











 nova página do texto(beta)
nova página do texto(beta) Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO 
 Permalink
PermalinkLacandonia schismatica E. Martínez & C. H. Ramos (Triuridaceae) is a plant from the Lacandona forest in Chiapas, Mexico. It is a plant displaying distribution of gynoecium surrounding the androecium (Márquez-Guzmán et al. 1989, Martínez & Ramos 1989). The cell nucleus of this species is characterized by the presence of granules of 32 nm in diameter called Lacandonia granules (Jiménez-García et al. 1992). These particles are ribonucleoproteins (RNPs), intermediate in size and distribution between interchromatin and perichromatin granules present in mammalian cell nucleus. Lacandonia granules contain SR proteins and poly-A RNA tail as perichromatin granules (Agredano-Moreno & Jiménez-García 2000). In addition, the morphology of these particles was previously analyzed using atomic force microscopy (Fragoso-Soriano et al. 2009).
Lacandonia granules are also present in the related Triuris brevistylis Donn.Sm., also a member of the order Pandanales as L. schismatica (Stevens 2017). The observation of Lacandonia granules in the nuclei of the gymnosperm tree Ginkgo biloba L., (Jiménez-Ramírez et al. 2002), suggests that granules are present in other groups and may display a general function. In the present work we report the presence of Lacandonia granules in the nuclei of Welwitschia mirabilis Hook.f., a member of Welwitschiaceae (order Gnetales; Stevens 2017).
Materials and methods
Plants. W. mirabilis seeds were disinfected with 1 % captain for 1 hour and washed with deionized water. Seeds were cultivated in tepojal (small volcanic grain covered with clay), substrate in a germination chamber Lab-Line at 26 °C, 60 % relative humidity and photoperiod (light/dark): 16/8.
Transmission Electron Microscopy. 1 mm3 fragments of young leaves were processed following the standard protocol for electron microscopy (Jiménez-García & Segura-Valdez 2004). Briefly, fragments were fixed overnight at room temperature in a mixture of 6 % glutaraldehyde and 4 % paraformaldehyde, in PBS buffer (pH 7.2). Post-fixation was performed with 2 % osmic acid overnight. Samples were subsequently dehydrated in a graded series of ethanol and embedded in an epoxy resin at 60 °C for 48 h following the standard protocol for electron microscopy. Thin sections were placed on copper grids covered with formvar. Contrast was conducted with 5 % uranyl acetate and 0.5 % lead citrate. Grids were observed with a transmission electron microscope (JEOL, JEM 1010, Peabody, MA) working at 80 kV. Images were obtained with a charge-coupled device camera coupled to the microscope. The diameter of the nuclear granules was determined on 100,000x electron micrographs of thin sections stained with Bernhard´s EDTA technique (Bernhard 1969).
Light microscopy. Thin sections were stained with toluidine blue and 100x pictures were taken in brightfield illumination with an optical microscope (Nikon, Eclipse E800).
Atomic force microscopy. Atomic force microscopy was conducted as previously described (Jiménez-García & Segura-Valdez 2004, Segura-Valdez et al. 2010). Briefly, semithin sections (about 250 nm thickness) were mounted on glass slides and observed with an atomic force microscope (model BioScope, Digital Instruments, Santa Barbara CA, USA) working in contact mode. The scan size was from 30 to 100 µm at a scan rate of 2.1 Hz. Images were produced with the NanoScope IIIa control system. The AFM tips were silicon nitride tips with a curvature radius of 20-60 nm (model NP).
EDTA staining for RNPs. Thin sections of samples fixed with 6 % glutaraldehyde and 4 % paraformaldehyde without osmium tetroxide were used for Bernhard's EDTA technique, for preferential staining of ribonucleoproteins (RNPs) (Bernhard 1969). Basically, 5 % uranyl acetate was used for 3 min, followed by treatment with EDTA for 13 min and 0.5 % lead citrate for 3 min.
Osmium amine. Specific staining of DNA in the cell nucleus of W. mirabilis was performed according to Vázquez-Nin et al. (1995) with modifications. Briefly, 60-90 nm sections mounted in gold grids without formvar were floated on a drop of 5N HCl for 1 hour at room temperature (acid hydrolysis). Grids were rinsed with deionized water and incubated in a wet chamber containing a drop of osmium amine solution. Finally, grids were washed and observed with an electron microscope without additional staining.
Results
The cell nuclei of Welwitschia mirabilis were observed with light, atomic force, and transmission electron microscopy.
Compact chromatin. In the cell nuclei of Welwitschia mirabilis dense strands are observed between areas of low density (Figure 1A, B and C). The three dimensional arrangement of the strands is evident when they are observed with light (Figure 1A) and atomic force microscopy (Figure 1B). Conventional staining for transmission electron microscopy shows the strands heavily stained with uranyl acetate and lead citrate (Figure 1C). To determine whether the dense strands observed in the nuclei of W. mirabilis correspond to DNA, we used the osmium ammine technique specific for DNA. The strands were densely stained indicating that they correspond to DNA. Little or no DNA is present in the nucleoplasm and nucleolus and no staining is observed in the cytoplasm (Figure 2).
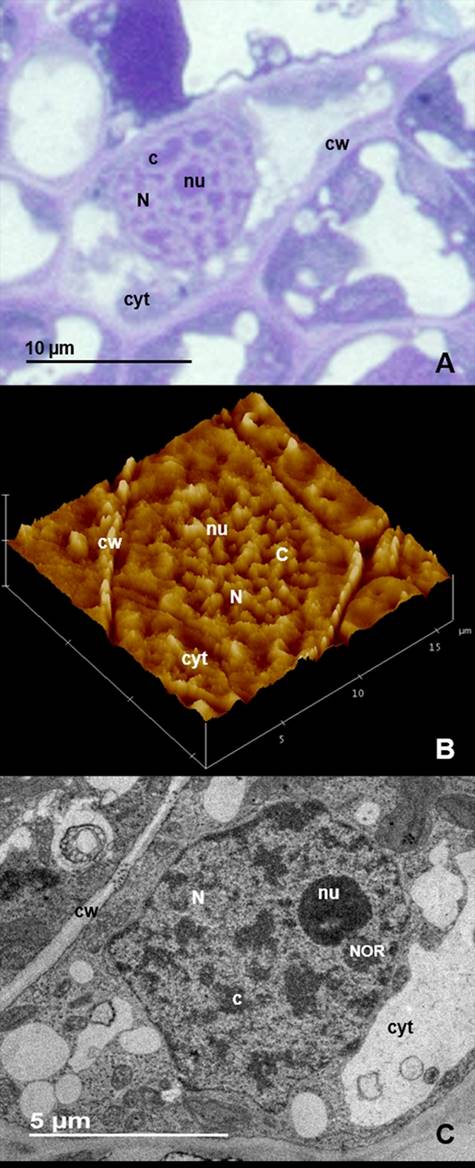
Figure 1 Welwitschia mirabilis cell nuclei show a reticulated pattern of compact chromatin. (A) Bright field of a cell nucleus stained with toluidine blue. (B) Atomic force microscopy of this organelle showing three dimensional arrangement of intranuclear strands. (C) Conventional staining for transmission electron microscopy showing strands of compact chromatin (c). Nucleus (N), cytoplasm (cyt), cell wall (cw), Nucleolar Organizer Region (NOR), nucleolus (nu).

Figure 2 Nucleus of W. mirabilis stained with osmium amine specific for DNA. Dense strands (c), are stained with this technique. Nucleolus (nu) and cytoplasm (cyt) are not stained.
Nuclear particles. The nucleoplasm of W. mirabilis is composed of a fibrogranular environment among the strands of chromatin, with electrodense particles about 32 ± 1.7 nm in diameter. Granules are associated or interconnected to electron dense fibers (Figures 3-4), both, in interchromatin and perichromatin areas (Figure 4). The EDTA regressive technique for ribonucleoproteins bleached the compact chromatin strands and granules and fibers are heavily stained (Figure 5). The association of W. mirabilis extranucleolar granules to reticulated chromatin, their location, abundance and ribonucleoproteic nature indicate that these particles correspond to Lacandonia granules reported in the species L. schismatica, T. brevystilis (Jiménez-García et al. 1992) and Ginkgo biloba (Jiménez-Ramírez et al. 2002).
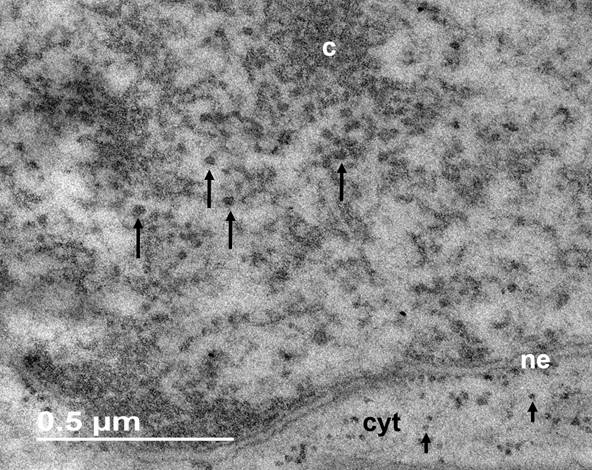
Figure 3 Nucleus of W. mirabilis stained with uranyl acetate and lead citrate. Abundant granules (large arrows), bigger than ribosomes (small arrows), are observed in the nucleoplasm. Cytoplasm (cyt). Chromatin (c), nuclear envelope (ne).
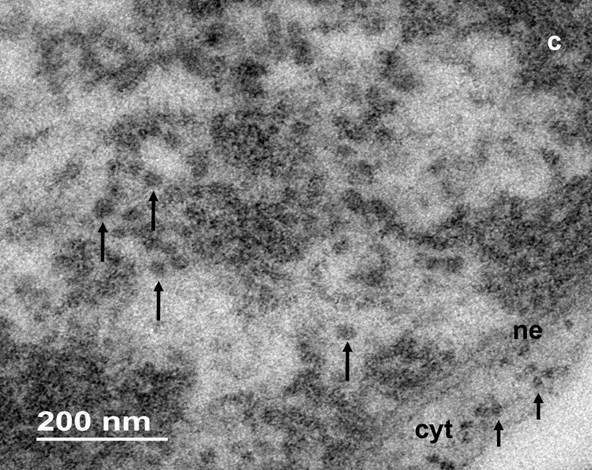
Figure 4 Intranuclear granules in a cell of W. mirabilis (large arrows). Ribosomes (small arrows) are indicated in the cytoplasm (cyt), compact chromatin (c), nuclear envelope (ne).

Discussion
Compact chromatin. Optical microscopy showed strands that stain with toluidine blue distributed in the nuclei of cells of Welwitschia mirabilis (Figure 1A). Thin sections of samples prepared for transmission electron microscopy and observed with the electron microscope corroborated the reticulated pattern of the strands (Figure 1B) and atomic force microscopy showed the three dimensional arrangement of the strands in the nucleoplasm. The strands are also positive for the osmium amine technique specific for DNA. Therefore, these strands correspond to the reticulated pattern reported previously using also results from atomic force microscopy (Jiménez-Ramírez et al. 2002).
Nuclear particles. In previous studies, the presence of ribonucleoprotein granules has been described in nuclei that have a reticulated pattern such as Lacandonia schismatica and G. biloba (Agredano-Moreno et al. 1994, Agredano-Moreno et al. 2000, Fragoso-Soriano et al. 2009, Jiménez-García et al. 1992, Jiménez-García & Fragoso-Soriano 2000, Jiménez-Ramírez et al. 2002). Therefore, we searched for Lacandonia granules in cell nuclei of W. mirabilis. Using standard transmission electron microscopy, we observed granules in the nucleoplasm, about 32 nm in diameter, associated to fibers in the periphery of chromatin. These granules were stained after the EDTA regressive staining preferential for ribonucleoproteins, while compact chromatin is observed with low contrast. These particles also may correspond to similar although scarce, particles observed in some members of bryophytes (Alonso-Murillo & Jiménez-García 2015).
Therefore, the size, distribution, shape, positivity to the EDTA technique, and fibrogranular arrangement of the nuclear particles in W. mirabilis reported here, correspond to Lacandonia granules described previously in the plant Lacandonia schismatica (Jiménez-García et al. 1992, Agredano-Moreno & Jiménez-García 2000, Jiménez-García & Fragoso-Soriano 2000, Jiménez-Ramírez et al. 2002, Fragoso-Soriano & Jiménez-García 2009).

