











 nova página do texto(beta)
nova página do texto(beta) Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO 
 Permalink
Permalink
México figura entre los cinco países con mayor diversidad vegetal en el mundo pues alberga alrededor de 23,314 especies de plantas vasculares, de las cuáles se calcula que el 50 % son endémicas del país (Villaseñor 2016). Se ha documentado que la riqueza florística no se distribuye de manera homogénea en el territorio mexicano, sino que responde a un patrón que se inicia en los estados de Chiapas y Oaxaca y se prolonga hacia las cadenas montañosas Sierra Madre del Sur, Sierra Madre Occidental, Faja Volcánica Transmexicana y la Sierra Madre Oriental (Rzedowski 1991a, b). Sin embargo, el endemismo vegetal en México no presenta el mismo patrón, ya que se concentra en las zonas áridas del país (Rzedowski 1991b, 1993, Sosa & De-Nova 2012, Salinas-Rodríguez et al. 2017, Sosa et al. 2018).
Una de las explicaciones más recurrentes a esta riqueza es la posición geográfica de México, en una zona orográfica compleja, en la que convergen las regiones biogeográficas Neártica y Neotropical. Esta zona ha sido definida como la Zona de Transición Mexicana (ZTM) (Halffter 2003). La ZTM está integrada por las provincias biogeográficas Sierra Madre Occidental (SMOC), Sierra Madre Oriental (SMOR), Sierra Madre del Sur (SMS), Eje Volcánico Transmexicano (EVT) y Tierras Altas de Chiapas (TC) (Morrone et al. 2017, Morrone 2019), que además representan áreas con la mayor diversidad vegetal del país (Sosa & De-Nova 2012, Villaseñor 2016). La SMOr junto con la SMS constituyen las cadenas montañosas más antiguas de México, con un origen estimado de 65 Ma, durante el paleoceno (Mastretta-Yanes et al. 2015). La primera está constituida casi en su mayoría por rocas sedimentarias marinas (Salinas-Rodríguez et al. 2017), mientras que la SMS es heterogénea, con un ensamble complejo de distintos tipos de roca de diferentes edades, que deriva en una vasta gama de tipos de suelos (Centeno-García 2004, Velasco de León et al. 2016). La SMS está conformada por cadenas montañosas disyuntas con dirección NO-SE que mantienen una orientación paralela al Océano Pacífico, desde Cabo Corrientes, en Jalisco, hasta el Istmo de Tehuantepec, en Oaxaca. Esta sierra limita con las provincias Cuenca del Balsas (CB), Costa Pacífica Mexicana (CPM), EVT y Veracruzana (V) (Morrone 2017, Aragón-Parada et al. 2019) y se considera que posee identidad propia, tanto en ecosistemas, como en composición biótica (Espinosa et al. 2016, Luna-Vega et al. 2016).
La SMS comprende una extensión de ca. 1,200 km y una elevación que varía de los 300 a 3,730 m. En esta accidentada topografía dos ríos importantes interrumpen su continuidad fisiográfica: i) el río Armería, en los límites de Colima y Jalisco y ii) el río Balsas, en los límites de Guerrero y Michoacán (Espinosa et al. 2016). La SMS se ha dividido en tres porciones: Jalisciense, Michoacana y Guerrerense-Oaxaqueña (Espinosa et al. 2016). Su compleja historia geológica y paleoclimática ha generado una heterogeneidad de paisajes y ambientes que han permitido la presencia de una elevada riqueza de especies, taxones endémicos, ecosistemas templados y transicionales con ecosistemas tropicales deciduos, xerófilos y perennifolios (Rzedowski 1978, Morrone 2001, Espinosa et al. 2016).
El concepto de endemismo propone formalizar la relación entre la evolución de un taxón con su área de distribución (Noguera-Urbano 2016, 2017). Comprender los patrones de endemismo es de suma importancia para definir sitios y/o áreas prioritarias para la conservación, pues son varios los atributos asociados a éstos, como la rareza local y global de los taxones, el aislamiento filogenético y la diversidad filogenética y funcional (Cracraft 1985, Forest et al. 2007, Crain et al. 2011). En consecuencia, el endemismo se relaciona con taxones restringidos a un área geográfica o que presentan un área de distribución reducida (Anderson 1994, Casagranda et al. 2009), la cual se entiende como el área del espacio geográfico donde un taxón está presente e interactúa en forma no efímera con el ecosistema (Zunino & Palestrini 1991) y una forma de representarla es por medio de la transformación de datos de puntos de ocurrencia en un mapa (Roig-Juñent et al. 2002).
Cerca del 50 % de las especies de plantas mexicanas se consideran endémicas (Villaseñor 2016). Las familias Asteraceae, Cactaceae Rubiaceae, Orchidaceae y Poaceae son las que tienen el mayor número de especies restringidas (Rzedowski 1991b). Este alto endemismo obedece a factores geológicos, edáficos, climáticos, fisiográficos y de aislamiento ecológico, que han permitido la diversificación y establecimiento de especies únicas en el territorio mexicano (Rzedowski 1991b). En el nivel nacional se han propuesto regiones que concentran altos valores de endemismo, como el matorral rosetófilo del noreste (Nuevo León-Tamaulipas), la Sierra Gorda (Querétaro-San Luis Potosí), la porción sur del Desierto Chihuahuense (Hidalgo), el Valle de Tehuacán-Cuicatlán (Oaxaca-Puebla), la Sierra de Quila (Jalisco), el área occidental de la Cuenca del Río Balsas (Guerrero-Morelos), el Istmo Tehuantepec (Oaxaca) y la Depresión Central de Chiapas (Sosa & De-Nova 2012, Salinas-Rodríguez et al. 2017, Sosa et al. 2018).
Dentro de la ZTM, la SMS concentra ca. 7,016 especies de plantas vasculares (Espinosa et al. 2016), pero aún se desconocen los taxones restringidos a esta provincia biogeográfica, el área que ocupan y los tipos de vegetación en los que se encuentran. Los objetivos de este trabajo son: 1) enumerar las plantas vasculares endémicas de la provincia biogeográfica SMS por familias, géneros, subprovincias y estados, 2) identificar las familias y géneros con mayor número de especies endémicas, 3) verificar si las especies enumeradas se encuentran bajo alguna categoría de riesgo y 4) reconocer en la SMS los tipos de vegetación más ricos en endemismo. Esta información, podrá incidir, junto con la riqueza florística, genética y ecosistémica, en proponer de manera más eficiente áreas prioritarias para la conservación.
Materiales y métodos
Área de estudio. La SMS tiene una extensión de 1,200 km, se ubica paralela al Océano Pacífico en dirección sur occidente de México, en los estados de Guerrero, Jalisco, Michoacán, Oaxaca y áreas marginales de Puebla y Veracruz, entre las provincias biogeográficas CB, CPM, EVT y V (Morrone et al. 2017). Dentro de ésta se reconocen tres subprovincias biogeográficas y cinco distritos (Morrone 2017). La Sierra Madre del Sur Occidental (SOC), incluye a los distritos Jalisco y Jalisco-Manantlán, con intervalos altitudinales que van de los 300 a los 2,890 m. La Sierra Madre del Sur Central (SCE) corresponde al distrito Michoacán y sus elevaciones varían entre 300 y 2,890 m. Finalmente, la Sierra Madre del Sur Oriental (SOR) se divide en los distritos de Guerrero y Oaxaca, con elevaciones que oscilan entre los 200 y 3,730 m. Se utilizó el polígono correspondiente a la SMS como provincia biogeográfica propuesto por Morrone (2017). Debido a que algunas montañas quedaban cortadas por el polígono, se decidió agregar una extensión de 2.5 km de amortiguamiento en los bordes de éste (Figura 1).
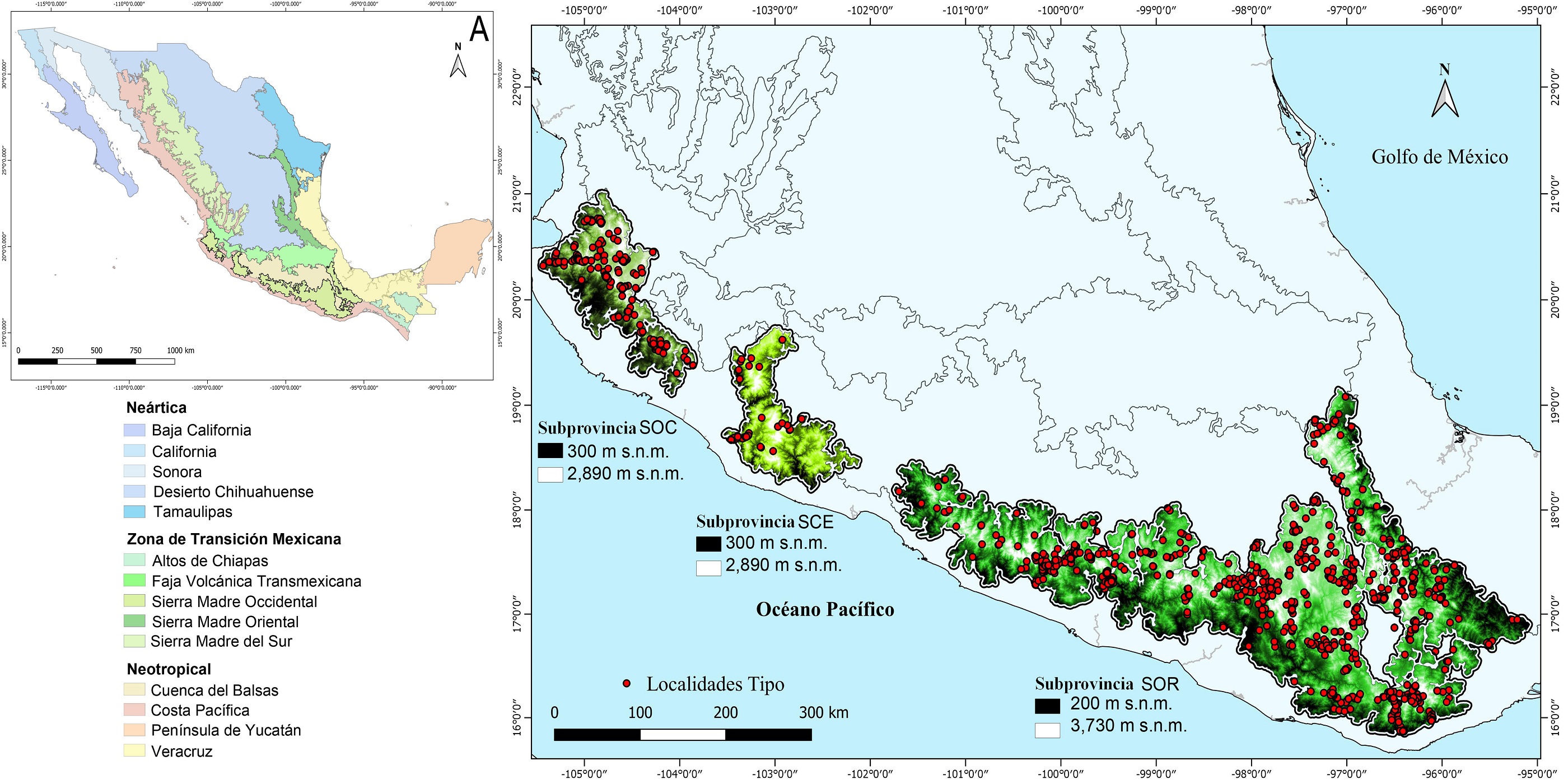
Figura 1 Provincias biogeográficas de México. A. Provincia biogeográfica Sierra Madre del Sur. B. Subprovincias biogeográficas Sierra Madre del Sur Occidental (SOC), Sierra Madre del Sur Central (SCE) y Sierra Madre del sur Oriental (SOR). (•) Localidades tipo de 1,133 especies endémicas de la SMS. Mapas de J. Aragón-Parada con base en Morrone et al. (2017).
Los tipos de vegetación reportados para la SMS son: bosque de coníferas y encino (4 7 %), bosque tropical caducifolio (24 %), bosque mesófilo de montaña (15 %) y, en conjunto (14 %), el bosque tropical perennifolio, bosque tropical subcaducifolio, matorral xerófilo, pastizal y otros tipos de vegetación (Rzedowski 1978, Santiago-Alvarado et al. 2016). Las montañas con mayor elevación en la SMS son el Cerro de las Capillas (2,890 m), en Jalisco, Las Conchas (2,890 m), en Michoacán, Teotepec (3,550 m), en Guerrero, Yatin (3,370 m), del Águila (3,370 m), Zempoaltépetl (3,420 m) y Nube Flandes (3,710 m), en Oaxaca.
Debido a la compleja orografía de la SMS, se presenta una gran diversidad de climas. Aunque su ubicación está dentro de los trópicos, su clima no es totalmente tropical, ya que se presentan variaciones desde climas lluviosos y cálidos, en la porción oriental de la SMS, hasta secos y fríos, en las montañas más altas. El clima dominante es el templado C, con amplia variación en humedad. También se presentan los climas templados húmedos C(f), C(m), C(m)(f) y subhúmedos C(w1), C(w2), C(w0), con menor extensión los cálidos, A, ya sea húmedos o subhúmedos y los semicálidos A(C) (Hernández-Cerda et al. 2016). Los niveles más altos de precipitación ocurren en las regiones más altas ubicadas en las laderas de barlovento, mientras que, en las de sotavento, el efecto Föehn produce corrientes de aire seco que generan temperaturas altas y baja precipitación (Hernández-Cerda et al. 2016).
Compilación de información. La información correspondiente a los taxones endémicos, su distribución geográfica, curatorial y ecológica se recopiló entre los años 2016 y 2020, a través de la consulta de especímenes en diferentes herbarios de México (CHAPA, ENCB, FCME, GUADA, HUAP, HUMO, HUNSIJ, IBUG, IEB, MEXU, OAX, SERO, UAGC, UAMIZ y ZEA), así como imágenes de especímenes resguardados en colecciones extranjeras (BH, CAS, F, K, MO y NY; acrónimos según Thiers 2019), que se consultaron en las páginas de la colección o a través del portal Global Plants (Global Plants 2020). También se integró información de bases de datos electrónicas como La Red de Información de Biodiversidad Global (CONABIO 2020) y de Global Biodiversity Information Facility (GBIF 2020). Así mismo se revisó extensamente literatura florística (e.g., Dávila et al. 2018, de Santiago-Gómez 1996, Diego-Escobar et al. 2013, Espejo-Serna 2012, Espejo-Serna & López-Ferrari 2018, García-Mendoza et al. 2004, García-Mendoza & Meave 2011, González-Gallegos et al. 2014, Hernández-López 1995a, León-Velasco 2014, Martínez et al. 2017, Martínez-Gordillo et al. 2017, Sánchez-Ken 2019, Villaseñor 2016, 2018). Para la nomenclatura se usó, en el caso de helechos y plantas afines la propuesta de Christenhusz et al. (2011b), para gimnospermas a Christenhusz et al. (2011a) y para las angiospermas APG IV (2016). La estandarización de todos los nombres se realizó con el paquete Taxonstand v. 2.1 (Cayuela et al. 2012) para R (R Core Team 2018), la cual se basa en The Plant List (2013). Para el caso de taxones irresueltos, se consensó de acuerdo con Villaseñor (2016) y analizando la historia nomenclatural del taxón (Croft et al. 1999). Se consultó La Lista Roja de la UICN (IUCN 2017) y la NOM-059-SEMARNAT (SEMARNAT 2010) para verificar las categorías de riesgo en las que se encuentran los taxones.
La información taxonómica, geográfica, ecológica y curatorial para los taxones endémicos de la SMS fue capturada en la base de datos Vitex (Sahagún-Godínez 2003). La información fue proyectada y cartografiada con QGIS 2.16.3 (QGIS Development Team 2015). Se consideró como especie endémica a aquella con puntos de ocurrencia exclusivos dentro de la provincia biogeográfica SMS.
Se cuantificó el número de especies endémicas en la SMS por familia y géneros, así como por subprovincias, estados y tipos de vegetación. Para clasificar los tipos de vegetación presentes en la SMS, se utilizaron los polígonos de la vegetación potencial de México propuestos por el INEGI (2017), reclasificados de acuerdo a Rzedowski (1978) y proyectados en QGIS 2.16.3 (QGIS Development Team 2015). Se registraron ocho tipos de vegetación: el bosque de coníferas y encino (BCE), el bosque mesófilo de montaña (BMM), el bosque tropical caducifolio (BTC), el bosque tropical subcaducifolio (BTSC), el bosque tropical perennifolio (BTP), el matorral xerófilo (MX), el pastizal y palmar (PP) y otros tipos de vegetación (OTV). Para comparar la riqueza de especies en áreas con diferente tamaño (subprovincias biogeográficas), se utilizó el índice de biodiversidad taxonómica, que calcula el número de especies registradas divididas por el logaritmo natural del área en km2 (IB = E/lnA), donde E es el número de especies registradas y A el área (Squeo et al. 1998).
Resultados
Riqueza endémica. La flora vascular endémica de la SMS comprendió 6,020 registros correspondientes a 1,133 especies que se incluyen en 370 géneros y 91 familias de plantas vasculares endémicas (Material suplementario, Tabla S1) (Figuras 2 y 3). La flora mostró un claro predominio de Eudicotiledóneas, con la menor riqueza en helechos y licofitas (Tabla 1). La ubicación geográfica de 1,133 localidades tipo se muestra en la Figura 1.
Tabla 1 Riqueza de especies endémicas en la Sierra Madre del Sur, por grupo taxonómico.
| Familias | Géneros | Especies | |
|---|---|---|---|
| Licofitas | 1 | 1 | 3 |
| Helechos | 8 | 11 | 19 |
| Gimnospermas | 2 | 3 | 11 |
| Magnólidas | 4 | 9 | 29 |
| Monocotiledóneas | 13 | 77 | 272 |
| Eudicotiledóneas | 63 | 269 | 799 |
| TOTAL | 91 | 370 | 1,133 |

Figura 2 Especies de plantas vasculares endémicas de la Sierra Madre del Sur, México. A) Pinus georginae. B) P. jaliscana. C) P. luzmariae. D) Zephyranthes ita-andivi. E) Agave gypsicola. F) A. cremnophila. G) Bessera tuitensis. H) Tigridia hintonii. I) T. pugana. J) T. inusitata. K) Fosteria oaxacana. L) Laelia furfuracea Lindl. M) Prosthechea karwinskii. N) Rhynchostele madrensis. Ñ) Isochilus oaxacanus. O) Maianthemum comaltepecense. P) Matudanthus nanus. Q) Echeandia llanicola. Fotos. A. Rodríguez (H), L.M. Vázquez-García (H, J), J. Aragón-Parada (A-D, F, K-Q), J.P. Ortiz-Brunel (G) y P. Carrillo-Reyes (E).

Figura 3 Especies de plantas vasculares endémicas de la Sierra Madre del Sur, México. A) Dahlia hintonii. B) Hieracium macdonaldii. C) Mixtecalia teitaensis. D) Ruellia jaliscana. E) Sedum compactum. F) S. chloropetalum. G) S. commixtum. H) S. grandisepalum. I) Echeveria viridissima. J) E. globulosa. K) E. subcorymbosa. L) E. moranii. M) Prockia oaxacana. N) Salvia mexiae. Ñ) S. confertispicata. O) Galphimia langlassei. P) Mandevilla mexicana. Q) Euphorbia xylopoda. R) E. colligata. S) Lupinus jaimehintoniana. T) Cephalocereus parvispinus. U) Pinguicula laueana. V) Vallesia conzattii. W) Begonia lachaoensis. Fotos. A. Castro-Castro (A), P. Carrillo-Reyes (D-G, K, N, O, R, T) y J. Aragón-Parada (B, C, H-J, L-M, Ñ, P, Q, S, U-W).
Las familias con mayor número de géneros y de especies endémicas en la SMS fueron Asteraceae (75 y 250, respectivamente), Orchidaceae (40, 123), Rubiaceae (22, 74), Bromeliaceae (5, 64) y Lamiaceae (10, 63), Fabaceae (22, 52) y Crassulaceae (4, 49) (Tabla 2). Los géneros que registraron mayor número de especies endémicas fueron (Tabla 2): Salvia (46), Ageratina (39), Tillandsia (37), Echeveria (29), Lepanthes y Verbesina (22 cada uno). Se registraron 15 géneros endémicos de la SMS que en su conjunto agrupan 20 especies (Material suplementario, Tabla S1): Omiltemia (3), Amoana, Petronymphe y Stramentopappus (2 cada uno), así como Ainea, Artorima, Bellizinca, Dahliaphyllum, Fosteria, Matudanthus, Mixtecalia, Ortegocactus, Paneroa, Placocarpa y Wimmeranthus (una especie cada uno).
Tabla 2 Familias con mayor número de géneros y taxones endémicos de la Sierra Madre del Sur.
| Familias | Géneros | Especies | Género | Especies |
|---|---|---|---|---|
| Asteraceae | 75 | 250 | Salvia | 46 |
| Orchidaceae | 40 | 123 | Ageratina | 39 |
| Rubiacaea | 22 | 74 | Tillandsia | 37 |
| Bromeliaceae | 5 | 64 | Echeveria | 29 |
| Lamiaceae | 10 | 63 | Lepanthes | 22 |
| Fabaceae | 22 | 52 | Verbesina | 22 |
| Crassulaceae | 4 | 49 | Stevia | 19 |
| Asparagaceae | 9 | 34 | Bouvardia | 17 |
| Euphorbiaceae | 7 | 24 | Epidendrum | 16 |
| Melastomataceae | 2 | 20 | Sedum | 15 |
| Acanthaceae | 9 | 18 | Lobelia | 12 |
| Apocynaceae | 9 | 18 | Psacalium | 12 |
| Cactaceae | 5 | 17 | Arachnothryx | 11 |
| Campanulaceae | 4 | 15 | Castilleja | 10 |
| Poaceae | 9 | 14 | Mammillaria | 9 |
De acuerdo con el índice de diversidad taxonómica, la subprovincia SOR concentró la mayor riqueza florística por km2 (80.58), seguida de la SOC (18.96) y la SCE (5.33). En cuanto a especies registradas, la subprovincia SOR registró el mayor número de endemismos, con 940 especies, seguida de la SOC, con 183 y la SCE, con 49. Únicamente Ruellia jaliscana Standl., Gonolobus spiranthus Juárez-Jaimes, W.D. Stevens & Lozada-Pérez y Begonia lachaoensis Ziesenh. se distribuyen en las tres subprovincias de la SMS y 27 especies se comparten entre dos subprovincias (Tabla 3). El estado en el que se registró el mayor número de especies endémicas fue Oaxaca (731), seguido de Guerrero (310), Jalisco (189), Michoacán (36), Puebla (14), Veracruz (12) y Colima (ocho) (Tabla 4).
Tabla 3 Riqueza de especies endémicas por grupos taxonómicos presentes en las subprovincias de la Sierra Madre del Sur. SOC = Subprovincia Sierra Madre del Sur Occidental, SCE = Sierra Madre del Sur Central, SOR = Sierra Madre del Sur Oriental.
| Grupos taxonómicos | SOC | SCE | SOR | SOC/SCE | SOC/SOR | SCE/SOR | SOC/SCE/SOR |
|---|---|---|---|---|---|---|---|
| Total | 183 | 49 | 940 | 3 | 15 | 9 | 3 |
| Eudicotiledóneas | 118 | 31 | 638 | 3 | 13 | 8 | 3 |
| Monocotiledóneas | 36 | 5 | 230 | 0 | 1 | 1 | 0 |
| Magnólidas | 5 | 0 | 22 | 0 | 1 | 0 | 0 |
| Helechos y Licofítas | 3 | 0 | 19 | 0 | 0 | 0 | 0 |
| Gimnospermas | 3 | 1 | 7 | 0 | 0 | 0 | 0 |
Tabla 4 Riqueza de especies por estado que forman parte de la Sierra Madre del Sur. COL = Colima, GRO = Guerrero, JAL = Jalisco, MICH = Michoacán, OAX = Oaxaca, PUE = Puebla, VER = Veracruz.
| Grupos taxonómicos | COL | GRO | JAL | MICH | OAX | PUE | VER |
|---|---|---|---|---|---|---|---|
| Total | 8 | 310 | 188 | 37 | 731 | 14 | 12 |
| Eudicotiledóneas | 5 | 209 | 139 | 31 | 517 | 11 | 7 |
| Monocotiledóneas | 3 | 84 | 38 | 5 | 177 | 2 | 3 |
| Magnólidas | 0 | 13 | 5 | 0 | 15 | 0 | 1 |
| Helechos y Licofítas | 0 | 4 | 3 | 0 | 17 | 0 | 0 |
| Gimnospermas | 0 | 0 | 3 | 1 | 5 | 1 | 1 |
Especies bajo alguna categoría de riesgo. De las 1,133 especies de plantas vasculares endémicas en la SMS, 125 se identificaron bajo alguna categoría de riesgo (Material suplementario, Tabla S1). En la lista de la IUCN (2017), se incluyen una en la categoría Extinta, 15 especies en la categoría En Peligro Crítico, 30 En Peligro, cinco en Vulnerable, dos en Casi Amenazada y seis en Preocupación Menor. En la NOM-059 (SEMARNAT 2010), 15 especies se enlistan como En Peligro de Extinción, 24 Amenazadas y 27 Sujetas a Protección Especial. Un total de 11 taxones se reportaron en ambas listas. Las familias con el mayor número de especies dentro de alguna categoría de riesgo son Orchidaceae (24 especies), Cactaceae (15), Zamiaceae (ocho), Magnoliaceae (siete), Rubiaceae (seis) y Crassulaceae (cuatro) (Material suplementario, Tabla S1).
Tipos de vegetación. El bosque de coníferas y encino presentó el mayor número de especies endémicas (828), seguido del bosque mesófilo de montaña (235), el bosque tropical caducifolio (114), el bosque tropical subcaducifolio (94), el matorral xerófilo (32), el bosque tropical perennifolio (nueve), el palmar y pastizal (cuatro) y otros tipos de vegetación (dos) (Tabla 5, Figura 4).
Tabla 5 Riqueza de especies por grupo taxonómico y tipos de vegetación presentes en la Sierra Madre del Sur. BCE = Bosque de coníferas y encino, BTP = Bosque tropical perennifolio, BTSC = Bosque tropical subcaducifolio, BTC = Bosque tropical caducifolio, BMM = Bosque mesófilo de montaña, MX = Matorral xerófilo, PP = Pastizal y palmar, OTV = Otros tipos de vegetación.
| Grupos taxonómicos | BCE | BTP | BTSC | BTC | BMM | MX | PP | OTV |
|---|---|---|---|---|---|---|---|---|
| Total | 828 | 9 | 94 | 114 | 235 | 32 | 4 | 2 |
| Eudicotiledóneas | 621 | 6 | 73 | 83 | 149 | 21 | 1 | 1 |
| Monocotiledóneas | 189 | 1 | 19 | 28 | 58 | 11 | 3 | 0 |
| Magnólidas | 11 | 0 | 2 | 0 | 16 | 0 | 0 | 0 |
| Helechos y Licofítas | 12 | 0 | 0 | 0 | 10 | 0 | 0 | 0 |
| Gimnospermas | 7 | 2 | 0 | 3 | 2 | 0 | 0 | 1 |
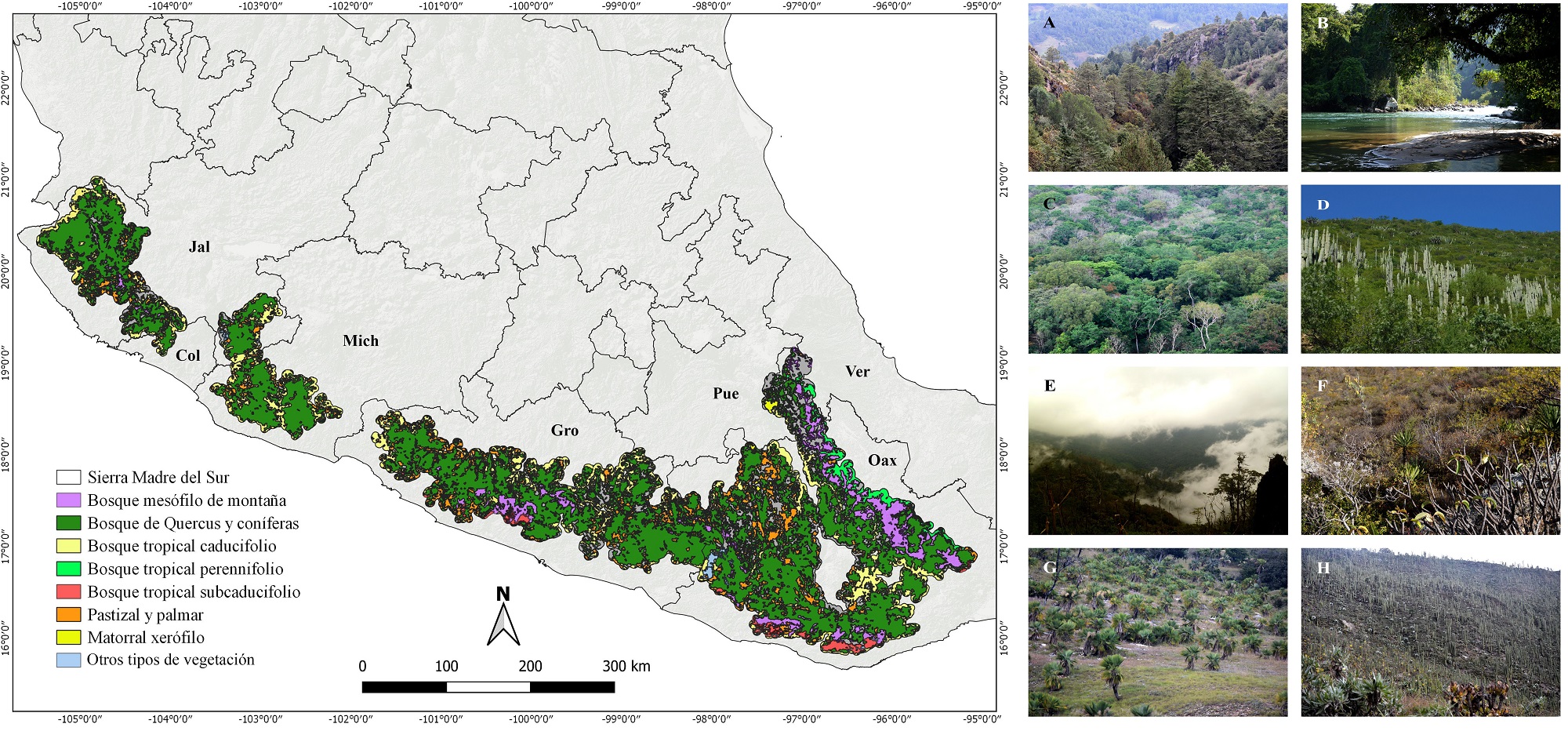
Figura 4 Vegetación potencial presentes en la SMS (basado en INEGI 2017). A) Bosque de coníferas y encino, B. Bosque tropical perennifolio. C. Bosque tropical subcaducifolio. D. Bosque tropical caducifolio. E. Bosque mesófilo de montaña. F. Matorral xerófilo. G. Palmar y pastizal. H. Vegetación gipsófila en el bosque tropical caducifolio.
Discusión
Con 1,133 especies endémicas, la SMS alberga ca. 10 % de la flora endémica mexicana, estimada en 11,600 especies (Villaseñor 2016). Esta cifra representa el 16.6 % de las 7,016 especies de plantas vasculares reportadas para la SMS (Espinosa et al. 2016). La riqueza del endemismo es similar al estimado para la SMOR por Salinas-Rodríguez et al. (2017) y mayor que para la FVT reportada por Rzedowski (2020) (Tabla 6). Previo al presente estudio, el endemismo en la SMS se había identificado de manera puntual para determinados grupos taxonómicos o funcionales, por ejemplo, helechos (Tejero-Díez et al. 2016), gimnospermas (Contreras-Medina 2016), En el caso de las angiospermas, diversos trabajos han reportado el endemismo por familias, grupos y géneros en específico (Castro-Castro et al. 2012, Vargas-Amado et al. 2013, Munguía-Lino et al. 2015, Aragón-Parada et al. 2019). Por otra parte, aunque no usaron un enfoque de regiones biogeográficas, Ibarra-Manríquez et al. (2015b) identificaron que el endemismo en las lianas de México se concentra en los estados de Oaxaca, Jalisco, Guerrero y Michoacán, en tanto que Tellez et al. (2020) identificaron seis principales áreas de concentración de especies endémicas de árboles, de las cuales, tres, se encuentran en los estados de Jalisco, Guerrero y Oaxaca.
Tabla 6 Comparación de grupos con alto número de especies endémicas entre diferentes provincias biogeográficas de la Zona de Transición Mexicana. SMOR = Sierra Madre Oriental, FVT = Faja Volcánica Transmexicana, SMS = Sierra Madre del Sur.
| Estudios | Familias | Géneros | Especies | Especies por familia | ||
|---|---|---|---|---|---|---|
| Asteraceae | Lamiaceae | Fabaceae | ||||
| SMOR (Salinas-Rodríguez et al. 2017) | 88 | 369 | 1,135 | 232 | 66 | 66 |
| FVT (Rzedowski 2020). | 71 | 213 | 544 | 118 | 18 | 32 |
| SMS (este trabajo). | 91 | 370 | 1,133 | 250 | 63 | 52 |
Familias y géneros. En México, 25 familias de plantas vasculares concentran el 65 % de la riqueza de la flora nacional (Villaseñor 2016). Dentro de éstas, las familias con mayor endemismo son Asteraceae, Fabaceae, Poaceae, Orchidaceae, Cactaceae y Rubiaceae (Rzedowski 1991b). Nuestros resultados concuerdan con la relevancia de estas familias (Tabla 2). Este patrón es similar al reportado por Salinas-Rodríguez et al. (2017) para la SMOR y Rzedowski (2020) para el EVT (Tabla 6).
Según Rzedowski (1991a), la representatividad de Asteraceae es marcada en el norte y centro de México. En cuanto a endemismo, Villaseñor (2018) registró 1,988 especies endémicas en el centro y sur del país en los bosques templados. Este último autor reporta que la mayor riqueza y endemismo para esta familia se registra en los estados del occidente y SE de México. Nuestros datos apoyan este patrón, pues se registraron 250 especies endémicas en la SMS (Tabla 6). Los estados que integran la SMS figuran entre los más diversos en endemismo de Asteraceae (Suárez-Mota & Villaseñor 2011, Villaseñor 2016, 2018). Los géneros más diversos en endemismo de esta familia en la SMS también concuerdan con lo reportado por Villaseñor (2018) (Tabla 2). La SMS, alberga el 28.8 % del endemismo de Ageratina, 20 % de Stevia y el 15.9 % de Verbesina. Se registraron tres géneros endémicos de Asteraceae para la SMS: i) Stramentopappus, que se distribuye en el bosque mesófilo de montaña de Oaxaca, en la vertiente del Golfo de México (Redonda-Martínez & Villaseñor 2014) y ii) Mixtecalia y Paneroa, que se encuentran en la Mixteca oaxaqueña (Schilling 2008, García-Mendoza et al. 2020).
Para la familia Orchidaceae se estima que al menos el 35 % de sus especies son endémicas del país (Rzedowski 1991b). Espejo-Serna (2012) reportó 585 especies endémicas de México, la mayoría presentes en el bosque mesófilo de montaña y los bosques tropicales. Las 125 especies endémicas de Orchidaceae registradas para la SMS en el presente estudio, principalmente en estos tipos de vegetación, reafirma lo señalado por los autores antes mencionados. Así mismo, dos de sus géneros concuerdan con los de mayor endemismo en monocotiledóneas para México (Tabla 2). Si se comparan las especies endémicas aquí registradas, la SMS resguarda el 46.6 % de Lephanthes y 33.3 % de Epidendrum (Espejo-Serna 2012). Se registraron dos géneros de Orchidaceae endémicos de la SMS. Amoana y Artorima, de hábito epífito y rupícola, respectivamente; el primero de éstos tiene dos especies restringidas al bosque de coníferas y encino, en Oaxaca, mientras que el segundo se encuentra en el bosque de coníferas y encino y en el bosque mesófilo de montaña, en Guerrero y Oaxaca (Leopardi et al. 2012).
Se estima que México resguarda alrededor de 677 especies de Rubiaceae y de éstas el 69 % son endémicas (Villaseñor 2016, Rzedowski 1991b). Las 74 especies aquí reportadas se encuentran principalmente en el bosque tropical subcaducifolio y bosque tropical caducifolio, en los cuales es sumamente diversa (Borhidi & Diego-Pérez 2002, Rzedowski & Calderón de Rzedowski 2013). Se registraron tres géneros endémicos de Rubiaceae de la SMS, Bellizinca, Omiltemia y Placocarpa. El primero es monotípico y se registra sólo en Oaxaca, en el bosque mesófilo de montaña (Borhidi et al. 2004). Omiltemia se restringe a Guerrero, en el bosque mesófilo de montaña y en el ecotono de éste con el bosque de coníferas y encino (Lozada-Pérez & RojasGutiérrez 2016). Placocarpa es también monotípico y se encuentra en Oaxaca y Veracruz, en el bosque de coníferas y encino.
Espejo-Serna & López-Ferrari (2018) registraron 422 especies de Bromeliaceae para México y de estas 318 son endémicas. Las 64 especies de la SMS concuerdan con las áreas de mayor riqueza y endemismo reportado por estos autores, las cuales se encuentran en los bosques templados y en el bosque mesófilo de montaña. En México, Tillandsia es el tercer género másdiverso con 230 especies (Villaseñor 2016, Espejo-Serna & López-Ferrari 2018). En la SMS se registraron 37 de las 177 especies endémicas del género, lo que representa el 19.7 %. Los otros dos géneros con elevado endemismo, tanto en México como en la SMS, son Hechtia (70 México/11 SMS) y Pitcairnia (41/8) (Espejo-Serna 2012, Espejo-Serna & López-Ferrari 2018).
Martínez-Gordillo et al. (2017) mencionan que Lamiaceae es más abundante en los bosques templados de las zonas montañosas. Los mismos autores reportan que en México se distribuyen 598 especies, de las cuales 396 son endémicas. Al ser el bosque de coníferas y encino dominante en la SMS, se esperaba encontrar un elevado número de especies endémicas, lo cual fue ratificado con 63 especies registradas (Tabla 6). Villaseñor (2016) reportó al género Salvia como el más diverso de México, con 328 especies. De este género, Martínez-Gordillo et al. (2017) registraron 236 endémicas y Martínez-Gordillo et al. (2017) mencionan que la diversidad en endemismo se concentra en los estados de Jalisco, Oaxaca, Guerrero, Puebla y Michoacán, mismos que forman parte de la SMS. Las 46 especies aquí enumeradas lo posicionan como el género más diverso en la SMS.
Fabaceae, con 1,903 especies, es la segunda familia más diversa en México (Villaseñor 2016). Su riqueza aumenta en general con el ascenso de la temperatura (Rzedowski 1991a) y al menos el 52 % de sus especies son endémicas de México (Rzedowski 1991b, Rzedowski & Calderón de Rzedowski 2013). Las 52 especies aquí enumeradas crecen principalmente en el bosque templado y el bosque tropical caducifolio; este número es similar al reportado por Salinas-Rodríguez et al. (2017) para la SMOR y Rzedowski (2020), para la FVT (Tabla 6). Los géneros, Dalea, Desmodium, Lupinus y Phaseolus son los mejor representados. Sousa & Delgado (1998) mencionan que Calliandra, Dalea y Phaseolus son géneros americanos, mientras que Desmodium y Lupinus son de amplia distribución.
Villaseñor (2016) reporta 372 especies de Crassulaceae para México; Pérez-Calix &Franco-Martínez (2004) mencionan que alrededor de 294 son endémicas. Las 54 especies endémicas para la SMS son comparables al número reportado por Salinas-Rodríguez et al. (2017) para la SMOR y por Rzedowski (2020), para el EVT (Tabla 6). En su mayoría, en el área de estudio se encuentran presentes en el bosque de coníferas y encino. Los géneros Echeveria (29) y Sedum (15) son los más diversos. En México están documentadas 132 especies de Echeveria y 133 de Sedum (Pérez-Calix 2008, Villaseñor 2016). Las especies de Echeveria y Sedum concuerdan con Pérez-Calix &Franco-Martínez (2004) y Aragón-Parada et al. (2019), quienes mencionan su preferencia hacia los bosques templados.
Se registraron también, como géneros endémicos de la SMS a Dahliaphyllum (Apiaceae), Ortegocactus (Cactaceae) y Wimmeranthus (Campanulaceae). El primero se reporta del bosque mesófilo de montaña, en el municipio de Atoyac de Álvarez, Guerrero (Constance & Breedolve 1994), Ortegocactus se conoce de pocas poblaciones en el bosque tropical caducifolio de Oaxaca y Wimmeranthus, se encuentra en el bosque de coníferas y encino de San Juan Mixtepec en la Mixteca Oaxaqueña (Rzedowski 2018). Se reportan también cuatro géneros endémicos de monocotiledóneas: Petronymphe (Asparagaceae), Ainea, Fosteria (Iridaceae) y Matudanthus (Commelinaceae). El primero incluye dos especies gipsófilas de áreas restringidas en Guerrero y Oaxaca (Gutiérrez et al. 2016). Los géneros de Iridiaceae son monotípicos y sus especies son geófitas del bosque de coníferas y encino de Oaxaca (Munguía-Lino et al. 2015). Por último, Matudanthus se distribuye en el bosque de coníferas y encino, en Guerrero y Oaxaca (Hunt 1978). Los 15 géneros endémicos reportados para la SMS representan un pequeño porcentaje (6.8 %) de los 219 contabilizados para México por Villaseñor (2004) y son una cifra equiparable a los 13 señalados para la SMOR (Salinas-Rodríguez et al. 2017), pero contrastante con el reporte de un único género endémico al EVT (Rzedowski 2020).
Subprovincias y estados. El índice de diversidad taxonómica y las especies que se registraron mostraron que la SOR fue la que registró la mayor riqueza comparada con la SCE y la SOC (Tabla 3). Estos valores pueden deberse a diferentes aspectos que se describen a continuación.
Oaxaca abarca la mayor extensión de la SOR y en él se han recolectado ejemplares botánicos desde el siglo XVIII y probablemente es el estado mejor conocido florísticamente de los que conforman la SMS. García-Mendoza & Meave (2011) mencionan 728 especies endémicas que, en conjunto, se distribuyen en todo el territorio oaxaqueño. En el presente estudio se registran 731 especies, y 611 son endémicas de Oaxaca (83.9 %).
Para el estado de Guerrero, Villaseñor & Ortiz (2014) reportaron 262 especies endémicas, un número menor al que se reporta en el presente trabajo (310 especies); de éstas, 176 son endémicas del estado y representan el 67.1 % del endemismo estatal y 134 se comparten principalmente con el estado de Oaxaca. Esta relación es consistente con lo encontrado por Ibarra-Manríquez et al. (2015a), quienes, basándose en la composición de lianas, identificaron estrechas afinidades florísticas entre ambos estados. Sin duda el número de endemismo en Guerrero se elevará cuando se integre un mayor conocimiento florístico del estado.
Hernández-López (1995a) registró para Jalisco y áreas adyacentes 501 especies endémicas. Si se considera que las montañas del SO del estado forman parte de la SCE y SOC de la SMS, con 188 especies aquí reportadas, éstas representan el 32.9 % de la flora endémica estatal. Hernández-López (1995b) cita para la flora de la ANP Sierra de Manantlán, dentro de la SOC, 31 especies endémicas, mientras que Morales-Arias et al. (2016) enumeran 32 especies endémicas en la flora vascular de Villas de Cacoma y Harker et al. (2015) y Hernández-López et al. (2021) mencionan que el municipio de San Sebastián del Oeste es un área importante en endemismos, ambas áreas dentro de la subprovincia SOC, de la SMS.
La porción geográfica de la provincia SOC en Colima es parte de la Reserva de la Biósfera Sierra de Manantlán y para esta área se encontraron cuatro endemismos estatales, dos de los cuales se incluyen en el trabajo de Hernández-López (1995a) y cuatro corresponden a especies que se comparten con Jalisco y Michoacán. Para el caso de Puebla y Veracruz, con 14 y 12 endemismos cada uno, no se encontraron trabajos que reporten el endemismo para las porciones de los estados que forman parte de la SMS. Además de esto, una de las áreas menos estudiadas, pero con importante número de especies endémicas, es la SCE en Michoacán y Jalisco. Los endemismos aquí reportados son 37, de éstos 17 son únicos y 20 se comparten con la SOC y SOR. Estos son los primeros datos consensados para esta área geográfica que forma parte de la SMS.
Especies en riesgo. Las orquídeas, cícadas y las plantas suculentas incluidos agaves, crasuláceas y cactáceas se encuentran entre los grupos más carismáticos y atractivos para los coleccionistas, lo que comúnmente las coloca dentro de alguna categoría de riesgo (Grace 2019, IUCN 2017). De las 124 especies aquí reportadas en alguna categoría de riesgo, 31.1 % pertenecen a estos grupos. Por otro lado, las 40 especies aquí enumeradas que se encuentran en el bosque mesófilo de montaña sufren presiones por la pérdida y fragmentación de su hábitat (González-Espinosa et al. 2011), lo que las incluye bajo alguna categoría de riesgo. Los géneros Dioon y Ceratozamia (Zamiaceae) en la SMS, con siete especies, se encuentran bajo alguna categoría de riesgo, ya sea Vulnerable, En Peligro Crítico o en Peligro, lo que puede deberse a la fragmentación de su hábitat por agricultura o ganadería extensiva, así como al reducido número de individuos en sus poblaciones y las pequeñas áreas geográficas en las que se encuentran. La Norma Oficial Mexicana NOM-059 (SEMARNAT-2010) y la IUCN Red List se encargan de registrar estas especies en el nivel nacional y mundial; sin embargo, aunque consideran a las especies endémicas como organismos susceptibles de desaparecer por su reducida área de distribución y densidad poblacional, en la mayoría de los casos no se cuenta con información suficiente sobre su distribución geográfica ni con estudios demográficos. Es imperativo reconsiderar las estrategias de conservación, no sólo de especies carismáticas, sino también de todas las especies con distribución geográfica restringida.
Vegetación. En cuanto al número de especies, los datos presentados aquí son similares con los publicados por Villaseñor (2016), quien menciona que los bosques templados y el bosque mesófilo de montaña albergan una alta riqueza de especies y son después de los matorrales xerófilos los que concentran un importante número de endemismos. Rzedowski (2019) indica que en afinidad ecológica destacan en cantidad de endemismos los componentes heliófilos de los matorrales, pero que también son frecuentes los encinares y pinares, así como los elementos endémicos adaptados a mayor humedad de los bosques mesófilos de montaña. Cuando comparamos los resultados con Salinas-Rodríguez (2017) para la SMOR, éstos varían, pues los autores señalan que los matorrales y sus variantes son un componente característico y dominante, el cual se influye por las condiciones de aridez que se crean en la ladera de sotavento, debido al efecto de sombra orográfica, en contraste con la SMS, donde el bosque de coníferas y encino cubre el 47 % de la superficie, promovido por mayores concentraciones de humedad en estas latitudes.
Para el BCE se registraron 814 especies exclusivas distribuidas en sus diferentes comunidades vegetales, ya sea bosque de pino-encino, bosque de encino-pino o bosque de pino-oyamel (Tabla 5). Trabajos en los que evalúan la diversidad y endemismo de diferentes grupos vegetales en México resaltan a este tipo de vegetación y la SMS como un área importante de endemismo (Castro-Castro et al. 2012, Vargas-Amado et al. 2013, Munguía-Lino et al. 2015, Anguiano-Constante et al. 2018, Aragón-Parada et al. 2019). García-Mendoza et al. (2004) y García-Mendoza & Meave (2011) mencionan que el BCE concentra el mayor número de endemismo y riqueza en Oaxaca. McDonald (2013) reconoce dentro de este tipo de vegetación una comunidad que denomina vegetación subalpina, con 19 especies endémicas. Nuestros datos apoyan este supuesto, pues se registraron más de 54 especies que se concentran en las altas montañas de la SMS. Ejemplo de estas áreas son el Cerro Quiexobra, Cerro Pelón-Humo Chico, Cerro Zempoaltépetl y Cerro Teotepec.
Según la Comisión Nacional para el Conocimiento y Uso de la Biodiversidad (CONABIO 2010), el BMM es el ecosistema más amenazado a nivel nacional y lo clasifica como la comunidad vegetal más compleja, que alberga una gran riqueza de especies, tanto endémicas como amenazadas. En la Tabla 5 se señala que 229 especies son endémicas del BMM y 6 se comparten principalmente con el BCE. Dentro de las 235 especies endémicas del BMM destacan las especies epífitas y arbustivas de Asteraceae, Bromeliaceae, Orchidaceae y Rubiaceae. Esto concuerda con Rzedowski (1996), quien refiere al BMM como una comunidad en la que los elementos boreal, tropical y nativo dan forma a una compleja interacción biótica en la que las familias mencionadas (excepto Bromeliaceae) tienen un importante número de especies endémicas. Estos datos fueron corroborados por Villaseñor (2010), quien reportó para el BMM de México 2,822 especies endémicas.
En el BTC se registraron 64 especies exclusivas, 50 se comparten con el MX o bien con el BCE; el 24 % del área geográfica de la SMS es BTC y su endemismo es menor que el BMM, pero mayor al del BTSC. Rzedowski (1991a) menciona que aproximadamente el 40 % de las especies del BTC son endémicas. Acosta et al. (2003) registraron 11 especies endémicas en la cuenca alta del Río Tehuantepec, en el estado de Oaxaca.
El BTSC que se distribuye en la vertiente del Pacífico de la SMS, alberga 57 especies endémicas exclusivas y 37 compartidas con el BMM y BCE. Rzedowski (1978) mencionó al BTSC como un conjunto de comunidades vegetales con características intermedias en su fisionomía y en sus requerimientos climáticos entre el BTP y BTC. En la SMS se han realizado colectas en comunidades de BTSC, en Jalisco, Guerrero Michoacán y Oaxaca, pero se desconocen las especies endémicas. Un área importante de la cual se han descrito al menos 16 especies nuevas para la ciencia es el BTSC de San Miguel del Puerto, en Oaxaca (Borhidi 2012, Leopardi et al. 2012, Lott & Martínez-Gordillo 2012).
El BTP en la SMS se encuentra en la vertiente del Golfo y se registraron nueve especies endémicas. Esto concuerda con Rzedowski (1991a), quien aseveró que en esta comunidad vegetal se presenta el menor endemismo en México, pues sus componentes tienen una distribución amplia, de afinidad meridional.
El MX ocupa una pequeña porción en el extremo N de la SMS, con límites en el Valle de Tehuacán-Cuicatlán y en la Mixteca oaxaqueña. Valiente-Banuet et al. (2000) mencionaron que el Valle de Tehuacán-Cuicatlán y áreas adyacentes es una zona con extraordinaria heterogeneidad ambiental y ésta determina la diversidad y distribución de las comunidades vegetales. Rzedowski (1991a) reconoció que, en el MX, alrededor del 60 % de sus componentes son endémicos. En esta porción del MX se registraron 16 especies exclusivas y 16 especies que se comparten con el BTC y BCE. Rzedowski (1978) reportó al Valle de Tehuacán-Cuicatlán como una de las áreas con mayor endemismo en México. Sosa & De-Nova (2012) mencionan que esta área registra los valores más altos de endemismo. García-Mendoza et al. (1994) registraron 163 taxones endémicos en la Mixteca Alta de Oaxaca y Puebla. En el MX se encuentran enclaves de afloramientos yesosos, de los que García-Mendoza et al. (2020) recientemente describieron a Mixtecalia. La vegetación gipsófila en proporción de extensión no se categorizó, pero registramos 21 especies endémicas a la SMS, de las cuales y al menos 17 se restringen a este tipo de vegetación. Las áreas de pastizales, palmares y otros tipos de vegetación en la SMS también son pequeñas, en muchos casos inducidas y presentes a orillas o dentro del BCE, BTSC o BTC. Los palmares y pastizales presentaron cuatro especies endémicas que se comparten con el BCE y dos especies se registraron además en otros tipos de vegetación. Según la SEMARNAT (2006), las actividades agropecuarias causan cambio de uso de suelo y modifican sustancialmente la vegetación del país, sin embargo, las áreas de vegetación secundaria juegan también un papel importante en la conservación de especies (e.g., Suárez-Mota et al. 2018)
Como se ha mencionado, la SMS es la provincia biogeográfica más compleja de México, tanto geológica, fisiográfica y florísticamente. La heterogeneidad de comunidades vegetales y climas hacen que en ella se distribuya el 10 % del endemismo nacional. Este se encuentra disperso en áreas dentro de las tres subprovincias que la conforma. Aún es necesaria la evaluación de estas áreas mediante métodos cuantitativos en los que se integren la historia evolutiva de los taxones, los atributos de rareza de las especies y el patrón espacial de diversidad filogenética en las áreas con mayor endemismo. Esto ayudará a delimitarlas y valorarlas para su conservación, manejo y posible uso y permitirá evaluar las áreas naturales protegidas ya establecidas dentro de la SMS con respecto al endemismo.
Material suplementario
El material suplementario de este manuscrito puede ser descargado de la siguiente liga: https://doi.org/10.17129/botsci.2682

