo se encontraron en un sitio, con una muy baja similitud florística entre estas áreas (Trejo & Dirzo 2002). Los patrones de diversidad beta en BTC y BTSC están explicados tanto por la heterogeneidad ambiental (como disponibilidad de agua, pendiente y exposición) como por la estructura espacial inherente, lo que promueve diferentes microhábitats que generan respuestas diferenciales en las especies (Balvanera et al. 2002, Trejo & Dirzo 2002, Gallardo-Cruz et al. 2010, Hernández-Stefanoni et al. 2012, López-Martínez et al. 2012). Actualmente, se ha hecho evidente que factores como las actividades pastoriles, el aprovechamiento de madera, el establecimiento de plantaciones o cultivos, la edad sucesional, la fragmentación y las propiedades del suelo pueden también influir en los patrones de diversidad beta en estos bosques (Lebrija-Trejos et al. 2010, López-Martínez et al. 2013, Cervantes-Gutiérrez et al. 2017). Estos factores modifican el ambiente local y disponibilidad de recursos, creando así oportunidades para el establecimiento de nuevas especies y reduciendo las poblaciones de las que ya están establecidas.
La diversidad beta suele evaluarse con índices tradicionales de disimilitud basados en el número de especies compartidas y el número de especies exclusivas a cada sitio (Koleff et al. 2003, Calderón-Patrón et al. 2012, Calderón-Patrón & Moreno 2019). Sin embargo, se ha propuesto la necesidad de dividir la disimilitud en dos componentes aditivos: la disimilitud derivada del recambio (reemplazo) de especies y la disimilitud derivada de las diferencias en el número de especies (anidamiento) entre los sitios (Baselga 2010, 2012). El enfoque de partición de la diversidad beta ayuda a entender los patrones de diversidad a partir de los mecanismos que la originan. El recambio (βsim) se genera por la sustitución de algunas especies por otras, como consecuencia de restricciones ambientales, espaciales o históricas. El anidamiento (βsne), conlleva a que el sitio más pobre en riqueza sea un subconjunto del sitio más rico, a causa de la pérdida (o ganancia) de especies en algunos de los sitios (Baselga 2010, 2012). La partición de la diversidad beta ha permitido detectar regiones con alto recambio de especies donde los índices tradicionales de disimilitud enmascaraban los efectos de las simples diferencias en riqueza entre sitios (Baselga 2012, Carvalho et al. 2012, 2013, Cardoso et al. 2015). Este nuevo enfoque de partición de la diversidad beta ha sido utilizado para la evaluación de la diversidad en paisajes modificados por humanos; Conradi et al. (2017) evaluaron cómo la ordenación ambiental, los procesos pasados y presentes regularon el recambio y anidamiento de las especies vegetales en pastizales; Alignier & Baudy (2016) determinaron los cambios en el recambio y el anidamiento de las comunidades vegetales en paisajes agrícolas, en diferentes períodos de tiempo y con distintas prácticas de manejo (pastoreo y fumigación con herbicidas); mientras que Heydari et al. (2017) evaluaron los efectos de gradientes de perturbación (severidad del fuego) sobre la diversidad alfa y beta (recambio y anidamiento).
A pesar de que Sierra de Vallejo y áreas circundantes presentan 10 tipos de ecosistemas terrestres, los cuales albergan 1,134 especies de plantas (CONANP 2012), el área no cuenta con estimaciones de diversidad alfa y beta. Los estudios se han enfocado en mostrar la composición de especies a partir de listados florísticos (Ramírez-Delgadillo & Cupul-Magaña 1999, Tejero-Diez et al. 2008, Gómez-Flores 2011) y la estructura de las comunidades vegetales de la zona costera (Bravo-Bolaños et al. 2013, 2016). Ante tal situación, el objetivo de este estudio fue evaluar la diversidad florística local y regional del BTC y BTSC de Sierra de Vallejo, Nayarit. La diversidad alfa o local se evaluó en términos de riqueza de especies y número efectivo de especies leñosas; mientras que la diversidad beta o regional se analizó en términos de similitud, recambio y anidamiento de las especies leñosas. Con base en el objetivo del estudio se propusieron cuatro predicciones. Primero, se espera que el número de especies por sitio de estudio se encuentre entre las 42 a 103 especies, riqueza específica reportada en los BTC de Jesús María, Nayarit (Trejo & Dirzo 2002) y Chamela, Jalisco (Lott et al. 1987), respectivamente. En segundo lugar, se espera que todos los sitios sean bastante homogéneos en cuanto al número efectivo de especies comunes. En tercer lugar, se espera que todos los sitios tengan un bajo número efectivo de especies dominantes. Las predicciones sobre la diversidad alfa se basan en la amplia heterogeneidad ambiental que caracteriza a los BTC y BTSC, lo que genera respuestas de las especies a variaciones sutiles pero continuas en el ambiente físico (Whittaker 1975, Balvanera et al. 2002, Trejo & Dirzo 2002, Gallardo-Cruz et al. 2009, Lott & Atkinson 2010). Además, en Sierra de Vallejo se han reportado actividades agropastoriles intensivas que afectan la riqueza de las especies y favorecen la presencia de especies leñosas dominantes (CONANP 2012). A partir del estudio de Suárez-Torres (2020), estas actividades se han podido catalogar como eventos de perturbación o disturbio crónico en algunas áreas dentro del polígono de estudio. Finalmente, se espera que la diversidad beta total esté explicada principalmente por el recambio o reemplazo de especies entre sitios, con una baja similitud en su composición florística. Esta predicción está basada en la heterogeneidad de condiciones (Balvanera et al. 2002, Trejo & Dirzo 2002, Gallardo-Cruz et al. 2010) y la estructura espacial inherente que caracteriza a estos bosques (Hernández-Stefanoni et al. 2012, López-Martínez et al. 2012), lo que promueve diferentes microhábitats que generan respuestas diferenciales en las especies. En este contexto, estudios que consideren los nuevos enfoques de índices de diversidad verdadera y partición de la diversidad beta pueden mejorar la comparación de los valores de diversidad a diferentes escalas espaciales y temporales, sin incurrir en los sesgos presentes en los índices tradicionales que se han usado para cuantificar la diversidad alfa y beta (Baselga 2010). Se espera que la información generada en este estudio sirva como referencia para conocer el estado actual de la diversidad local y regional del BTC y BTSC en Sierra de Vallejo, área de estudio que forma parte de la propuesta a declaratoria como Área Natural Protegida de Recursos Naturales Sierra de Vallejo-Río Ameca.
Materiales y métodos
Área de estudio. Los sitios con BTC y BTSC estudiados se localizan en Sierra de Vallejo (Figura 1). La Sierra se encuentra en los municipios de Compostela y Bahía de Banderas en la zona sur del estado de Nayarit (INEGI 2019), localizada en la provincia biogeográfica Costa Pacífica Mexicana (Morrone 2019). El área de estudio comprende una superficie de 1,018.5 km2 y se encuentra entre las coordenadas extremas 20 ° 08-21 ° 08´ latitud N y 104 ° 97-105 ° 17 longitud O, como parte del Área de Protección de Recursos Naturales Sierra de Vallejo-Río Ameca (CONANP 2012).
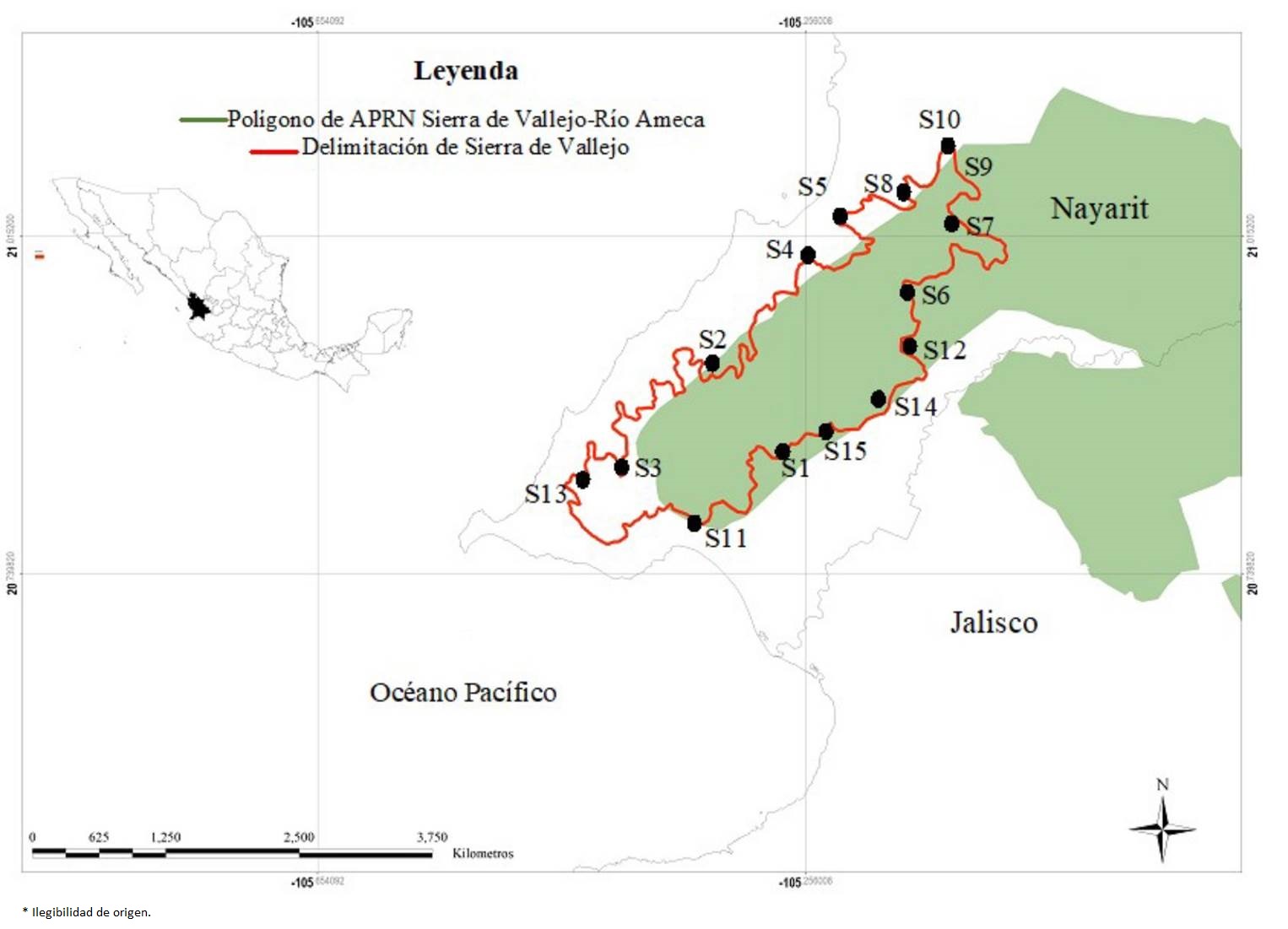
Figura 1 Área de estudio Sierra de Vallejo como parte del Área de Protección de Recursos Naturales, Sierra de Vallejo-Río Ameca.
Dentro de Sierra de Vallejo, la agricultura y la ganadería son las principales actividades antrópicas que tienen efectos negativos sobre la diversidad de los BTC y BTSC (Gómez-Mora et al. 2005, Suárez-Torres 2020). Las tierras agrícolas cubren el 10.3 % del área de estudio, en las cuales se suele cultivar principalmente maíz, chile, frijol, sandía y yaca (González et al. 2014, Suárez-Torres 2020). Asociado a esto, el uso de agroquímicos modifica la calidad de los suelos a largo plazo (Nava-Cruz 2006). La ganadería comprende principalmente el pastoreo libre del ganado vacuno; solo en el 2 % del área de estudio se desmonta el bosque para el cultivo de pastizales con importancia forrajera (Suárez-Torres 2020). En algunas zonas de la parte media y alta de Sierra de Vallejo se realiza la extracción de maderas para su uso local, como amapa (Handroanthus chrysanthus), capomo (Brosimum alicastrum), huanacaxtle (Enterolobium cyclocarpum), higuera (Ficus sp.) y papelillo (Bursera simaruba; González et al. 2014).
Muestreo de vegetación. Para el estudio se eligieron 15 sitios dentro del BTC y BTSC presente en Sierra de Vallejo (Tabla 1). Los criterios para seleccionar los sitios fueron: i) una distancia mínima de 3 km entre sitios seleccioandos, ii) sitios solo con vegetación de BTC y B TSC, descartando sitios que presentaron vegetación secundaria o de sucesión, y iii) sitios de fácil acceso. En cada sitio se estableció un transecto de 20 × 50 m (0.1 ha), en el cual se registró cada planta leñosa con un diámetro a la altura del pecho (a 1.30 m, DAP) > 3 cm. En el caso de los individuos no identificados en campo, se tomaron fotografías digitales y se colectaron muestras para su posterior cotejo con los ejemplares del Herbario Luz María Villarreal de Puga (IBUG) del Centro Universitario de Ciencias Biológicas y Agropecuarias (CUCBA) de la Universidad de Guadalajara, Zapopan, Jalisco.
Tabla 1 Características geográficas y tipos de vegetación de los 15 sitios censados en Sierra de Vallejo, Nayarit. BTC = bosque tropical caducifolio y BTSC = bosque tropical subcaducifolio.
| Clave del sitio | Población más cercana al sitio |
Municipio | Localización geográfica | Altitud (m snm) |
Tipo de vegetación |
|---|---|---|---|---|---|
| S1 | Valle de Banderas | Bahía de Banderas | 20 ° 49' 53.78" N; 105 ° 14' 48.67" O |
108 | BTSC |
| S2 | Úrsulo Galván | Compostela | 20 ° 55' 3.56" N; 105 ° 19' 43.92" O |
74 | BTSC |
| S3 | El Guamúchil | Bahía de Banderas | 20 ° 49' 27.94" N; 105 ° 22' 50.29" O |
394 | BTSC |
| S4 | La Peñita de Jaltemba | Compostela | 20° 59' 56.50" N; 105° 15' 45.58" O |
95 | BTC |
| S5 | La Peñita de Jaltemba | Compostela | 21° 3' 23.17" N; 105° 13' 6.57" O |
85 | BTSC |
| S6 | Las Cañadas | Compostela | 20° 59' 34.35" N; 105° 7' 55.63" O |
320 | BTSC |
| S7 | Cardoncillos | Compostela | 21° 1' 30.90" N; 105° 8' 48.02" O |
386 | BTSC |
| S8 | Altavista | Compostela | 21° 4' 2.82" N; 105° 9' 59.77" O |
169 | BTSC |
| S9 | Altavista | Compostela | 21° 4' 22.37" N; 105° 8' 48.01" O |
295 | BTSC |
| S10 | Altavista | Compostela | 21° 6' 7.30" N; 105° 8' 37.81" O |
419 | BTSC |
| S11 | Valle de Banderas | Bahía de Banderas | 20° 47' 18.40" N; 105° 17' 54.89" O |
101 | BTC |
| S12 | Fortuna de Vallejo | Bahía de Banderas | 20° 55' 48" N, 105° 07' 53" O |
142 | BTSC |
| S13 | San Quintín | Bahía de Banderas | 20° 48' 55.64" N; 105° 24' 44.03" O |
251 | BTC |
| S14 | El Colomo | Bahía de Banderas | 20° 53' 3.48" N; 105° 9' 3.36" O |
95 | BTSC |
| S15 | El Colomo | Bahía de Banderas | 20° 51' 46.56" N; 105° 11' 1.50" O |
92 | BTSC |
La distribución geográfica de las especies identificadas se categorizó considerando a Villaseñor (2016) y la base de datos digital Naturalista (www.naturalista.mx), considerando las siguientes categorías:
Endémica de la región.- son aquellas especies que están restringidas a una región (para este estudio los estados de Jalisco-Nayarit) y por lo tanto no se pueden encontrar en algún otro sitio.
Endémica de México.- se refiere a aquellas especies que tienen un área de distribución bien identificada y restringida a un territorio (para este estudio fue México; Ramírez-Marcial et al. 2003).
Nativa de México.- son aquellas que crecen de forma natural en una región, sin la intervención ni la ayuda del hombre. Se desarrollan de forma silvestre y solo se encuentran en determinados sitios (incluye aquellas especies cuya distribución geográfica abarca Norteamérica, Mesoamérica o Sudamérica; Ramírez-Marcial et al. 2003).
Exótica.- son aquellas que no tienen su origen dentro de la región donde actualmente se encuentran y su presencia se debe a que fueron traídas directamente por el hombre o que han llegado por dispersión de otro sitio de manera accidental (incluye aquellas que su origen y distribución geográfica es Europa, Asia, África, Oceanía y Sudamérica; Ramírez-Marcial et al. 2003).
Diversidad alfa. La diversidad local se estimó en número efectivo de especies, es decir, q0, q1 y q2 que son las unidades de medición de la diversidad verdadera (Jost 2006). En estos índices, el orden de diversidad cero (q0) representa la diversidad de especies sin considerar la abundancia; el orden q1 es el número efectivo de especies considerando las abundancias observadas; y el orden q2 es el número efectivo de especies tomando en cuenta solo las especies con la mayor abundancia. La diversidad de los tres órdenes se estimó a una misma completitud de inventario (SC = 80 %; Chao & Jost 2012). La completitud del inventario tiene valores entre 0-100 expresado en porcentaje, el cual mide la proporción del número total de individuos en la comunidad que pertenecen a las especies representadas en la muestra (Chao & Jost 2012). Para determinar si existían diferencias entre los sitios para los tres órdenes de diversidad, se realizó un análisis de varianza parámetrico considerando los intervalos de confianza del 95 %, los cuales se construyeron con base en rutinas de interpolación y extrapolación con 100 aleatorizaciones, estandarizando a un mismo número de individuos (300 individuos) en el programa en línea iNEXT (Hsieh et al. 2016; chao.shinyapps.io/iNEXTOnline). En caso de encontrar diferencias significativas, se aplicó una prueba de rangos múltiples por pares de Tukey estandarizada (HSD) para identificar los sitios donde se presentan las diferencias. Los análisis se realizaron en PAST 4.02 (Hammer et al. 2001) y JMP 8.0 (SAS 2008).
Curvas rango-abundancia. Para visualizar los patrones de la riqueza, la estructura y la composición de las especies leñosas en los sitios se realizaron curvas de rango-abundancia, utilizando las abundancias totales de cada especie ordenadas de mayor a menor (Magurran 2004).
Partición de la diversidad beta. La diversidad beta (regional) de las especies leñosas de Sierra de Vallejo se evaluó con base en la propuesta de Baselga (2010, 2012), donde la diversidad beta total (β) es particionada en sus componentes de recambio (β jtu) y anidamiento (β jne). Esta propuesta define al recambio como la sustitución de algunas especies por otras como consecuencia de un arreglo espacial o ambiental. Mientras que el anidamiento ocurre cuando las especies de las comunidades con bajo número de especies, son un subconjunto de las especies de las comunidades con mayor riqueza de especies (Baselga 2007, 2010, 2012, Baselga et al. 2007). Para este análisis, se construyó una matriz de similitud con base en el índice de Jaccard, utilizando los datos de presencia y ausencia de las especies leñosas de cada uno de los sitios. Los análisis se realizaron en el programa R usando el programa beta.part (Baselga & Orme 2012, R Core Team 2017). Para calcular el anidamiento y el recambio para el conjunto de todos los sitios se utilizó la función beta.multi y para calcularlos entre pares de sitios se usó la función beta.pair (Baselga & Orme 2012).
Diversidad beta-análisis de similitud. Para este análisis se utilizó el índice de Jaccard, el cual expresa el grado en el que dos sitios son semejantes por las especies presentes en ellas. El índice considera la incidencia de las especies en cada sitio y el resultado se puede visualizar como la proporción o porcentaje de especies compartidas (Pielou 1975, Magurran 2004). Para visualizar la similitud entre sitios se realizó un análisis de clúster, que se construyó con el método de agrupamiento de pares con la media aritmética no ponderada (UPGMA). Para establecer agrupaciones significativas se realizó la prueba de SIMPROF con base en 1,000 permutaciones con un α = 0.05. Así mismo, para visualizar las relaciones de similitud de especies entre los sitios, se realizó un escalamiento multidimensional no métrico (NMDS) a partir de la matriz de similitud de Jaccard (Clarke & Gorley 2006). Los análisis de similitud y NMDS se realizaron en el programa PRIMER v6 + PERMANOVA® (Clarke & Gorley 2006, Anderson et al. 2008).
Resultados
Diversidad alfa. Se censaron un total de 1,556 individuos leñosos pertenecientes a 45 familias, 99 géneros, 126 especies y 46 morfoespecies, nueve de las morfoespecies se identificaron a nivel de familia (Apéndice 1). Las familias más abundantes fueron Fabaceae (21 especies) y Rubiaceae (15), mientras que 19 familias sólo contaron con una especie. Bursera simaruba fue la especie para la que se registró el mayor valor de importancia relativa (VI = 9.60 %) basado en su abundancia (111 individuos). Se registraron 22 especies endémicas a México, una especie endémica a la región de Jalisco-Nayarit, 84 nativas a México y cuatro exóticas. Por sitio, la riqueza específica varió de 13 a 37 especies y la abundancia de 44 a 229 individuos (Tabla 2).
Tabla 2 Diversidad alfa (índices de diversidad verdadera y cobertura de la muestra) y diversidad beta (recambio y anidamiento) para los sitios del bosque tropical caducifolio y subcaducifolio de Sierra de Vallejo. Los valores de los índices de diversidad verdadera se comparan a un valor de 80 % de cobertura de la muestra. q0= Número total de especies, q1 = Número efectivo de especies, q2 = Número efectivo de especies, βjtu = Componente de recambio de disimilitud de Jaccard, βjne = Componente de anidamiento de la disimilitud de Jaccard. Los valores de las medias ± intervalos de confianza del 95 % con diferente letra indican diferencias estadísticamente significativas, según la prueba de Turkey HSD (P ≤ 0.05).
| Clave del sitio | Composición florística | Diversidad verdadera | Diversidad beta | |||||
|---|---|---|---|---|---|---|---|---|
| Abundancia | Especies | Familias | q0 | q1 | q2 | βjtu (%) | βjne (%) | |
| S1 | 72 | 17 | 11 | 11 ± 2efg | 7 ± 1cde | 5 ± 1de | 87 | 2 |
| S2 | 93 | 20 | 7 | 15 ± 2bcdef | 12 ± 1bcd | 11 ± 2abc | 90 | 1 |
| S3 | 84 | 15 | 13 | 7 ± 1g | 4 ± 1e | 3 ± 1e | 86 | 3 |
| S4 | 142 | 37 | 22 | 27 ±3a | 19 ±3a | 14 ± 3a | 87 | 4 |
| S5 | 82 | 17 | 11 | 11 ± 2defg | 10 ± 1cde | 8 ± 1bcde | 87 | 3 |
| S6 | 90 | 19 | 14 | 10 ± 2fg | 7 ± 2cde | 6 ± 1cde | 88 | 2 |
| S7 | 74 | 23 | 19 | 19 ± 4bc | 11 ± 3bcd | 7 ± 2bcde | 86 | 3 |
| S8 | 100 | 19 | 15 | 11 ± 2defg | 6 ± 2de | 4 ± 1e | 84 | 3 |
| S9 | 229 | 27 | 15 | 13 ± 1bcdefg | 1 ± 1cde | 6 ± 1cde | 82 | 4 |
| S10 | 177 | 33 | 15 | 20 ± 2b | 14 ± 2ab | 12 ± 2ab | 83 | 4 |
| S11 | 92 | 23 | 12 | 17 ± 2bcde | 13 ± 3abc | 10 ± 2abcd | 84 | 3 |
| S12 | 125 | 22 | 11 | 13 ± 2bcdefg | 10 ± 2bcde | 7 ± 1bcde | 84 | 3 |
| S13 | 104 | 23 | 17 | 14 ± 2bcdef | 8 ± 2cde | 5 ± 2cde | 83 | 3 |
| S14 | 47 | 13 | 12 | 13 ± 3cdefg | 9 ± 2cde | 6 ± 2cde | 83 | 3 |
| S15 | 44 | 14 | 11 | 18 ± 4bcd | 13 ± 3abc | 9 ± 4abcde | 92 | 1 |
A una cobertura de muestra del 80 %, los análisis entre sitios mostraron diferencias para el índice de diversidad de orden q0 (F14,30 = 14.38, P < 0.0001). El sitio S4 presentó el valor más alto con respecto al resto de los sitios (P ≤ 0.05, Tukey HSD), con un número efectivo de 27 ± 3 especies, mientras que el sitio S3 obtuvo siete especies siendo este el valor más bajo registrado (Figura 2, Tabla 2). La diversidad de orden q1 mostró diferencias entre sitios (F14,30 = 9.46, P < 0.0001). El S4 obtuvo 19 ± 3 especies que son consideradas efectivas, mientras que el sitio S9 obtuvo una especie siendo este el valor más bajo registrado (Figura 2, Tabla 2). Para la diversidad de orden q2, los análisis entre sitios mostraron diferencias (F14,30 = 7.54, P < 0.0001), el sitio S4 presentó 14 ± 3 especies efectivas, mientras el valor más bajo registrado fueron los sitios S3 con solo tres especies efectivas (Figura 2, Tabla 2).
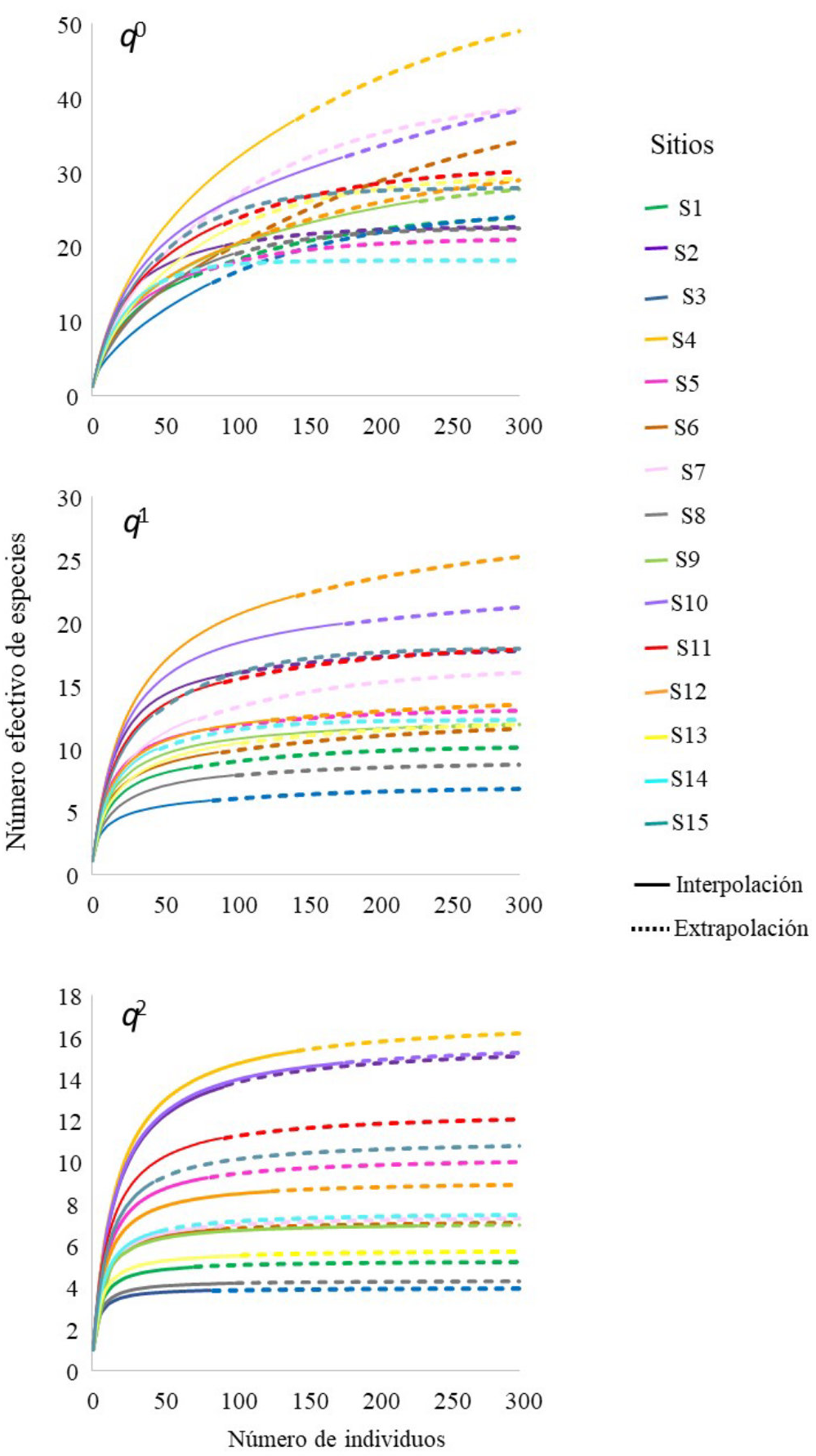
Figura 2 La diversidad verdadera o número efectivo de especies en los sitios de bosque tropical caducifolio y subcaducifolio de Sierra de Vallejo.
La distribución y la dominancia de las especies leñosas de Sierra de Vallejo fue distinta entre los sitios (Figura 3). Las especies que presentaron más abundancia en S1 fue Urera corallina (29 individuos), en S2 fue Heliocarpus occidentalis (12 individuos), en S3 fue Ardisia revoluta (33 individuos), en S4 la especie exótica Hibiscus tiliaceus (20 individuos), en S5 fue B. simaruba, en S6 fue A. revoluta (23 individuos), en S7 fue la Morfoespecie 50 (26 individuos), en S8 fue la Morfoespecie 22 (45 individuos), en S9 fue la Morfoespecie 50 (73 individuos), en S10 fue la Morfoespecie 25 (26 individuos), en S11 fue Lysiloma divaricatum (19 individuos), en S12 fue Eugenia salamensis (29 individuos), en S13 fue la especie endémica a México Cnidoscolus spinosus (40 individuos), en S14 fue Hura polyandra (15 individuos) y en S15 fue Guazuma ulmifolia (11 individuos). La especie nativa B. simaruba es la más frecuente, presentándose en 14 de los 15 sitios.
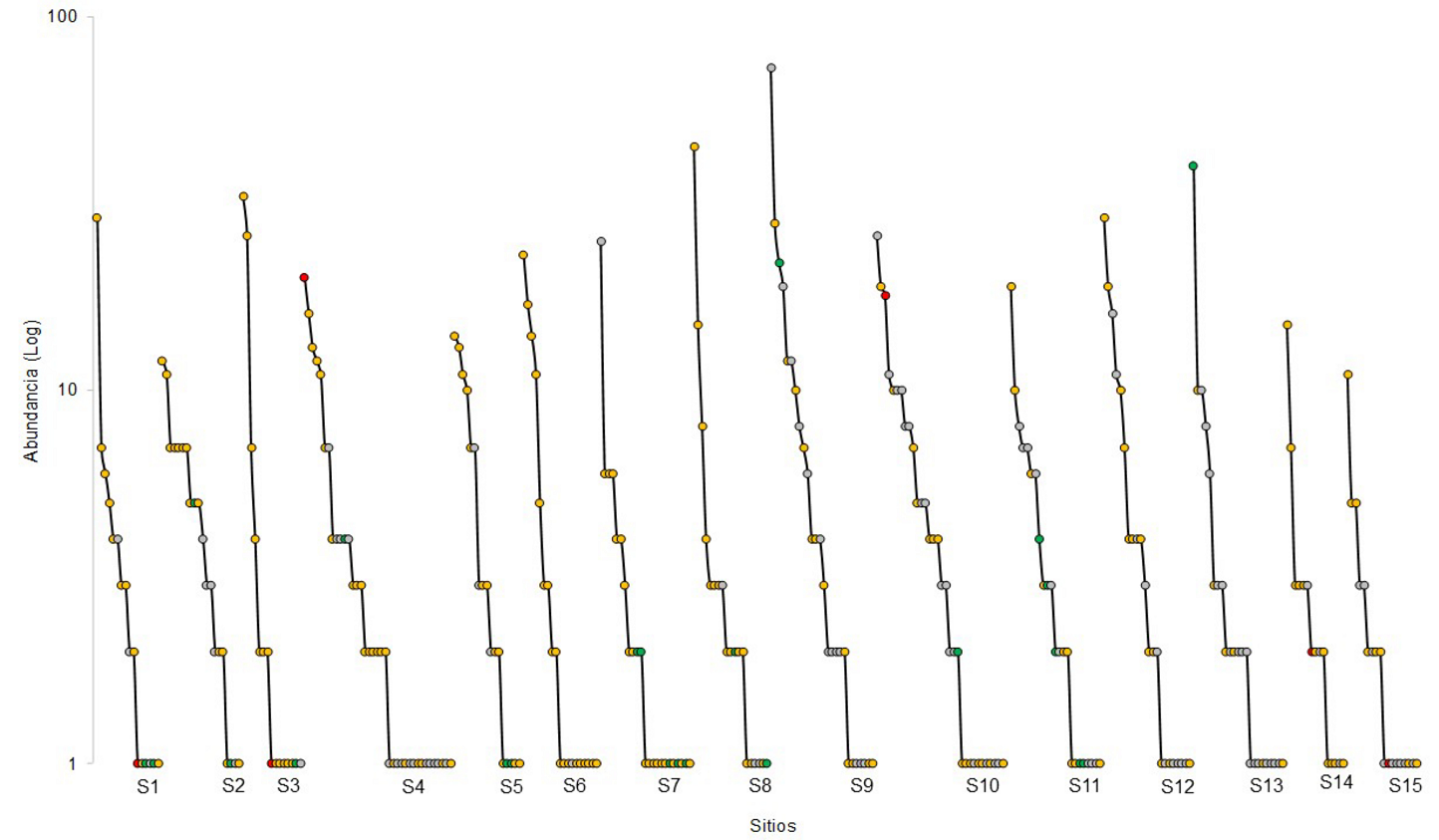
Figura 3 Curvas rango-abundancia de plantas leñosas en los sitios de bosque tropical caducifolio y subcaducifolio en Sierra de Vallejo. Los círculos muestran cada una de las especies por sitio. Los círculos verdes representan las especies endémicas de México, los amarillos a las especies nativas, los rojos a las especies exóticas y los grises a las morfoespecies. La letra S y número indican el sitio de estudio.
Diversidad beta. La diversidad beta total estuvo principalmente explicada por el recambio de especies entre sitios de BTC y BTSC de Sierra de Vallejo (Tabla 2). El índice de disimilitud de Jaccard muestra una beta total del 96 %, el recambio de especies presenta un valor alto (βjtu = 96) y un anidamiento bajo (βjne = 0.9). Considerando el índice de Jaccard, 15 sitios obtuvieron un valor > 80 % para el recambio de especies, mientras que los sitios S2 y S15 fueron los únicos que registraron > 90 % para el recambio de especies. Para el anidamiento, los valores fueron bajos, se registraron 12 sitios con un intervalo entre 1 a 3 %, mientras que los sitios S4, S9 y S10 registraron 4 % de anidamiento.
El análisis de similitud de Jaccard mostró que la similitud de especies entre los sitios de Sierra de Vallejo fue del 8 % (Figura 4). Los 15 sitios se agruparon en cinco grupos estadísticamente significativos (P < 0.05; SIMPROF), de acuerdo con la similitud en su composición de especies leñosas. El primer grupo está conformado por el sitio S13 con una similitud de 8 %, el segundo grupo compuesto por los sitios S15, S4, S11, S2, S1 y S5 con una similitud de especies del 13 %, el tercer grupo estuvo compuesto por S7 y S8 (37 %), el cuarto grupo estuvo conformado por los sitios S12, S9 y S10 (22 %) y el quinto por los sitios S14, S3 y S6 con una similitud 16 %. El análisis de escalamiento multidimensional no métrico mostró la ordenación de acuerdo la composición de especies entre los sitios, en donde se mostró la formación de cinco grupos (Figura 5), de acuerdo con la similitud en su composición de especies (Figura 4).
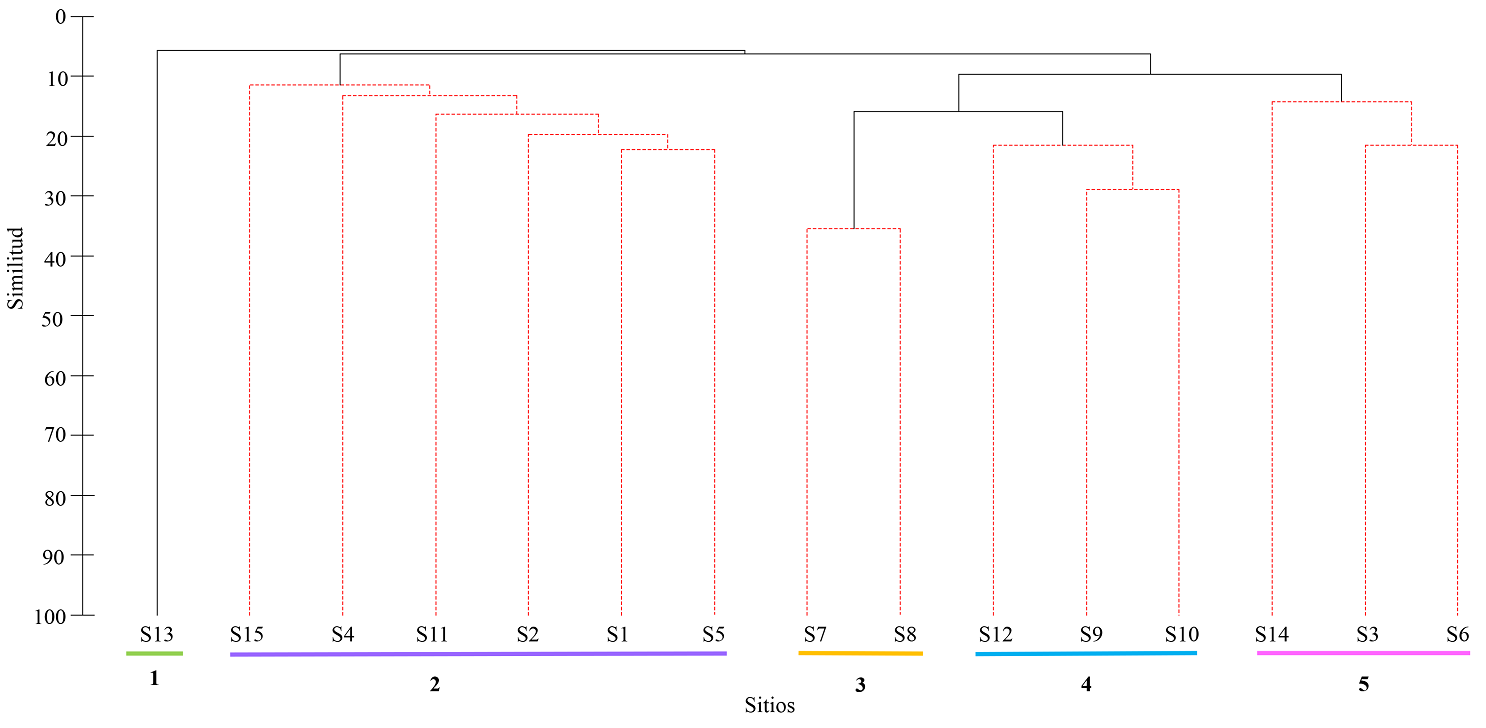
Figura 4 Dendrograma basado en el valor de disimilitud de Jaccard (0-100% similitud) de las especies para el bosque tropical caducifolio y subcaducifolio de Sierra de Vallejo. Las líneas rosas punteadas indican diferencias significativas (P < 0.05) a partir de la prueba de SIMPROF.
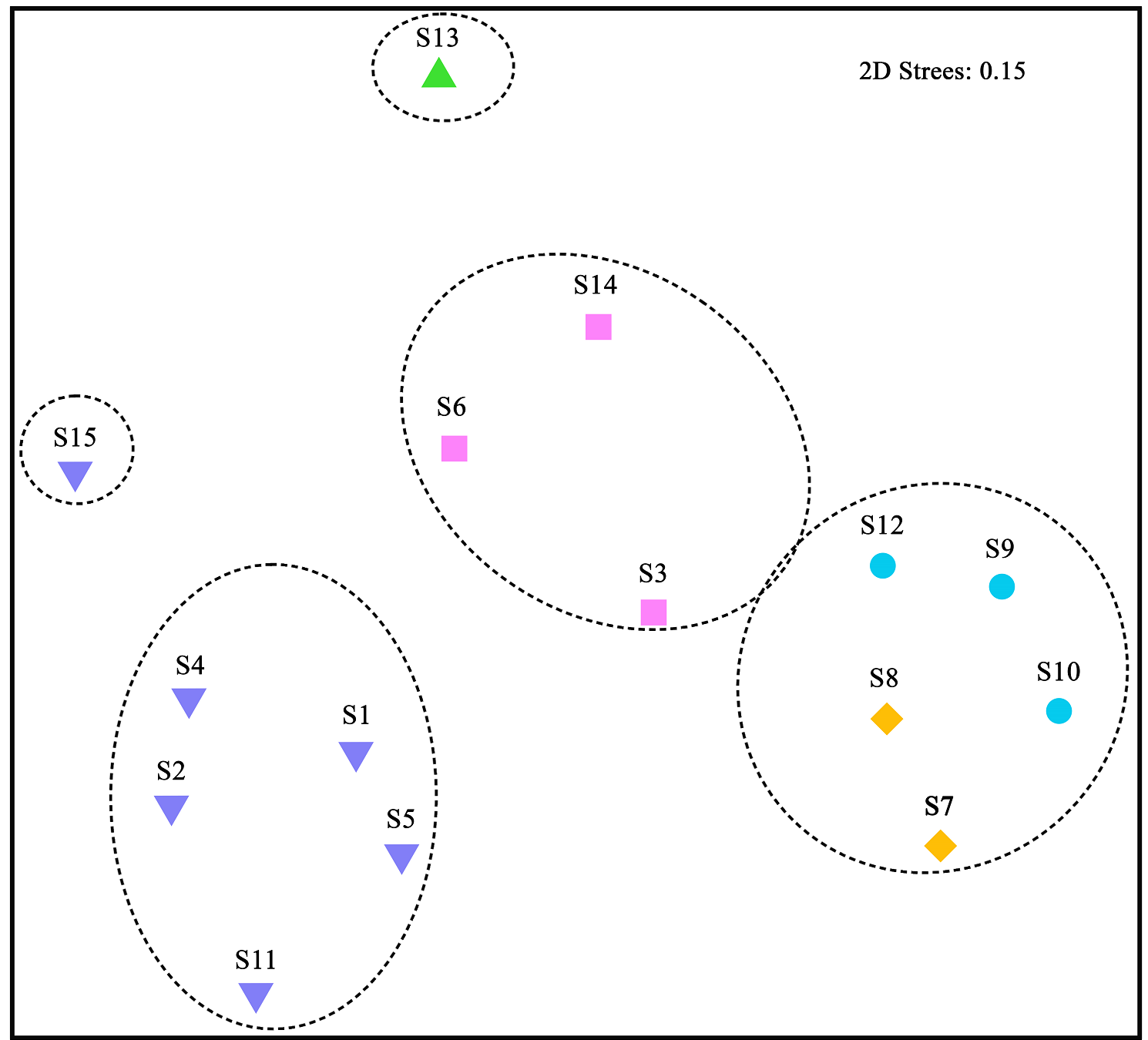
Figura 5 Análisis de escalamiento multidimensional no métrico (NMDS) para los 15 sitios y las 122 especies y 47 morfoespecies del bosque tropical caducifolio y subcaducifolio de Sierra de Vallejo. Los colores y símbolos representan a los cinco grupos identificados a partir del análisis de disimilitud de Jaccard. La letra S y número indican el sitio de estudio.
Discusión
La diversidad de plantas leñosas del BTC y BTSC de Sierra de Vallejo difiere notablemente a lo reportado para otros bosques tropicales estacionalmente secos de la costa del Pacífico Mexicano. En Sierra de Vallejo, el BTC y BTSC presentó un promedio bajo de riqueza específica (22 especies), en relación con lo reportado en Jesús María, Nayarit (42 especies, Trejo & Dirzo 2002), Chamela, Jalisco (103 especies, Lott et al. 1987), Cosalá, Sinaloa (58 especies, Trejo & Dirzo 2002) y Caleta, Michoacán (90 especies, Trejo & Dirzo 2002). La hipótesis planteada para la riqueza específica no se cumplió, debido a que el número de especies encontradas se mantiene muy por debajo del promedio reportado para bosques tropicales estacionalmente secos en México (58 especies; Trejo & Dirzo 2002). Los valores bajos de riqueza específica encontrados en Sierra de Vallejo pueden ser resultado del efecto de perturbación o disturbio crónico, principalmente por las actividades agropastoriles y el aprovechamiento de la madera para uso local, como ha sido reportado en estudios previos en otros BTC (Lebrija-Trejos et al. 2010, Martínez-Cruz et al. 2013, Cervantes-Gutiérrez et al. 2017). En los últimos 50 años, algunos de los sitios de estudio han presentado la actividad de libre pastoreo, la cual se realiza de cuatro a ocho meses al año (Suárez-Torres 2020). Otros trabajos han mostrado que los valores bajos de riqueza de especies en BTC y BTSC se deben al tipo de suelo (Durán et al. 2002), a los incendios forestales (Maass et al. 2010), a la falta de regulación de la carga de ganado y a las actividades extractivas (Martínez & Ceballos 2010, Martínez-Cruz et al. 2013), cuyo efecto no ha sido evaluado en este trabajo, pero que pueden estar moldeando la estructura, composición y riqueza de especies (Durán et al. 2002, Trejo & Dirzo 2002, Banda-R et al. 2016).
En términos florísticos, la especie más representativa y abundante fue Bursera simaruba, la cual define la estructura y composición de los 14 sitios en los que se presenta. Este resultado esta explicado por diversas características ecológicas. Esta especie es capaz de tolerar diversas condiciones ambientales, debido a que presenta una amplia plasticidad adaptativa (Gutiérrez & Ricker 2012), así como una amplia tolerancia ecológica, lo que la hace una especie resistente a los efectos de perturbación o disturbio crónico. B. simaruba es característica de los BTSC en México y de otros tipos de vegetación, como el bosque tropical perennifolio (Rzedowski 1978, Pineda-García et al. 2007, Pennington et al. 2009), lo que hace que su distribución no se encuentre limitada a los BTC (Lott et al. 1987, Lott & Atkinson 2010). Esta especie es utilizada frecuentemente como cerco vivo por las empresas ganaderas y propietarios locales (Bravo-Bolaños et al. 2013); además suele utilizarse como medicina, alimento, sombra y forraje para el ganado en la región de Nayarit (Villanueva et al. 1996), por lo que su abundancia alta dentro de Sierra de Vallejo se encuentra justificada. Así mismo, B. simaruba es una especie con alto valor ecológico y cuenta con número importante de estudios en México (Pompa-García et al. 2017).
En Sierra de Vallejo, se encontró que el 40 % de las especies leñosas encontradas son nativas de México, es decir, su distribución suele abarcar parte o la totalidad del territorio nacional. Este porcentaje de especies nativas de México es similar al reportado para la flora de Chamela-Cuixmala, en donde 42 % de las especies presentan un patrón de distribución Pacífico-Mesoamericano, es decir, a lo largo de toda la vertiente Pacífica desde Sonora hasta Costa Rica (Lott & Atkinson 2010). Lo anterior implica composiciones florísticas, a nivel de especies, muy similares entre el BTC y BTSC de Sierra de Vallejo y el BTC de Chamela-Cuixmala, lo que supone un proceso de diversificación a escala regional. El 12 % de las especies leñosas de los sitios de Sierra de Vallejo presentó endemismo a nivel de especie, similar al encontrado en otros BTC y BTSC de Guerrero y Michoacán, México (Pineda-García et al. 2007, Méndez-Toribio et al. 2014). En México cerca de 60 % de las especies de los BTC son endémicas al país (Lott & Atkinson 2010). Para Sierra de Vallejo, Inga andersonii fue la única endémica para la región Jaslico-Nayarit y otras 19 especies endémicas para el país, como Cryosophila nana, Erythrina lanata y Bursera roseana (Apéndice 1).
Por otra parte, se encontraron cuatro especies exóticas, Annona cherimola, H. tiliaceus, Coffea arabica y Citrus x limon. La presencia de estas especies puede estar explicada por el sistema silvopastoril que se encontró en los alrededores de algunos de los sitios o por la antigua vocación de los sitios estudiados (cafetales abandonados). Posiblemente, la presencia de estas especies exóticas puede ocasionar alguna alteración en la dinámica de las poblaciones de especies nativas dentro del bosque tropical de Sierra de Vallejo. Esta situación podría estar sucediendo en los sitios donde H. tiliaceus y C. arabica son las especies dominantes en abundancia. Sin embargo, se requieren estudios para evaluar el estatus en el que se encuentran el BTC y BTSC de Sierra de Vallejo para determinar si las especies leñosas exóticas están causando algún impacto a nivel local o regional.
Diversidad alfa. La utilización de índices de diversidad verdadera permitió estimar la diversidad de Sierra de Vallejo bajo un enfoque más intutitivo al considerar números efectivos de especies. Así mismo, la incorporación de técnicas de interpolación y extrapolación permitió comparar la diversidad de las comunidades del BTC y BTSC bajo una misma completitud de muestra. Para el índice q0, se obtuvo un número promedio efectivo de 15 especies, valor aún más bajo que el registrado para la riqueza promedio de especies observadas, lo que evidencia el efecto de incorporar la estructura de las comunidades en las estimaciones de diversidad. En México, no existen estudios en BTC y BTSC que evalúen la diversidad en número efectivo de especies leñosas. A la fecha, solo se encontró un estudio en fragmentos de BTC en Córdoba, Colombia (Ballesteros-Correa et al. 2019). En este estudio se reportan valores para el índice q0 de 80 a 110, con rangos de completitud de 94 a 98 %. Los valores de q0 del BTC colombiano son cuatro veces más grandes que los obtenidos en este estudio, a pesar de que son fragmentos de BTC asociados a sistemas de manejo silvopastoriles y ganadería extensiva convencial.
Para el índice q1, que considera el número efectivo de especies comunes y las abundancias de las especies equitativamente, los sitios obtuvieron rangos de uno a 19 especies efectivas. Se esperaba que los sitios fueran homogéneos en cuanto al número efectivo de especies comunes, por lo que la hipótesis para la diversidad de orden q1 no se cumplió, debido a que los sitios fueron bastante heterogéneos en valores. El sitio S4 fue el que obtuvo la mayor cantidad de especies comunes, y considerando los resultados de la gráfica de rango abundancia (Figura 3), algunas de estas especies podrían ser H. tiliaceus, Roseodendron donnell-smithii, Psidium sartorianum, B. simaruba, Thouinia acuminata, Cedrela odorata y A. revoluta. Para el sitio S10, que presenta la mayor cantidad de especies comunes, estas podrían ser Faramea occidentalis, C. arabica, Brosimum alicastrum y Pouteria sp. Los resultados de este estudio están muy por debajo de los reportados para los fragmentos de BTC colombiano, para los cuales se registraron de 33 a 44 especies comunes (Ballesteros-Correa et al. 2019). Una posible explicación para la diferencia tan marcada de valores del índice q1, podría deberse a las formas de vida consideradas en los censos de vegetación. En los fragmentos de BTC colombiano se incluyeron a los árboles, arbustos, herbáceas trepadoras y lianas, mostrando la importancia de considerar una mayor variedad de formas de vida en las estimaciones de la diversidad verdadera.
La hipótesis para el índice q2 planteaba que los sitios tendrían un bajo número efectivo de especies dominantes, lo cual no se cumplió. Los sitios de estudio en Sierra de Vallejo tuvieron valores altos de especies dominantes. Posiblemente, este resultado se debe al aprovechamiento y extracción selectiva de la madera para uso local, actividades realizadas durante los últimos 50 años en algunos de los sitios (Suárez-Torres 2020), así como el desmonte y transformación de la cobertura vegetal original, como se ha registrado en la vegetación de la zona costera (Bravo-Bolaños et al. 2013, 2016). El sitio S4 sigue el patrón de presentar los valores más altos de los 15 sitios estudiados, debido a que presentó un total de 14 especies dominantes. Este valor puede ser resultado del efecto de manejo que se le ha dado al sitio durante años (53 años) y la intensidad de uso (8 meses al año) de pastoreo libre del ganado vacuno (Suárez-Torres 2020), lo que puede estar favoreciendo la presencia de especies leñosas dominantes, entre las que destacan la exótica H. tiliaceus y las nativas R. donnell-smithii, P. sartorianum y B. simaruba. Por otro lado, el sitio S8 presentó de los valores más bajos de especies dominantes, con sólo cuatro especies dominantes y todas ellas nativas a México, entre las que se encuentran B. simaruba, Spondias mombin y Bixa orellana.
Actualmente, la diversidad alfa se mide más frecuentemente con los índices tradicionales de entropía en comparación con los índices de diversidad verdadera (Moreno et al. 2018). Este nuevo enfoque metodológico de "diversidad verdadera" expresado en número efectivo de especies (q0, q1 y q2), sigue siendo poco utilizado en comunidades vegetales y más aún en los BTC y BTSC, lo que dificultó la comparación de los resultados obtenidos en Sierra de Vallejo con respecto a otros bosques del mismo tipo en México o América Latina, siendo un área de oportunidad para futuros estudios.
Diversidad beta. En los sitios del BTC y BTSC de Sierra de Vallejo, la diversidad beta total muestra un valor alto de recambio (91-92 %), anidamiento bajo (0.9-2 %) y similitud baja (8 %). En este estudio se esperaba que la diversidad beta total estuviera explicada principalmente por el recambio o reemplazo de especies entre sitios, con una baja similitud en su composición florística, por lo que la hipótesis se cumplió. Los resultados del análisis de similitud muestran que los BTC y BTSC de Sierra de Vallejo coinciden con los patrones generales de diversidad beta reportados principalmente para el BTC en otras regiones de México, es decir, un alto recambio y bajo anidamiento de especies leñosas, generado principalmente por la heterogeneidad ambiental que existe en estos bosques (Trejo 1998, Balvanera et al. 2002, Hernández-Stefanoni et al. 2012, López-Martínez et al. 2012). Los próximos estudios de diversidad beta en Sierra de Vallejo podrán mostrar si el recambio es resultado de una selección de las especies por el ambiente (teoría de nicho) o si el recambio es resultado de procesos de dispersión de las especies (teoría neutral; Calderón-Patrón & Moreno 2019), como ya ha sido mostrado en los BTC de la Península de Yucatán (López-Martínez et al. 2013). Los autores mostraron que la diversidad beta de árboles y lianas está explicada por la teoría del nicho, mientras que la diversidad beta de los arbustos esta explicada por la teoría neutral.
El valor bajo de anidamiento del índice de Jaccard indicó que la distribución de las especies no es uniforme en los sitios de BTC y BTSC de Sierra de Vallejo. El anidamiento disminuye a medida que aumenta la heterogeneidad ambiental (Wright & Reeves 1992). Junto con la heterogeneidad ambiental se deben de considerar otros factores para explicar los valores de la partición de beta total, sobre todo considerando que los sitios de estudio sufren el efecto de perturbación o disturbio crónico, por las actividades agropastoriles y el aprovechamiento de la madera para uso local, así como la modificación de la cobertura vegetal original (Bravo-Bolaños et al. 2013, González et al. 2014, Bravo-Bolaños et al. 2016, Suárez-Torres 2020). En otros BTC de México, el recambio se ha relacionado con la edad sucesional (López-Martínez et al. 2012), así como con la separación entre sitios (estructura espacial) y el grado de perturbación (Villers-Ruiz & Trejo-Vázquez 1997, Trejo & Dirzo 2002), en donde estos factores son los mediadores de los procesos ecológicos en BTC. Los estudios indican que para México y América Latina no hay registros específicamente con los índices de disimilitud de Jaccard (recambio y anidamiento) para plantas leñosas en BTC y BTSC. Solo por fines de comparación, los resultados de este estudio son similares a los obtenidos en el bosque tropical tucumano-boliviano, de Tarija, Bolivia, donde obtuvieron el 6.5 % de anidamiento y 93.5 % de recambio (Gallegos et al. 2019). Sin embargo, la comparación debe de tomarse con cautela, debido a que en el estudio de Bolivia se consideran un mayor número de formas de vida (árboles, arbustos y lianas) y el bosque caducifolio es principalmente montano, con dosel de 10 a 30 m de altura.
Los sitios del BTC y BTSC de Sierra de Vallejo se congregaron en cinco grupos como resultado de la baja similitud (8 %) en la composición florística. La especie B. simaruba se presentó en cuatro de los grupos, con una contribución que va de 9 a 21 %, mientras que R. armata fue otra especie con un alto valor de importancia en dos de los grupos, con valores de 10 y 22 %. Ambas especies son nativas a México, con amplia distribución y tolerancia ecológica por lo que su distribución no se encuentra limitada a BTC y BTSC. El valor de similitud del BTC y BTSC de Sierra de Vallejo es consistente con un estudio realizado en 20 localidades en la vertiente del Océano Pacífico con BTC en México, con una similitud de 9% especies entre sitios (Trejo & Dirzo 2002). De manera complementaria, el análisis de escalamiento multidimensional no métrico, el cual está basado en la comparación de la composición de especies de cada sitio, muestra que los sitios más cercanos son aquellos que más especies comparten.
En Sierra de Vallejo se ha presentado la pérdida de hábitat y la fragmentación del BTC y BTSC, resultado de los años de perturbación y disturbio crónico que han sufrido algunos de los sitios y zonas aledañas (Bravo-Bolaños et al. 2013, González et al. 2014, Bravo-Bolaños et al. 2016, Suárez-Torres 2020). El pastoreo libre del ganado vacuno, el aprovechamiento de la madera para uso local y el desmonte de la cobertura vegetal original para el establecimiento de plantaciones y cultivos, son actividades antrópicas que han generado un bajo número de especies leñosas, un muy variable número efectivo de especies comunes y dominantes. Por otra parte, los patrones de diversidad beta siguen siendo explicados por la heterogeneidad ambiental, al obtener un alto recambio de especies, y por ende, una baja similitud entre los sitios. Sumado a esto, se registró la presencia de cuatro especies leñosas exóticas. Los resultados obtenidos en este estudio muestran como las nuevas aproximaciones metodológicas que incorporan atributos de las comunidades vegetales (en la diversidad verdadera) se podrían integrar con los valores de recambio y anidamiento de la diversidad beta total para generar una propuesta de conservación.
Finalmente, el presente estudio brinda información que puede orientar las estrategias de conservación de los BTC y BTSC en Sierra Vallejo. El alto recambio de especies que se evidencia sugiere que las propuestas para la conservación de la diversidad florística de los BTC y BTSC deben priorizar la protección de un mayor número de sitios, así como el de mantener la conectividad entre estos. Por otro lado, los nuevos proyectos en la zona deberán generar más información sobre otros grupos de organismos que habitan en los BTC y BTSC, información que dará soporte a la propuesta de decreto de Sierra de Vallejo-Río Ameca como un Área de Protección de Recursos Naturales (CONANP 2012).

